光周期对糜子生长发育及叶片内源激素的调控效应
2021-03-08王君杰田翔秦慧彬王海岗曹晓宁陈凌刘思辰乔治军
王君杰,田翔,秦慧彬,王海岗,曹晓宁,陈凌,刘思辰,乔治军
光周期对糜子生长发育及叶片内源激素的调控效应
王君杰,田翔,秦慧彬,王海岗,曹晓宁,陈凌,刘思辰,乔治军
山西农业大学农业基因资源研究中心/农业部黄土高原作物基因资源与种质创制重点实验室/杂粮种质资源发掘与遗传改良山西省重点实验室,太原 030031
【】明确光周期对糜子生理生态指标的影响,为糜子光周期遗传调控网络及相关基因定位奠定基础。通过盆栽试验设置4种光周期处理,选用光敏感性差异不同的3个品种(N1-光钝感-内糜1号,N2-光中间-内糜2号,N3-光敏感-宁糜14号),调查不同处理植株的物候期,测定不同处理抽穗期内源激素含量、成熟期株高、籽粒蛋白质含量和光钝感品种抽穗后的净光合速率。长日照显著延长糜子生育期,增加株高,光敏感品种N3全生育期长日照生育期天数和株高较短日照分别增加62.07%、104.24%,光钝感品种N1为3.51%、33.35%;随着生育进程的推进,抽穗后叶片净光合速率表现出明显差异,延长光照能显著增强糜子叶片净光合速率,增加干物质积累;光周期处理下不同光敏感品种内源激素变化明显,一定浓度IAA促进抽穗,IAA与GA和ZR呈极显著正相关,相关系数分别为0.51和0.40,与ABA呈极显著负相关(-0.62),ABA与GA和ZR呈极显著负相关,分别为-0.70和-0.39,GA与ZR呈极显著正相关(0.47);全生育期短日照条件下不同光敏感品种籽粒蛋白质含量显著高于其他处理,全生育期短日照N1-2、N2-2、N3-2蛋白质含量较全生育期长日照N1-4、N2-4、N3-4分别高18.89%、80.50%和61.23%。光敏感品种较光钝感品种的籽粒蛋白质含量更容易受短日照影响。光周期显著影响不同光敏感性糜子生长发育及内源激素含量,长日照显著延长糜子生育期、增加植株株高和增强叶片净光合速率,但降低籽粒蛋白质含量,一定浓度IAA含量调控糜子由营养生长向生殖生长转变。
光周期;糜子;内源激素;株高;净光合速率;籽粒蛋白质含量
0 引言
【研究意义】糜子抗旱、耐瘠薄,属于短日照作物,对光周期敏感性较其他作物强。光周期贯穿于作物的整个生长发育阶段,光周期变化涉及许多复杂的生理生化过程,针对其变化植株及时作出应答,使植物生长发育处于相对最优的状态[1-2]。对糜子主要生理生化指标的光周期敏感性进行系统和深入的研究,是探索糜子光周期敏感性遗传调控网络、分子机制、相关基因定位和克隆的前提条件。【前人研究进展】作物90%以上干物质的形成主要来源于光合作用[3],Hendriks等[4]研究表明,光周期对植物光合作用影响呈正相关,增加光照时间,光合作用时间延长,导致更多光合产物生成,促进了植物碳水化合物的积累,进而促进植物的生长发育。籽粒蛋白质含量是评价作物品质的主要指标之一,前人通过光周期处理对籽粒蛋白质含量的研究进行了相关报道[5-9]。针对光周期对内源激素等长势指标的研究主要集中在经济作物和大宗作物上,光周期为8和10 h·d-1时,丝瓜幼苗叶片GA3、IAA和ZT的含量较低,在光周期为6和14 h·d-1时较高,但ABA含量变化趋势却相反[10]。Nováková等[11]研究指出烟叶中IAA、ABA、CTK在不同光周期条件下发生不同水平的积累。GA的生物合成直接受光照的调节,在植物的光形态建成、细胞延伸等过程中发挥作用[12-13]。韩天富等[14-15]报道指出,光周期对大豆生育期的调控作用存在于出苗至成熟的全过程,生育期不同的大豆品种在不同光周期处理下的ABA、IAA、ZR和GA具有明显的差异,开花期对早熟品种进行短日处理,明显加快开花后发育进程,降低叶片中的ABA和ZR含量。延长光周期或长日照能显著增加作物的净光合速率[16-17]。冉午玲等[18]研究得出,小麦抽穗后夜间补光能显著增加旗叶GA、IAA、ZR的含量,且激素含量的升高幅度随着补光时间的延长而增加,但ABA含量受影响较小。针对糜子光周期方面的研究比较少,王君杰等[19]研究了糜子不同性状光周期敏感性的综合评价,得出地上鲜重和株高可以作为糜子光周期敏感性主要评价指标性状。董明等[20]研究了不同光照长度对糜子形态建成及幼穗发育的影响,得出每天增加光照和天数有利于干物质的积累;短日照促进糜子从营养生长向生殖生长转变,而长日照抑制向生殖生长转变。陈明江等[21]报道水稻株高的遗传基因主要与内源激素代谢和信号转导有关,表明株高主要受内源激素的调控,这些内源激素包括赤霉素、油菜素内酯和独脚金内酯。【本研究切入点】糜子光敏感性强,不同生态区引种发现株高和抽穗期对光周期响应明显,同时,株高和抽穗期主要受内源激素和生物钟的调控。