3个杨树品种移栽后生长特性比较
2021-03-09刘春鹏姚昱浓徐振华李向军
刘春鹏,姚昱浓,滑 磊,徐振华,李向军,王 法
(1河北省林业和草原科学研究院,河北 石家庄 050061;2河北省林木良种技术创新中心,河北 石家庄 050061;3华中农业大学 园艺林学学院,湖北 武汉 430070;4河北省英烈纪念园管理处,河北 石家庄 050000;5晋冀鲁豫烈士陵园管理处,河北 邯郸 056001)
近年来,我国北方主要造林树种杨树因“飞絮”污染问题而备受质疑[1-3],一些地区甚至明令禁止栽植杨树。然而,我国是世界第一木材资源消费大国,每年消耗木材量超过6亿m3,国内木材产量严重不足[4],尚有超过50%的木材需求来源于国外。而杨树具有生长迅速、适应性强等优点,被广泛用于营建人工用材林,其木材产量占国内木材总量的18.14%,是我国国产木材的主要来源之一[5-6]。在环境保护与木材需求的双重要求下,发展短周期的杨树人工林,并在开花前将其采伐,既能避免产生“飞絮”污染,又能满足我国大量的木材需求,是杨树产业发展的必然趋势[7]。
对于营建短周期杨树人工林而言,促进杨树前期快速生长尤为重要。然而,植物移栽后普遍存在缓苗过程[8-10],杨树也不例外,且以毛白杨的缓苗现象最为突出[11]。毛白杨是我国特有的乡土树种,与其他杨树品种相比,具有材质优良的巨大优势[12-13];20世纪90年代前一直占据着杨树生产的主导地位,之后逐渐被国外引进的欧美杨系列等所取代[14]。究其原因,主要是由于毛白杨造林后前期生长较慢,且缓苗期较长。因而解决毛白杨等杨树品种移栽后生长缓慢的问题,不仅能够增加短周期杨树人工林的木材产量,而且有助于杨树人工林材质优良品种的选择。
关于杨树品种移栽后早期缓苗的研究,目前主要集中在栽植后大水大肥管理、打头控侧等促进杨树移栽后生长的技术措施上[11,15],而有关其移栽后生长缓慢的机制研究等尚未见报道。本研究以欧美107杨、小美旱杨和毛白杨1316为试验对象,调查特定生长期间内各器官生物量及叶面积,比较3个杨树品种的同化产物生产、生物量分配、同化速度、净光合速率等,从生理生态学方面解释3个杨树品种移栽后的生长差异,以期为揭示一些杨树品种移栽后生长缓慢的成因提供理论支撑,并为促进移栽后快速生长的相关研究提供思路。
1 试验区概况
试验在河北省林业和草原科学研究院(114°28′ E,38°08′ N)进行。试验地位于河北省石家庄市新华区,在石家庄市区西北部,西接太行山,北临滹沱河。气候为暖温带大陆性季风气候,四季分明,降水分布不均,多集中于夏季,年均降水量552.8 mm。年均气温12.9 ℃,7月份最高,月均26.5 ℃左右;1月份最低,月均-2.9 ℃左右;极端最高和最低气温分别为41.5和-19.3 ℃,年无霜期200 d左右。试验地地势平坦,土壤肥力均匀,土质为沙壤土,pH值为7.5,富含有机质[16-17]。
2 材料与方法
2.1 试验材料
毛白杨1316(Populustomentosa‘1316’)、欧美107杨(Populus×euramericana‘74/76’)、小美旱杨(PopuluspopularisHsü(1-0)3个杨树品种均用2年根1年干苗木。其中,毛白杨1316为“一条鞭”嫁接苗,扦插成活接芽(毛白杨1316)生长至30 cm高时,采用高培土方法培育其自生根,次年剪掉砧木萌生根,保留接芽萌生根并平茬,生长1年后待用。欧美107杨和小美旱杨为扦插苗,培育前与毛白杨1316处理一致,培育其新生枝条自生根,次年剪掉扦插原枝条萌生根,保留新枝条萌生根并平茬,生长1年后待用。3个品种杨树苗均于2017年3月进行栽植,采用每品种3重复的随机区组设计,每小区40株;栽植后,对根基部进行截干处理。
