油菜高油酸种质的创建及高油酸性状遗传与生理特性的分析
2021-03-08龙卫华浦惠明高建芹胡茂龙张洁夫陈松
龙卫华,浦惠明,高建芹,胡茂龙,张洁夫,陈松
油菜高油酸种质的创建及高油酸性状遗传与生理特性的分析
龙卫华,浦惠明,高建芹,胡茂龙,张洁夫,陈松
江苏省农业科学院经济作物研究所/农业部长江下游棉花与油菜重点实验室,南京 210014
【】创建高油酸(high oleic,HO)油菜新种质,探明HO新种质中高油酸性状的遗传模式,明确HO新种质油酸含量变化的生理特性,为培育HO油菜新品种奠定基础。采用辐射处理油菜萌动发芽种子,获得初级诱变群体后在后续世代利用极端选择法结合小孢子培养技术筛选油菜HO新种质。以HO新种质分别与3个不同遗传背景的常规油菜品系为亲本组合杂交构建6个世代(P1、P2、F1、BC1P1、BC1P2和F2)的遗传群体,测定各群体脂肪酸含量后应用主基因+多基因混合遗传模型联合分析方法对遗传群体的高油酸性状进行遗传分析。测定HO新种质种子发芽过程中子叶、不同温度处理下苗期营养器官以及角果成熟过程中种子的油酸含量,明确其变化规律及生理效应。通过辐射获得油酸含量显著变化的初级诱变群体,在后续世代持续利用极端选择法得到平均油酸含量升高20个百分点的高世代群体,采用小孢子培养得到纯合稳定的油菜HO双单倍体群体,最终根据品质性状筛选成功得到HO新种质B161,其油脂中油酸含量为85%,亚麻酸含量为3%。以B161为HO亲本和常规品系杂交配置得到3个具有不同遗传背景的遗传群体并测定获得各群体的油酸含量表型数据。脂肪酸含量相关性分析表明,十八碳脂肪酸中油酸含量与亚油酸含量和亚麻酸含量具有显著负相关关系。遗传分析结果表明,该种质中高油酸性状由2对具有加性效应的主效基因控制,并且2对基因对油酸含量的效应值接近。生理分析表明,常温下HO品系营养器官(根、茎、叶和叶柄)的油酸含量均显著高于常规品系,亚麻酸含量显著低于常规品系。低温下HO品系营养器官的油酸含量降低,但仍高于常规品系;常规品系油酸含量在低温下稳定。低温下两类品系营养器官的亚麻酸含量均显著提高,但HO品系亚麻酸含量仍低于常规品系。在种子成熟过程和种子发芽过程中,HOLL品系中油酸含量持续显著高于常规油菜,而亚麻酸含量则持续显著低于常规油菜。成功创制了油菜HO新品系B161,明确了新种质HO性状的遗传规律和生理特性。获得的HO品系具有潜在育种利用价值。
油菜;辐射诱变;种质创新;高油酸性状;遗传分析;低温响应
0 引言
【研究意义】油菜是中国三大主要油料作物之一,菜籽油在中国居民日常生活中具有良好的消费基础[1]。近年来,菜籽油已成为国产植物油的第一大来源,因此,菜籽油的品质(主要是脂肪酸含量比例)对中国居民健康具有重要影响[2]。在中国油菜全面实现“双低化”(2003年)后,十八碳脂肪酸含量的改良成为目前油菜品质育种的重要目标[3]。与普通双低菜籽油相比,高油酸(high oleic,HO)菜籽油[4-5]具有四大优势:(1)较耐贮藏,货架期较长;(2)可降低低密度脂蛋白含量,减少胆固醇形成,预防人体心血管疾病;(3)甲酯化程度高,燃烧值高,更有利于生产生物柴油;(4)有利于提高菜籽油的氧化稳定性,是菜籽油不容易变质生成反式脂肪酸(或位置异构脂肪酸)和产生异味[6]。创建HO甘蓝型油菜新种质并以此培育HO油菜品种是提供高品质菜籽油的有效途径。【前人研究进展】截至目前,国内外学者通过不同方法获得了HO甘蓝型油菜种质。国际上,最早用甲基磺酸乙酯(ethylmethylsulfone,EMS)诱变油菜种子并在后代筛选出高油酸突变系,油酸含量最高可达88%[7-8]。