光周期显著影响作物光合作用,进而影响干物质积累,导致产量和品质发生变化。前人对不同光敏感性糜子的生理生态研究较少,所以系统研究不同光敏感性糜子的株高、叶片净光合速率、内源激素含量和籽粒蛋白质含量对光周期响应机制具有重要意义。【拟解决的关键问题】本研究选用光周期敏感性不同的3个品种为研究对象,研究不同光周期处理下糜子抽穗期叶片生长素(IAA)、赤霉素(GA)、脱落酸(ABA)和玉米素核苷(ZR)含量变化及株高、光合性能、籽粒蛋白质含量等生长发育及品质变化,揭示不同光敏感性品种内源激素含量影响糜子抽穗的主要代谢途径,为不同生态区糜子引种和育种家选育优异品种提供数据参考。
1 材料与方法
1.1 试验材料
选用光周期敏感性不同的3个糜子品种,分别为光周期钝感性N1(内糜1号)、光周期中间性N2(内糜2号)和光周期敏感性N3(宁糜14号)。
1.2 试验方法
试验于2019年在忻州定襄良种场(38°33′N,112°54′E)进行,海拔780 m,年降水量430 mm,年均气温8.7℃,无霜期158 d左右,年日照时数达2 734.6 h,温、光、热资源适宜糜子的生长发育。采用盆栽方法,每盆播种30粒种子,每处理2盆。设4种光照处理(表1),出苗后每盆选留生长一致的幼苗15株,3叶期开始进行光周期处理。其中,短日照处理光照长度为8 h(晚上17:00至次日早晨9:00置于暗室,其余时间置于自然光照下),长日照处理光照长度为16 h(晚上22:00至次日早晨6:00置于暗室,其余时间置于自然光照下,光照时间不够用白炽灯补充,白炽灯光照强度为25 µmol·m-2·s-1)。抽穗期开始每隔10 d选取长势一致的不同光敏感性植株倒二叶测定其净光合速率,重复3次,同时摘取不同光敏感性品种的倒二叶用液氮速冻后放置-80℃的冰箱里,待后期测定生长素(IAA)、赤霉素(GA)、脱落酸(ABA)和玉米素核苷(ZR),其中抽穗期测定的内源激素含量用于分析影响糜子抽穗的主要代谢途径,其余测定的内源激素含量用于进行激素间相关性分析。

表1 不同光周期处理
1.3 测定指标与方法
1.3.1 内源激素 采用酶联免疫检测技术(ELISA)测定糜子叶片的生长素(IAA)、赤霉素(GA)、脱落酸(ABA)和玉米素核苷(ZR)。
1.3.2 净光合速率 采用便携手持式光合仪CI-340测定糜子植株倒二叶的净光合速率(n),选择在上午9:00—11:00的天气晴朗、无风或风速很小的天气条件下进行。
1.3.3 株高 成熟期选取5株长势一致的植株,用剪刀剪去根部,用直尺测定植株基部至穗顶部的长度,即为株高,取均值为该性状的最终值。
1.3.4 籽粒蛋白质 成熟期选取10株长势一致的主茎穗,自然晾干脱粒、脱壳和粉碎,用全自动凯氏定氮仪[22]测定籽粒氮含量,籽粒蛋白质含量(%)=籽粒氮含量×6.25。
1.4 数据处理
采用DPS和Excel统计软件对糜子株高、籽粒蛋白质含量、n、IAA、GA、ABA和ZR进行方差、相关性分析和作图分析。
2 结果
2.1 不同光周期处理对株高的影响
光钝感糜子品种N1在不同光周期处理下的变化趋势与N2和N3有一定差异,N1的株高表现为N1-4>N1-1>N1-3>N1-2(图1-A),抽穗前短日照N1-1株高较N1-2高17.83%,抽穗前长日照N1-4株高较N1-3高20.95%,抽穗前短日照抽穗后长日照N1-1株高较抽穗前长日照抽穗后短日照N1-3高6.87%,全生育期长日照N1-4的株高较全生育期短日照N1-2高33.35%。N2和N3的株高变化趋势均表现为N-4>N-3>N-1>N-2(图1-B、C),抽穗前短日照N2-1、N3-1株高较N2-2、N3-2分别高11.19%和31.82%,抽穗前长日照N2-4、N3-4株高较N2-3、N3-3分别高5.65%和6.49%,抽穗前长日照抽穗后短日照N2-3、N3-3株高较抽穗前短日照抽穗后长日照N2-1、N3-1分别高42.91%和45.50%,全生育期长日照N2-4、N3-4的株高较全生育期短日照N2-2、N3-2分别高67.88%和104.24%。光敏感糜子品种N3-2株高(65.51cm)低于光钝感品种N1-2(67.44cm),进一步说明光钝感品种受短日照影响小。
2.2 不同光周期处理对生育期的影响
不同光周期敏感性糜子品种在处理1和处理2下,生育期的天数都相同,品种间表现为N3=N2>N1,说明抽穗前糜子对短日照敏感性强,抽穗后对长日照不敏感,在光周期处理2下,N3-2的生育期天数高于N1-2 3.51%(表2)。在处理3和处理4下,同一品种的生育期差异不显著,但光敏感品种显著高于光中性和光钝感品种,表现为N3>N2>N1,在光周期处理4下,N3-4的生育期天数高于N1-4 62.07%。不同光周期敏感性品种在抽穗前长日照下生育期天数高于抽穗前短日照,其中光钝感品种N1差异不明显,光中间型和光敏感品种差异显著。