2.2 测定指标及方法
在调查1个生长季后的杨树植株地径、树高等指标的基础上,分别于2017-08-01和2017-09-14,每个品种各选3株生长中等的植株进行取样。采用完全取样方式,分根、茎和叶分别进行收获,测定鲜质量(称重法)和叶面积(用北京雅欣理仪科技有限公司生产的Yaxin-1241叶面积仪)后,于105 ℃烘箱中烘干72 h,称取干物质量。此外,在2017-08-30上午09:00-11:00,使用 LI-6400XT便携式光合测量系统(美国LI-COR公司),在光强1 500 μmol/(m2·s) LED人工光源照射下,对3个杨树品种相同生长部位叶片(树干1.5 m)的净光合速率进行测定,并利用SPAD-502 Plus便携式叶绿素测定仪(日本柯尼卡美能达)测定同一叶片的叶绿素含量(SPAD值)。
2.3 数据处理
根据植物生长解析方法[18-19],计算相对生长速率(RGR)、净同化速率(NAR)、比叶面积(LSA)以及各部分(根、茎和叶)同化产物分配比例(P)等指标。各指标计算公式为:
RGR=(lnW2-lnW1 mean)/T;
NAR=(W2-W1 mean)·(lnA2-lnA1 mean)/(A2-A1 mean)T;
LSA=A2/L2;
P=(G2-G1)/(W2-W1)×100%。
式中:W2为第2次取样时的整株生物量(g),W1 mean为第1次取样整株生物量的平均值(g),W1为第1次取样时的整株生物量(g),T为2次取样的间隔时间(d),A2为第2次取样时的总叶面积(m2),A1 mean为第1次取样总叶面积平均值(m2),L2为第2次取样时的叶片总生物量(g),G1为第1次取样时的各部分生物量(g),G2为第2次取样各部分生物量(g)。
将获取的数据利用EXCEL 2003进行整理、计算和制图。利用SPSS 13.0对3个杨树品种的各指标进行单因素方差分析,当差异达显著水平时,采用LSD法进行多重比较。
3 结果与分析
3.1 3个杨树品种的地径、树高及生物量比较
3个杨树品种1个生长季后地径和树高的生长情况见图1。由图1可见,3个杨树品种的地径和树高表现出一定差异,欧美107杨的地径和树高均显著高于小美旱杨和毛白杨1316,毛白杨1316的地径和树高稍高于小美旱杨,但二者差异不显著。
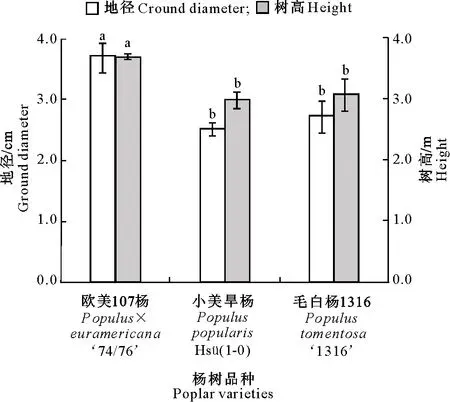
图柱上标不同小写字母表示品种间差异显著(P<0.05),下图同Different small letters indicate significant difference between varieties at the P<0.05 level,the same below图1 3个杨树品种1个生长季后的地径和树高Fig.1 Ground diameter and height of three poplar varieties after one growing season