此后,有报道用叠氮化钠诱变出高油酸油菜突变体[9]。国内学者也重视HO材料创制并得到HO种质。和江明等[10]最早用EMS诱变处理油菜小孢子筛选出油酸含量为80.3%的高油酸突变体材料,随后多位学者通过化学诱变得到高油酸突变体[11-13]。而物理方法主要是采用放射性射线辐照产生变异。运用这种方法成功获得高油酸种质的共有2例,一是利用60Co射线辐照油菜干种子后连续定向选择获得最高油酸含量达93.5%(近红外法测定)的株系;二是通过航天诱变方法获得一个高油酸含量(87.22%)的突变株系[13-14]。此外,转基因方法也可以获得高油酸种质。多位学者利用RNAi技术获得了油酸含量超过80%的株系,但转基因种质因需要经过严格的释放前环境评审而不能较快在育种中得到应用,而且目前还未有成功释放的先例[15-20]。综合来看,采用非转基因方法得到的高油酸种质具有可即时应用的比较优势。不同学者对油菜高油酸性状的遗传模式进行了探索。品系19782/7531的高油酸性状(C18:1=78.4%)由2个突变位点控制[21]。对高油酸亲本DMS100(C18:1=77%)与普通品系衍生的DH群体进行QTL定位后发现高油酸性状主要由1个主效位点控制[11]。HO自交系Y539(C18:1=87.22%)的高油酸性状是由分别位于A05和A01染色体上的2个主效基因控制[15]。H005的高油酸性状(C18:1=83.10%)由2对具有加性和显性效应的主效基因控制[22]。HO品系SW Hickory(C18:1=78%)的高油酸性状由位于A5染色体上1个主效QTL控制[23]。HO品系N1379T(C18:1=85%)的高油酸性状则是由位于A5和C5染色体上的2个位点共同控制[24]。【本研究切入点】尽管通过不同手段得到了HO种质,但控制其HO性状的遗传模式并非一致,表明这些种质在遗传上具有不同的机制。同时,已有HO种质的生理特性也未有深入研究。21世纪初江苏省农业科学院经济作物研究所油菜研究室利用辐射方法获得油菜脂肪酸组分的变异,并结合多种育种方法,最终成功创建一个全新的HO种质B161。【拟解决的关键问题】本研究通过对B161中HO性状的遗传模式及其油酸含量变化的生理特性进行研究,推进该种质的育种利用进程,进一步改良菜籽油品质,为提供高品质菜籽油奠定基础。
1 材料与方法
1.1 材料
诱变原始材料为高世代油菜自交系L13-306-171,油酸(C18:1)与亚麻酸(C18:3)含量经近红外分析含量分别约为72.21%和4.18%(后期经气相色谱分析C18:1≈68%,C18:3≈7%),其系谱见傅寿仲等[25]。常规油菜品系N15、N27和N137为3个不同遗传背景的高世代育种自交系。以上材料均由江苏省农业科学院经济作物研究所提供。
1.2 油菜HO种质的诱变及选育过程
选取L13-306-171干净一致的1 000 粒种子(即M0代)排列在培养皿内湿润滤纸上于25℃暗培养,每皿100粒种子,共10皿。待胚根萌动露白后送至扬州市辐照中心进行800 Gray60Co-ɣ射线辐照(辐照剂量参考徐华军等[26])。辐射处理后的萌动种子继续发芽后栽于大田。2004年在辐射当代(即M1代)选择优势单株套袋自交收获种子。测定脂肪酸组分后按油酸含量从高到低选择30株单株种子作为基础选择群体(即M2代)。从每个单株自交种子中分别取等量种子混合种植。随后连续3年重复上年度的方法获得M5代基础群体。在本世代选择质量较好的种子进行半粒法脂肪酸含量测定。2007年秋选取油酸含量>80%的对应半粒种子在室内发芽后栽于大田形成M6代。2008年继续选择油酸高值单株种植于大田,于2009年春季选择单株花蕾进行小孢子培养得到若干DH株系(M7代),选取油酸含量较高且亚麻酸含量较低的DH系作为候选株系。次年经全生育期观察比较后筛选最优HO株系。