不同的小写字母表示5%水平下差异显著。下同 different small letters are significantly different at 5% probability level. The same as below

表2 不同处理对生育期的影响
2.3 不同光周期处理对净光合速率的影响
以光钝感糜子品种N1为例,研究抽穗后不同光周期处理对植株叶片净光合速率的影响(图2)。处理N1-1和N1-3随着生育进程的推进呈现抛物线的变化趋势,处理N1-2和N1-4随着生育进程的推进呈现逐渐下降的变化趋势。在7月27日糜子抽穗10 d后,全生育期长日照N1-4净光合速率显著高于其他处理,抽穗前短日照N1-2净光合速率较N1-1高3.60%,抽穗前长日照N1-4净光合速率较N1-3高10.47%,抽穗前长日照抽穗后短日照N1-3净光合速率较抽穗前短日照抽穗后长日照N1-1高17.42%,全生育期长日照N1-4的净光合速率较全生育期短日照N1-2高25.20%。在8月6日抽穗20 d和8月16抽穗30 d后,抽穗后转置到长日照N1-1的净光合速率显著高于其他处理,分别为28.77和18.53 μmol·m-2·s-1,说明延长光照能显著增强糜子叶片净光合速率,从而积累更多干物质;抽穗前长日照N1-3净光合速率较N1-4分别高42.36%和9.12%,这与抽穗10 d的变化趋势不同,说明随着成熟期的推进,植株为了适应新的环境,防止过早衰落,通过自生调节能力,增强叶片净光合速率,从而增加干物质的积累量。
2.4 不同光周期处理对抽穗期内源激素含量的影响
由表3可以看出,光钝感糜子品种N1在抽穗期(N1-1和N1-2在7月17日抽穗,N1-3和N1-4在7月18日抽穗)ABA、GA和ZR均表现为抽穗前长日照N1-3和N1-4显著高于抽穗前短日照N1-1和N1-2,IAA含量表现为抽穗期显著高于抽穗前,说明IAA含量增加到一定浓度促进植株抽穗。7月24日抽穗时,光敏感中间性糜子品种N2在3叶期—抽穗期短日照处理下(N2-1、N2-2)ABA、GA和ZR含量显著低于N2-3和N2-4,IAA变化趋势相反;7月31日抽穗时,3叶期—抽穗期长日照处理下(N2-3、N2-4)IAA和ZR含量显著高于N2-1和N2-2,ABA和GA变化趋势相反,表明IAA积累到一定浓度,即能促进植株抽穗。N2-1(7月31日)处理的IAA、GA和ZR含量高于N2-2,ABA显著低于N2-2,说明抽穗后延长光照有利于IAA、GA和ZR积累,减低ABA积累,从而促进植株生长。7月24日抽穗时,光敏感性糜子品种N3在3叶期—抽穗期短日照处理下(N3-1、N3-2)IAA和ABA含量显著高于N3-3和N3-4,GA和ZR显著低于N3-3和N3-4;8月11日抽穗时,3叶期—抽穗期长日照处理下(N3-3、N3-4)IAA含量显著高于N3-1和N3-2,GA变化趋势相反,同理说明IAA是影响糜子抽穗的主要物质。N3-1(8月11日)处理的GA和ABA含量显著高于N3-2,IAA显著低于N3-2,说明抽穗后延长光照有利于GA和ABA积累,减低IAA积累,从而促进植株生长发育。可见不同光敏感性糜子品种在抽穗前后长短日照交替影响内源激素的机制是具有差异的。
内源激素间相关性分析可知,IAA与GA和ZR呈极显著正相关,与ABA呈极显著负相关;ABA与GA和ZR呈极显著负相关;GA与ZR呈极显著正相关(表4)。
2.5 不同光周期处理对籽粒蛋白质的影响
不同光周期敏感性品种在不同处理下籽粒蛋白质含量变化趋势不一致(图3)。全生育期短日照N-2籽粒蛋白质含量显著高于N-1、N-3和N-4。光钝感糜子品种N1在不同光周期处理下,抽穗后短日照N1-2、N1-3籽粒蛋白质含量较抽穗后长日照N1-1、N1-4分别高8.14%和18.89%,全生育期短日照N1-2蛋白质含量较全生育期长日照N1-4高18.89%。N2和N3在不同光周期处理下,抽穗前短日照N-2籽粒蛋白质含量显著高于抽穗前长日照N-1,N2-2较N2-1高2.78%,N3-2较N3-1高21.86%,说明营养生长阶段植株通过短日照更有利于氮素的积累,后期把更多的氮素营养转移到籽粒中,从而提高籽粒蛋白质含量。全生育期短日照N2-2、N3-2籽粒蛋白质含量较全生育期长日照N2-4、N3-4分别高80.50%和61.23%,说明光敏感品种较光钝感品种的籽粒蛋白质含量更容易受短日照影响。

图2 不同处理对叶片净光合速率的影响