3个杨树品种的生物量如表1所示。从表1可以看出,与地径、树高生长趋势相似,3个杨树品种的叶片、茎干生物量以及总生物量间也呈现出一定差异,且以欧美107杨最高,而小美旱杨与毛白杨1316差异不显著;3个杨树品种的根系生物量间差异均不显著。

表1 3个杨树品种的生物量Table 1 Biomass of three poplar varieties
3.2 3个杨树品种生长速率及生物量分配的比较
3个杨树品种阶段性生长情况及同化产物的分配情况见表2和图2。从表2可知,2017-08-01-2017-09-14,小美旱杨和毛白杨1316的相对生长速率均较低,二间之间无显著差异,且均显著低于欧美107杨。欧美107杨平均叶面积与毛白杨1316无显著差异,但均显著大于小美旱杨。净同化速率以小美旱杨最高,欧美107杨次之,毛白杨1316最低,三者之间均存在显著差异。

表2 3个杨树品种阶段性生长的比较Table 2 Comparison of stage growth of three poplar varieties

图2 3个杨树品种同化产物的阶段性分配规律Fig.2 Comparison of stage assimilation allocation of three poplar varieties
由图2可知,3个杨树品种向叶片分配的同化产物比例无显著差异,而向茎和根分配的同化产物比例在3个杨树品种间存在差异。相对于欧美107杨而言,小美旱杨和毛白杨1316同化产物向茎的分配比例显著降低,而向根系的分配比例显著升高。
3.3 3个杨树品种叶片生长和光合特性的比较
由表3可知,毛白杨1316总叶面积、单叶面积和比叶面积与欧美107杨无显著差异,但均显著大于小美旱杨;叶片数量则以小美旱杨最高,显著高于另外2个品种。
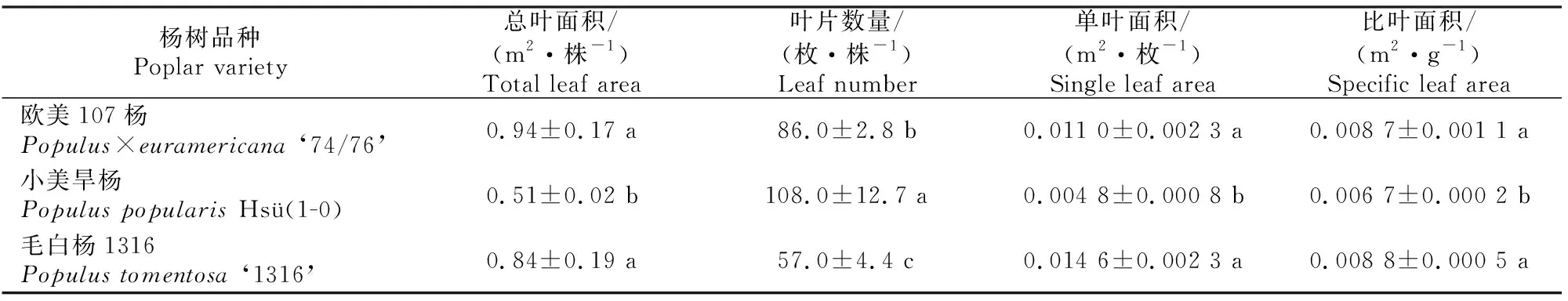
表3 3个杨树品种叶片生长指标的比较Table 3 Comparison of leaf growth of three poplar varieties
图3显示,欧美107杨和小美旱杨净光合速率及叶绿素含量(SPAD值)均显著高于毛白杨1316,说明欧美107杨和小美旱杨单位叶面积的同化能力均显著高于毛白杨1316。
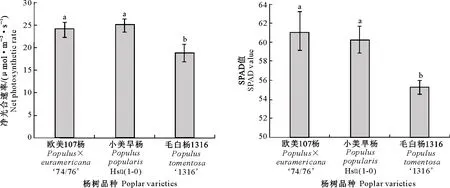
图3 3个杨树品种净光合速率和SPAD值的比较Fig.3 Comparison of net photosynthetic rate and SPAD value of three poplar varieties
4 讨 论
胸径(地径)、树高和生物量均是评价林木生长的重要指标,一般容易受到林分密度[20]、水肥条件[21]等外界环境因素的影响。本研究所选3个杨树品种栽植在同一试验地中,栽植密度、水肥管理等外界因素均保持一致,因此其地径、树高和生物量(叶、茎以及总生物量)间的差异取决于各自的品种特性。本研究发现,欧美107杨的地径、树高和生物量(叶、茎以及总生物量)的增长均显著大于小美旱杨和毛白杨1316,这说明在外界环境条件一致的条件下,杨树品种间地径、树高以及生物量存在一定的正相关。这与黄秦军等[6]对‘渤丰1号’杨和‘渤丰2号’杨的研究结果相一致。