1.3 高油酸性状的遗传群体构建
以中选的HO种质B161(P1,C18:1=(85.36±0.50)%)和常规油菜品系N15、N27和N137亲本(P2)种植于江苏省农业科学院溧水植物科学基地油菜育种田,2014年春季杂交分别获得3个杂交组合的共6个正反交F1。2015年将各亲本自交,F1自交得到F2分离群体种子,同时F1与2个亲本分别回交得到BC1代的足量种子(BC1P1和BC1P2)。
1.4 低温与常温下HO品系与常规品系的组织样品制备
将B161和N137的干净种子在培养皿中发芽后至下胚轴长约0.5cm后,移栽至直径为15cm且装满营养土的盆钵中,每品系6钵,每钵4株苗,置于光照培养箱(25℃ 12h光照/15℃12h黑暗,光照强度=2 000 lx)内生长。于三叶期时将对应品系盆钵分成2部分:一半继续放置在原培养箱;另一半置于另一培养箱(12℃ 12h光照/6℃ 12h黑暗,光照强度=2 000 lx)内培养。待7d后将盆钵取出并选取各培养箱内生长一致的菜苗,迅速洗净根系,用剪刀分离根(下胚轴以下部分)、茎(去掉最后一片叶连接点以上部分)、叶(包括去掉主叶脉后的整张叶片)和叶柄等组织,分别将两品系不同单株的相同组织混合置于牛皮纸袋中编号,烘干备用。
1.5 油菜种子成熟及种子发芽过程的样品制备
于大田油菜开花后选择长势均衡、花期一致的B161和N137植株,用棉线标记主轴及倒一、倒二分枝同一天开放的花朵,每个材料标记约10株,每株标记约100朵花,一周后再标记一次作为重复。由于早期的种子含水量很大,故而自角果龄15日开始每7天取样一次,直至种子成熟。大田中角果置于冰盒中带至实验室后,迅速将幼嫩种子从角果中小心剥出,每品系每个样品至少选取100粒种子,及时烘干编号备用。
发芽试验按照粮油检验发芽试验(GB/T 5520-2011)的方法进行,选取B161和N137的饱满种子自开始之日起每天于同一时间剥出发芽种子中的子叶,直至第5天成苗。取样时迅速用镊子剥取子叶部分,每品系每个样品至少选取50个子叶组织,混合后立即烘干编号备用。
1.6 取样及其脂肪酸含量测定
用于遗传分析的6个世代群体种子,按照亲本(P1、P2)、F1、BC1P1、BC1P2和F2分别取出粒数不等的种子,按照高建芹等[27]的半粒法测定种子脂肪酸含量。各个组织样品按照国标(GB/T17377-1998)的方法在安捷伦GC6910型气相色谱仪上测定脂肪酸含量。由于根、茎、叶和叶脉组织的含油量低,因此,其干样至少保证在0.5g以上。在加石油醚-乙醚混合液之前,样品需要充分打样磨碎。
1.7 数据分析、图像绘制以及遗传模型分析
采用Microsoft Excel计算最大值、最小值、平均值、变异系数及相关系数。采用Graphpad Prism7软件进行绘图。根据作物数量性状混合遗传模型及主基因+多基因多世代联合分析方法,采用章元明等[28]发布的植物数量性状软件包(the R software package of SEgregation analysis,SEA)对3个不同遗传背景的6个世代分析群体的高油酸性状进行24个遗传模型的极大似然分析,采用最小AIC值准则筛选最适遗传模型,用最小二乘法估计相应的遗传参数,并估计主基因和多基因效应等遗传参数。