表3 不同处理对内源激素的影响
不同的小写字母表示 5%水平下差异显著different small letters are significantly different at 5% probability level
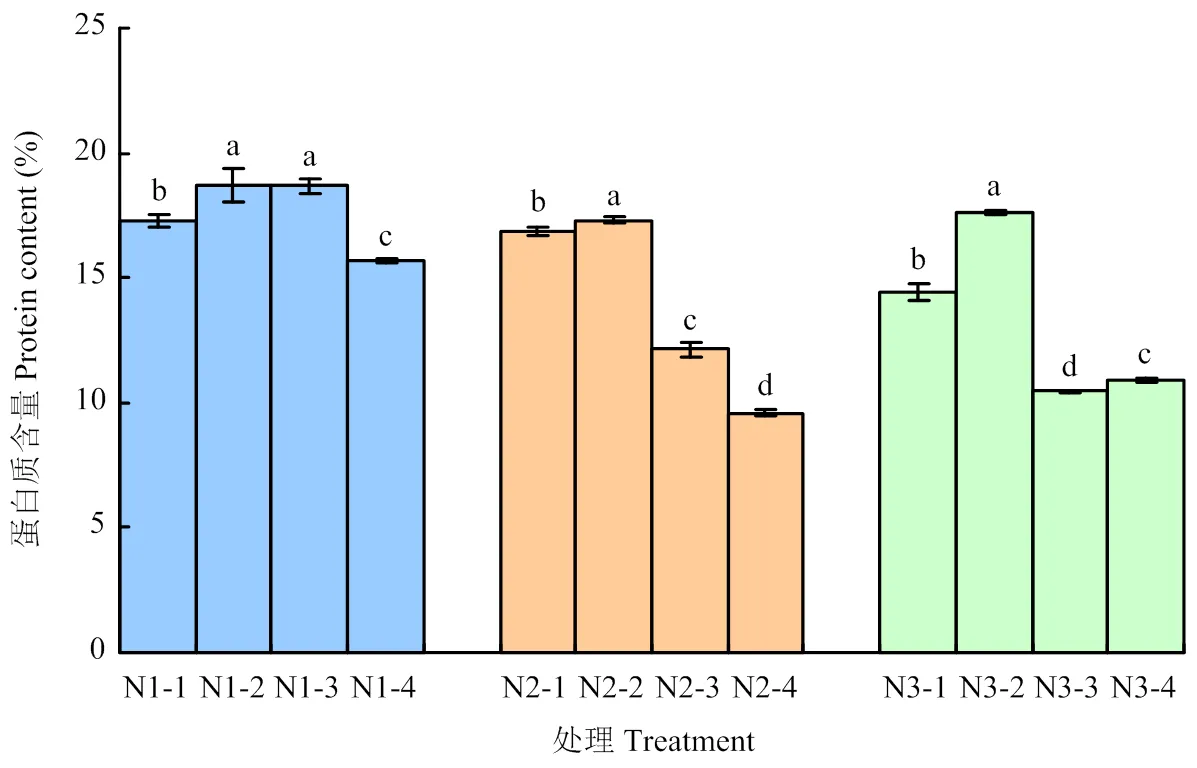
图3 不同处理对籽粒蛋白质含量的影响
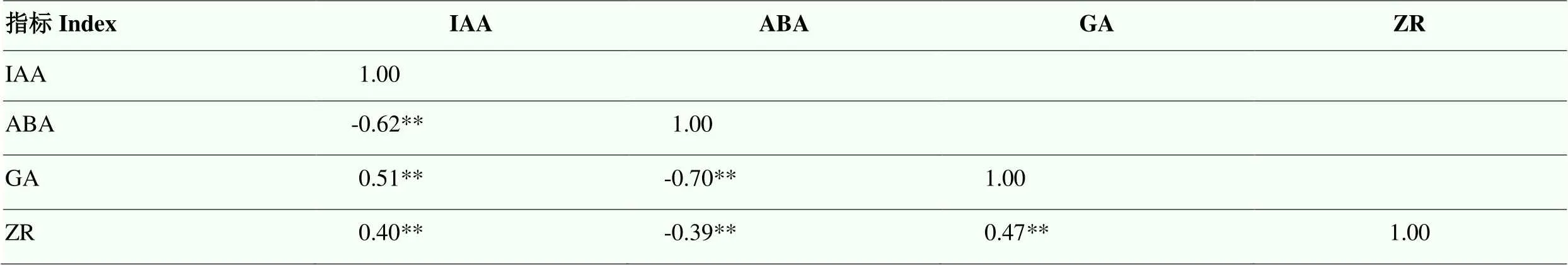
表4 内源激素间的相关性分析(n=210)
**和*分别表示1%和5%水平上差异显著
* and ** indicate significant correlations at 0.05 and 0.01 levels, respectively
3 讨论
3.1 株高的光周期敏感性
株高是构成植株的主要性状之一,适宜的株高能大幅度提高作物产量。20世纪60年代,矮秆水稻的选育使产量提高20%—30%[23]。株高主要受内源激素的调控,赤霉素、油菜素内酯和独脚金内酯作为重要的植物促生激素,彼此存在着相互协同作用,参与调控了作物株高的生理生化过程。Bai等[24]研究发现,PRE、IBH1和HBI1这3个蛋白中任何一个发生变化都会影响拟南芥对BR、GA、温度及光照的敏感性,从而影响株高的伸长。糜子较其他作物对光周期敏感,短日照加快生殖生长的进程,长日照延长营养生长的进程,同一品种在不同生态区株高有显著差异,海南植株株高仅为10 cm左右。沈韫赜等[6]研究表明,小麦株高随着开花前光周期(12、16、20和24 h)的延长而减小,开花后对株高影响不显著。庞琳娜等[25]通过冬小麦拔节期开始遮光直至收获,发现不同程度遮光对冬小麦株高影响不明显。这与本试验结果基本一致,由于小麦为长日照作物,糜子为短日照作物,所以光周期影响株高的变化趋势相反,本试验研究得出3个不同光敏感性糜子品种抽穗前株高随着光照时间的延长而增大,均表现为N-4>N-1。王英等[9]发现,初花后长日照16 h可增加大豆株高的长度。前人通过水稻移栽后开始遮光,出穗后停止遮光,发现抽穗前株高明显高于对照,出穗后遮光处理的株高明显低于对照[26-27]。本试验研究结果表明,不同光敏感型糜子品种全生育期长日照能显著增加株高长度,光钝感糜子品种株高变化趋势与其他2个不同光敏感性品种有一定差异,抽穗后长日照株高N1-1高于抽穗后短日照N1-3,而N2、N3品种抽穗前长日照株高显著高于抽穗后长日照,说明光敏感品种在营养生长阶段对光周期敏感性较生殖生长阶段强,而光钝感品种变化相反,可能是由于光钝感品种通过增加株高截获更多光能,调节内源激素含量比例增强自身适应能力,以适应长日照逆境对其影响。有关光周期处理下内源激素对不同光敏感性糜子株高的调控机理有待进一步研究。
3.2 净光合速率的光周期敏感性
叶片是植株进行光合作用的主要部位,光周期显著影响植株光合速率,一定范围内光照时间延长,叶片光合速率增强,从而植株积累更多碳水化合物,进而促进植株生长发育[28]。冉午玲等[18]研究指出,小麦抽穗后叶片净光合速率随着夜间人工补光时间的延长而增加,至抽穗后35 d,各处理间无显著差异。本试验测定了光钝感品种开花后10、20和30 d的净光合速率,得出30 d净光合速率除N1-1外其他处理差异不显著。王亚江等[29]研究发现,齐穗后10、20和30 d均呈现抽穗后遮光叶片净光合速率显著低于抽穗前遮光,这与本试验结果有一定差异。植物对光的适应通常表现在光合特性和生物化学适应性的改变,可能是由于糜子光敏感性强,抽穗后植株通过调节自身生理生化反应来更好地适应长短日照逆境对其影响。刘杰等[16]研究发现,延长光周期能显著提高水培生菜的净光合速率,以250 µmol·m-2·s-1光强和16 h·d-1光周期组合最有利于产量和光能利用。孙善军等[30]研究了初花期遮阴对2个绿豆品种光合特性的影响,发现遮光30%时,大花叶子绿豆的净光合速率较全光照显著下降了24%,小槐花园叶绿豆无显著变化,但遮光72%时,2种绿豆的净光合速率均受到显著抑制。这与本试验结果基本一致,说明延长光照能显著增加糜子叶片净光合速率,从而积累更多有机物质,进而提高产量。
3.3 抽穗期内源激素的动态变化