本研究中,在相同生长期内,3个杨树品种的地径、树高生长表现出一定差异,这主要是同化产物生产和分配造成的。与欧美107杨相比,毛白杨1316和小美旱杨的相对生长速率较低;另外,其同化产物向茎分配的比例显著降低,而向根分配的比例显著升高。同化产物的分配主要体现为植物生物量的分配,而植物生物量的分配与其生长时期[22]、年龄[23]、水肥管理[21,24]、环境胁迫[25]等多种因素有关,但根本上仍取决于植物自身的遗传特性[26]。本试验所选的3个杨树品种是我国北方地区普遍栽植的品种,分别代表不同的杨树派别,其遗传特性具有较大差异。
相对生长速率表示植物整体同化产物生产的效率,净同化速率表示光合器官生产物质的效率,二者均是评价植物同化生产效率的重要指标[27]。荀挚峰等[18]认为,相对生长速率取决于净同化速率和叶面积。本研究发现,相较欧美107杨,小美旱杨和毛白杨1316的相对生长速率均较低,这可能主要是因为小美旱杨的叶面积较小,而毛白杨1316的净同化速率较低所致。这与荀挚峰等[18]的观点一致,同时也说明不同杨树品种间相对生长速率存在一定差异,且差异机制并不完全相同。
叶片是植物同化作用的主要场所,直接影响植株有机物质的合成和积累[28]。植物的总叶面积由叶片数量和单叶面积所决定,单叶面积、单株叶片数量越大,其总叶面积就越大,同化产物积累量就越多,植物生长量也就越大[29]。本研究中,与欧美107杨相比,小美旱杨叶片数量虽然较多,但其单叶面积小,最终导致其总叶面积较低。这与麻雪艳等[30]关于干旱条件下玉米总叶面积的研究结论相一致。
净光合速率能够较好地解释净同化速率的变化。有研究表明,净光合速率与净同化速率呈正相关[18],这与本研究结果一致。本研究中,毛白杨1316的净光合速率显著低于欧美107杨和小美旱杨,导致其净同化速率最低。叶绿素含量在一定程度上决定着光合速率的变化[31-32],叶绿素含量(SPAD值)越高,其光合速率越大,二者呈正相关关系[33]。本研究发现,与欧美107杨和小美旱杨相比,毛白杨1316的SPAD值最低,与其净光合速率的表现一致,这验证了张雪悦等[33]的观点,也说明叶绿素含量较低可能是造成毛白杨1316净光合速率较低的原因之一。
5 结 论
1)移栽1个生长季后,3个供试杨树品种的地径、树高、叶片、茎干以及总生物量均表现为欧美107杨显著高于小美旱杨和毛白杨1316,而小美旱杨与毛白杨1316差异不显著。
2)欧美107杨的相对生长速率显著高于小美旱杨和毛白杨1316,欧美107杨和毛白杨1316的平均叶面积均显著高于小美旱杨;三者净同化速率均存在显著差异,表现为小美旱杨>欧美107杨>毛白杨1316。就同化产物分配而言,小美旱杨和毛白杨1316同化产物向根系的分配比例显著高于欧美107杨,向茎的分配比例则显著低于欧美107杨。
3)小美旱杨的总叶面积、单叶面积和比叶面积均显著小于欧美107杨和毛白杨1316,而后两者间差异不显著。但小美旱杨的叶片数量最多,显著高于欧美107杨和毛白杨1316。欧美107杨和小美旱杨的净光合速率及叶绿素含量(SPAD值)均显著高于毛白杨1316。
综上所述,相较欧美107杨,毛白杨1316生长缓慢的主要原因可能是其净光合速率较低,从而导致净同化速率和相对生长速率较低,而叶绿素含量较低可能是造成其净光合速率不高的原因之一。小美旱杨生长缓慢的原因可能是由于单叶面积较小,进而导致总叶面积较小,因此,虽然其具有较高的净光合速率和净同化速率,但相对生长速率无法提高。另外,同化产物向茎的分配比例较低,也可能是导致小美旱杨和毛白杨1316生长缓慢的一个原因。