内源激素对植物的生长发育具有重要的调节作用,同时,不同植物激素对光周期的响应各不相同,但其信号通路之间却存在互作关系,从而形成复杂的网络状调控路径[31]。抽穗期是鉴别营养生长和生殖生长的关键时期,董伟欣等[32]研究得出短日照诱导使小豆开花期和成熟期提前,是由于叶片内的ABA和ZR含量升高,GA和IAA减低所导致;林贵玉等[33]研究得出,短日照促进菊花花芽分化,是由于CTK和ABA含量升高,GA和IAA减低;樊文娜等[34]研究认为,短日照条件下苜蓿ABA含量最大,GA含量最低,袁娟等[35]研究得出短日照处理使扁豆GA形成高峰时间提前,ABA时间延后,其研究结果都是短日照促进ABA的合成,减低GA含量。Dong等[36]研究发现短日照条件下红小豆植株GA和ABA含量减低,这与前面研究结果不一致。本试验研究得出不同光敏感性糜子品种内源激素调控抽穗的代谢途径具有差异,短日照促进光钝感和光中间型品种抽穗期(N1、N2)IAA含量增加,减低ABA、GA和ZR含量;促进光敏感性品种抽穗期(N3)IAA和ABA含量增加,减低GA和ZR含量。造成差异的原因可能是由于不同敏感性品种对内源激素的敏感度不一样,同时光周期只是影响内源激素变化的某一种因子,不同内源激素在植物生长发育过程中发挥不同的作用,并且存在着复杂的交互作用。
3.4 籽粒蛋白质的光周期敏感性
蛋白质含量多少是衡量作物品质优良的主要指标之一,前人对光周期影响籽粒蛋白质的研究已有大量报道,王英等[9]研究指出,大豆长日照16 h与短日照12 h相比,能显著减低籽粒蛋白质含量。韩天富等[7]研究阐释了光周期对大豆籽粒品质有很大影响,长日照条件下能显著减低籽粒蛋白质含量,增加脂肪含量。贾士芳等[37]研究得出遮光处理使玉米籽粒蛋白质和粗脂肪含量显著提高,这与本试验的研究结果一致,本试验得出不同光敏感性品种在全生育期短日照的籽粒蛋白质含量显著高于其他处理。说明同一品种在不同生态区籽粒营养品质具有明显的差异,前人研究得出水稻蛋白质含量与稻米食味品质呈极显著负相关,随着水稻籽粒蛋白质含量提高的同时,也降低了米饭的食用品质[38],所以在不影响籽粒食味品质的前提下适当提高其蛋白质含量,从而增加其营养品质。根据糜子籽粒蛋白质含量对光周期敏感性的响应机理,可以优化作物布局和籽粒品质分级,培育适合不同生态区种植的优质高产糜子品种。
4 结论
长日照显著延长糜子生育期、增加植株株高、增强叶片净光合速率和降低籽粒蛋白质含量;光钝感品种主要通过IAA激素达到一定浓度时调控糜子抽穗,光中间型品种主要通过增加IAA含量、减低ABA和GA含量来调控糜子抽穗,光敏感品种主要通过增加IAA含量、减低GA含量来调控糜子抽穗。不同光敏感性品种性状指标的变化趋势受光周期影响具有一定差异性。
[1] 韩天富, 马凤鸣, 王金陵, 魏湜. 光周期对大豆叶片内源激素含量及其平衡的影响. 作物学报, 1996, 22(6): 661-667.
Han T F, Ma F M, Wang J L, Wei S. Photoperiodic effects on the amount and balance of endogenous hormones in soybean leaves. Acta Agronomica Sinica, 1996, 22(6): 661-667. (in chinese)
[2] Chory J, Wu D. Weaving the complex web of signal transduction. Plant Physiology, 2001, 125(1): 77-80.
[3] Donald C M. The breeding of crop ideotypes. Euphytica1969, 17(3): 385-403.
[4] Hendriks J H M, Kolbe A, Gibon Y, GeigenbergeR P. ADP-glucose pyrophosphorylase is activated by posttranslational redox-modification in response to light and to sugars in leaves ofand other plant species. Plant Physiology, 2003, 133(2): 838-849.
[5] 邢阿宝, 崔海峰, 俞晓平, 张雅芬, 叶子弘. 光质及光周期对植物生长发育的影响. 北方园艺, 2018(3): 163-172.
Xing A B, Cui H F, Yu X P, Zhang Y F, Ye Z H.Effects of different lights qualities and photoperiods on plant growth and development. Northern Horticulture, 2018(3): 163-172. (in chinese)
[6] 沈韫赜, 郭双生. 受控环境下光周期对小麦生长发育、产量及营养品质的影响. 麦类作物学报, 2015, 35(1): 64-70.
Shen Y Z, Guo S S. Effects of photoperiod on growth, development, yield and nutritional quality of wheat in controlled system. Journal of Triticeae Crops, 2015, 35(1): 64-70. (in chinese)
[7] 韩天富, 王金陵, 杨庆凯, 盖钧镒. 开花后光照长度对大豆化学品质的影响. 中国农业科学, 1997, 30(2): 47-53.
Han T F, Wang J L, Yang Q K, Gai J Y. Effects of post-flowering photoperiod on chemical composition of soybeans. Scientia Agricultura Sinica, 1997, 30(2): 47-53. (in chinese)
[8] 杨俊华, 唐李军, 陈以相, 张慧, 王青, 杨亚军, 覃鹏. 弱光对小麦产量性状及品质特性的影响. 西南农业学报, 2019, 32(2): 291-295.
Yang J H, Tan L J, Chen Y X, Zhang H, Wang Q, Yang Y J, QIn P. Effects of weak light on yield traits and quality characteristics of wheat. Southwest China Journal of Agricultural Sciences, 2019, 32(2): 291-295. (in chinese)
[9] 王英, 程立锐, 冷建田, 吴存祥, 候文胜, 韩天富. 开花后不同光周期条件下大豆农艺性状和品质性状的QTL分析. 作物学报, 2010, 36(7): 1092-1099.
Wang Y, Cheng L R, Leng J T, Wu C X, Hou W S, Han T F. QTL mapping of agronomic and quality traits in soybean under different post-flowering photoperiods. Acta Agronomica Sinica2010, 36(7): 1092-1099. (in chinese)
[10] 吴雪霞, 尚静, 张圣美, 朱宗文, 张爱冬, 查丁石. 不同光周期对丝瓜幼苗生长生理及其内源激素含量和性别分化的影响. 西北植物学报, 2019, 39(10): 1812-1818.
Wu X X, Shang J, Zhang S M, Zhu Z W, Zhang A D, Cha D S. Effect of different photoperiods on plant growth, contents of endogenous hormones and floral sex differentiation of luffa cylindrica in seeding stage. Acta Botanica Boreali-Occidentalia Sinica, 2019, 39(10): 1812-1818. (in chinese)
[11] Nováková M, Motyka V, Dobrev P I, Malbeck J, Gaudinová R. Diurnal variation of cytokinin, auxin and abscisic acid levels in tobacco leaves. Journal of Experimental Botany, 205, 56: 2877-2883.
[12] Foster K R, Morgan P W. Genetic regulation of developmemt in sorghum bicolor. Plant Physiology, 1995, 108(1): 337-343.
[13] Cowling R J, Harberd N P. Gibberellins control arabidopsis hypocotyl growth via regulation of cellular elongation. Journal of Experimental Botany, 1999, 50: 1351-1357.
[14] 韩天富, 盖钧镒. 大豆几种光周期处理效应的植物激素解析. 作物学报, 1999, 25(3): 349-355.
Han T F, Gai J Y. Phytohormonal analysis of some photoperiod effects in soybean. Acta Agronomica Sinica, 1999, 25(3): 349-355. (in chinese)
[15] 韩天富, 王金陵. 大豆开花后光周期反应的研究. 植物学报, 1995, 37(11): 863-869.
Han T F, Wang J L. Sdudies on the post-flowering photoperiodic responses in soybean. Acta Botanica Sinian, 1995, 37(11): 863-869. (in chinese)
[16] 刘杰, 胡笑涛, 王文娥, 冉辉, 方舒玲, 杨鑫. 光强和光周期对水培生菜光合及叶绿素荧光特性的影响. 西南农业学报, 2019, 32(8): 1784-1790.
Liu J, Hu X T, Wang W E, Ran H, Fang S L, Yang X. Effects of light intensity and photoperiod on photosynthetic characteristics and chlorophyⅡfluorescence of hydroponic lettuce. Southwest China Journal of Agricultural Sciences, 2019, 32(8): 1784-1790. (in chinese)
[17] 李冬梅, 谭秋平, 高东升, 陈修德, 李玲. 光周期对休眠诱导期桃树光合及PSⅡ光系统性能的影响. 应用生态学报, 2014, 25(7): 1933-1939.
Li D M, Tan Q P, Gao D S, Chen X D, Li L. Effects of photoperiod on photosynthesis and PSⅡperformance in peach during dormancy induction. Chinese Journal of Applied Ecology, 2014, 25(7): 1933-1939. (in chinese)
[18] 冉午玲, 郭家萌, 马野, 毛俊, 韩心培, 郑博元, 李东亮, 邵瑞鑫, 杨青华. 夜间补光对小麦叶片激素含量及光合特性的影响. 麦类作物学报, 2017, 37(9): 1181-1186.
Ran W L, Guo J M, Ma Y, Mao J, Han X P, Zheng B Y, Li D L, Shao R X, Yang Q H. Effect of supplemental illumination at night on endogenous plant hormones content and photosynthetic characteristics in wheat leaves. Journal of Triticeae Crops, 2017, 37(9): 1181-1186. (in chinese)
[19] 王君杰, 王海岗, 曹晓宁, 陈凌, 刘思辰, 田翔, 秦慧彬, 乔治军. 糜子不同性状光周期敏感性的综合评价. 中国农业科学, 2020, 53(3): 474-485.
Wang J J, Wang H G, Cao X N, Chen L, Li S C, Tian X, Qin H B, Qiao Z J. Comprehensive evaluation of photoperiod sensitivity based on different traits of broomcorn millet. Scientia Agricultura Sinica, 2020, 53(3): 474-485. (in chinese)
[20] 董明, 降彦苗, 李海权, 耿玲玲, 刘建烨, 乔志红, 刘国庆. 光周期变化对糜子形态建成及幼穗发育进程的影响. 中国农业科学, 2020, 53(6): 1118-1125.
Dong M, Jiang Y M, Li H Q, Geng L L, Liu J Y, Qiao Z H, Liu G Q. Effect of photoperiod changes on morphological characters and young panicle development in Proso Millet. Scientia Agricultura Sinica, 2020, 53(6): 1118-1125. (in chinese)
[21] 陈明江, 刘贵富, 余泓, 王冰, 李家洋. 水稻高产优质的分子基础与品种设计. 科学通报, 2018, 63(14): 1276-1289.
Chen M J, Liu G F, Yu H, Wang B, Li J Y. Towards molecular design of rice plant architecture and grain quality. Chinese Science Bulletin, 2018, 63(14): 1276-1289. (in chinese)
[22] 闵良, 姚文华, 徐国良, 王剑飞, 母其爱. 全自动凯氏定氮仪测定复合肥料中的总氮含量. 湖北农业科学, 2012, 51(1): 175-177.
Min L, Yao W H, Xu G L, Wang J F, Mu Q A. Determination of the nitrogen content of compound fertilizer by Kjeltec automatic azotometer. Hubei Agricultural Sciences, 2012, 51(1): 175-177. (in chinese)
[23] Peng J R, Richaids D E, Hartley N M, Murphy G P, Devos K M, Flintham J E, Beales J, Fish L J, Worland A J, Pelica F. “Green revolution” gene encode mutant gibberellins response modulators. Nature1999, 400: 256-261.
[24] Bai M Y, Min F, Eunkyoo O, Wang Z Y. A triple Helix- Loop-Helix/Basic Helix-Loop-Helix cascade controls cell elongation downstream of multiple hormonal and environmental signaling pathways in Arabidopsis. Plant Cell, 2012, 24(12): 4917-4929.
[25] 庞琳娜, 史文娟, 张建丰, 文利军, 李涛. 不同遮光度对冬小麦生长发育和水分利用效率的影响. 水土保持通报, 2018, 38(4): 53-59.
Pang L N, Shi W J, Zhang J F, Wen L J, Li T. Effects of different shading treatments on growth and water use efficiency of winter wheat. Bulletin of Soil and Water Conservation, 2018, 38(4): 53-59. (in chinese)
[26] 闫萌萌, 闫平, 吕守义, 吴金荣, 刘福英, 邢超. 遮光对水稻生长发育及稻米品质的影响. 农学学报, 2019, 9(4): 22-25.
Yan M M, Yan P, Lü S Y, Wu J R, Liu F Y, Xing C. Effect of shading on rice growth and quality. Journal of Agriculture, 2019, 9(4): 22-25. (in chinese)
[27] 徐波, 王宝祥, 邢运高, 孙志广, 杨波, 刘艳, 卢百关, 徐大勇. 遮光对水稻生长发育及稻米品质的影响. 热带农业工程, 2019, 43(1): 19-20.
Xu B, Wang B X, Xing Y G, Sun Z G, Yang B, Liu Y, Lu B G, Xu D Y. Effects of shading on rice growth and development and rice quality. Tropical Agricultural Engineering, 2019, 43(1): 19-20. (in chinese)
[28] Hendriks J H, Kolbe A, Gibon Y, Stitt M, Geigenberger P. ADP-glucose pyrophosphorylase is activated by posttranslational redox-modification in response to light and to sugars in leaves of Arabidopsis and other plant species. Plant Physiology, 2003, 133(2): 838-849.
[29] 王亚江, 颜希亭, 孟天瑶, 杨大柳, 魏海燕, 张洪程, 戴其根, 霍中洋, 许轲. 抽穗前后遮光对超级粳稻产量形成的影响. 中国稻米, 2014, 20(5): 18-21.
Wang Y J, Yan X T, Meng T Y, Yang D L, Wei H Y, Zhang H C, Dai Q G, Huo Z Y, Xu K. Effects of shading before and after heading on yield formation of super japonica rice. China Rice, 2014, 20(5): 18-21. (in chinese)
[30] 孙善军, 邹长明, 张晓红, 赵敏, 何涛. 遮阴对两个绿豆品种光合作用和生长发育的影响. 草业科学, 2017, 34(6): 1247-1254.
Sun S J, Zou C M, Zhang X H, Zhao M, He T. Effect of shading on photosynthesis and growth in two phaseolus aureus varieties. Pratacultural Science2017, 34(6): 1247-1254. (in chinese)
[31] 帅海威, 孟永杰, 陈锋, 周文冠, 罗晓峰, 杨文钰, 舒凯. 植物遮阴胁迫的激素信号响应. 植物学报, 2018, 53(1): 139-148.
Shuai H W, Meng Y J, Chen F, Zhou W G, Luo X F, Yang W Y, Shu K. Phytohormone-mediated plant shade responses. Chinese Bulletin of Botany, 2018, 53(1): 139-148. (in chinese)
[32] 董伟欣, 路战远, 任帅, 张彦立, 谢颖, 刘明, 魏岩, 张月辰. 短日照诱导对小豆叶片内源激素含量及其平衡的影响. 河南农业大学学报, 2015, 49(6): 723-728, 736.
Dong W X, Lu Z Y, Ren S, Zhang Y L, Xie Y, Liu M, Wei Y, Zhang Y C. Effects of short-day photoperiod on hormone contents and balance of endogenous hormones in adzuki bean leaves. Journal of Henan Agricultural University, 2015, 49(6): 723-728, 736. (in chinese)
[33] 林贵玉, 郑成淑, 孙宪芝, 王文莉. 光周期对菊花花芽分化和内源激素的影响. 山东农业科学, 2008, 1: 35-39.
Lin G Y, Zheng C S, Sun X Z, Wang W L. Effects of photoperiod on floral bud differentiation and contents of endogenous hormones in chrysanthemum. Shangdong Agricultural Sciences, 2008, 1: 35-39. (in chinese)
[34] 樊文娜, 孙晓格, 倪俊霞, 杜红旗, 史莹华, 严学兵, 王成章. 光周期对不同秋眠型苜蓿光敏色素和内源激素的影响. 草业学报, 2014, 23(1): 177-184.
Fan W N, Sun X G, Ni J X, Du H Q, Shi Y H, Yan X B, Wang C Z. Effect of photoperiod on phytochromes and endogenous hormones of alfalfa with different fall-dormancies. Acta Prataculturae Sinica, 2014, 23(1): 177-184. (in chinese)
[35] 袁娟, 武天龙, 陈典. 光周期对扁豆真叶内源激素及游离氨基酸含量的影响. 上海交通大学学报, 2004, 22(3): 215-219, 216.
Yuan J, Wu T L, Chen D. Effect of photoperiodic treatment on the endogenous hormones and dissociative amino acid of hyacinth bean leaves. Journal of Shanhai Jiaotong University, 2004, 22(3): 215-219, 216. (in chinese)
[36] Dong W X, Zhang Y Y, Zhang Y L, Ren S, Wei Y, Zhang Y C. Short-day photoperiod effects on plant growth, flower bud differentiation, and yield formation in adzuki bean. International Journal of Agriculture&Biology, 2016, 18(2): 337-345.
[37] 贾士芳, 李从锋, 董树亭, 张吉旺. 花后不同时期遮光对玉米粒重及品质影响的细胞学研究. 中国农业科学, 2010, 43(5): 911-921.
Jia S F, Li C F, Dong S T, Zhang J W. Effects of shading at different stages after anthesis on maize grain weight and quality at cytology level. Scientia Agricultura Sinica, 2010, 43(5): 911-921. (in chinese)
[38] 张春红, 李金州, 田孟祥, 王才林. 不同食味粳稻品种稻米蛋白质相关性状与食味的关系. 江苏农业学报, 2010, 26(6): 1126-1132.
Zhang C H, Li J Z, Tian M X, Wang C L. Relationship between protein-related traits and palatabilities of japonica rice (L. ) with distinct taste. Jiangsu Journal of Agricultural Sciences, 2010, 26(6): 1126-1132. (in chinese)
regulation effects of photoperiod on growth and leaf endogenous hormones in Broomcorn Millet
WANG JunJie, TIAN Xiang, QIN HuiBin, WANG HaiGang, CAO XiaoNing, CHEN Ling, LIU SiChen, QIAO ZhiJun
Center for Agricultural Genetic Resources Research, Shanxi Agricultural University/Key Laboratory of Crop Gene Resources and Germplasm Enhancement on Loess Plateau, Ministry of Agriculture/Shanxi Key Laboratory of Genetic Resources and Genetic Improvement of Minor Crops, Taiyuan 030031
【】In this study, the variation law of photoperiod on physiology and ecology index of broomcorn millet were observed, which would facilitate to develop the genetic regulation network and map related genes of photoperiod.【】The experiments were conducted using 3 broomcorn millet varieties (N1-light insensitive-Neimi 1, N2-light middle-Neimi 2, and N3-light sensitive-Ningmi 14) with different light insensitive degree and with four photoperiod treatments in the pot. The phenological period was investigated. The endogenous hormone content at heading stage, plant height and grain protein content at mature period, and net photosynthetic rate of light insensitive variety after heading were measured.】The growth period and plant height of broomcorn millet were significantly increase by long day compared with short day, which in light sensitive material N3 was 62.07% and 104.24% higher than that of short day, and light insensitive material N1 was 3.51% and 33.35%, respectively. The net photosynthetic rate of leaves after heading showed a significant difference with the development of the growth process, the net photosynthetic rate increased significantly with the prolonged light for accumulating more dry matter. Different photosensitive materials had different regulating mechanisms of endogenous hormones under different photoperiod treatments. The accumulation of IAA content to a certain concentration could promote the plant heading. IAA was significantly positively correlated with GA and ZR, with correlation coefficients of 0.51 and 0.40, respectively, while it was significantly negatively correlated with ABA, with the value of -0.62. The ABA content was negatively correlated with GA and ZR, with the value of -0.70 and -0.39, respectively, and the GA content was positively correlated with ZR, with the values of 0.47. The grain protein content of different light-sensitive materials under the short day in the whole growth period was significantly higher than that of other treatments. The grain protein content of N1, N2 and N3 under short day in the whole growth period were 18.89%, 80.50% and 61.23% higher than those of long day in the whole growth period, respectively, indicating that the light-sensitive material was more susceptible to short-day sunshine than the light-insensitive material.【】The photoperiod significantly affected the growth and endogenous hormone content in different photosensitivity materials. Long day significantly prolonged the growth period, increased plant height and enhanced leaf net photosynthetic rate, but decreased the grain protein content in broomcorn millet. IAA content to a certain concentration could promote the transformation of broomcorn millet from vegetative to reproductive growth.
photoperiod; broomcorn millet; endogenous hormones; plant height; net photosynthetic rate; grain protein content

10.3864/j.issn.0578-1752.2021.02.005
2020-04-20;
2020-06-29
山西省应用基础研究计划(201901D211564)、农业部国家谷子高梁产业技术体系项目(CARS-06-13.5-A16)、国家青年自然科学基金(31601457)、农作物种质资源保护与利用专项(2019NWB036-20)、国家农作物种质资源共享服务平台山西农作物种质资源平台(NICGR-2019-26)
王君杰,E-mail:xiaoleiwangjie@163.com。通信作者乔治军,E-mail:nkypzs@126.com
(责任编辑 杨鑫浩)