2 结果
2.1 诱变群体各世代油酸与亚麻酸的含量变化及HO种质B161的获得
采用辐射诱变后发现后续世代油酸和亚麻酸含量变异范围明显增加,筛选效果明显。自诱变当代到脂肪酸含量稳定的7个世代,油酸和亚麻酸含量在定向选择过程中均经历了由集中到分散然后又集中的变化趋势(表1)。对2004年(M1代)套袋自交收的单株种子进行脂肪酸分析发现,油酸含量变幅为68.75%—78.81%,亚麻酸含量变幅为3%—8%,表明诱变产生效果。经过4个世代(M2—M5)对油酸进行极端选择后,油酸含量变幅为70.54%—86.95%,但平均值大幅升高。进一步于M6代对极端单株进行小孢子培养后获得具有65个株系的DH系群体,根据脂肪酸测定结果筛选得到油酸含量≥85%且亚麻酸含量为3%—4%的多个株系。次年经全生育期生长势及农艺性状比较后获得HO稳定品系B161(C18:1为85.4%,C18:3为2.5%)。

表1 诱变群体不同世代中C18:1及C18:3含量分布
2.2 HO种质B161高油酸性状的遗传模式
2.2.1 遗传群体中全脂肪酸组分含量的性状变异 成功利用HO种质B161与3个常规品系构建了6个世代分离群体,其中,组合B161×N137的全脂肪酸含量情况如表2。正反交F1的油酸与亚麻酸含量值基本相当,表明HO性状不受细胞质效应的影响。3个组合衍生的6个世代分离群体油酸含量呈连续性分布,符合数量性状的特征。另外,油酸含量在3个群体中呈正态分布,表明该性状在遗传上受多基因控制。
2.2.2 菜籽油中各脂肪酸含量的相关性分析 对组合B161×N137的F2分离群体单株的脂肪酸含量进行了相关性分析(表3),结果表明,在各十八碳脂肪酸中油酸含量除了与亚油酸和亚麻酸含量具有显著负相关以外,还与棕榈酸和二十碳烯酸显著相关,而与硬脂酸含量相关较小。
2.2.3 HO种质B161高油酸含量性状的遗传分析 利用SEA软件对3个遗传分析群体油酸含量数据分别计算所有的遗传模型,获得各个遗传模型的最大似然值以及AIC值。根据最小AIC的遗传模型优选准则,选取各遗传群体中3个最小AIC值的遗传模型作为备选(表4)。结果显示,油酸含量大部分符合2对主效基因的遗传模型,但不同遗传群体估算的基因互作效应略有差别。油酸含量的最佳遗传模型为2MG-A,即2对加性主基因模型。

表2 组合B161×N137的6个世代遗传群体脂肪酸含量统计
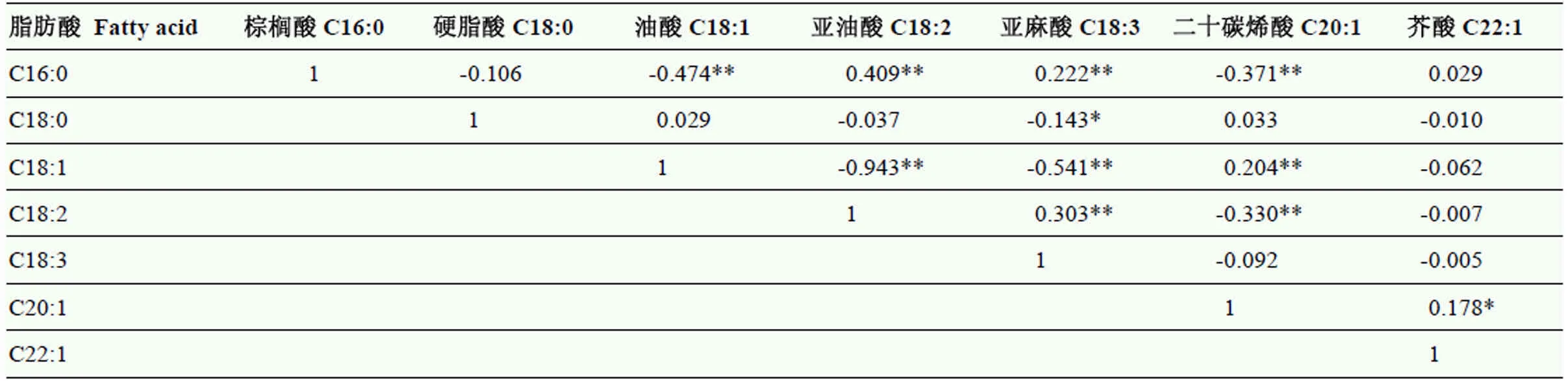
表3 组合B161×N137的F2群体中各脂肪酸含量的相关系数
*表示在0.05水平上差异显著,**表示在0.01水平上差异显著
* means the significance at the 0.05 level,** means the significance at the 0.01 level
对筛选得到的最佳遗传模型估计了遗传参数(表5)。结果显示,控制高油酸含量的2对主效基因在3个组合遗传群体中体现出的效应值均较为接近,每个主效基因可提高油酸含量4—6个百分点。
2.3 HO种质B161与常规品系苗期营养器官中油酸与亚麻酸含量的比较
不同温度下HO品系与常规品系苗期组织中油酸与亚麻酸含量具有显著变化(表6)。常规品系根、茎、叶及叶柄等组织中的油酸含量在常温和低温下没有差异;但HO品系各组织在常温下油酸含量均较常规品系高,低温使各组织油酸含量显著降低,以叶柄降幅最大。无论在低温或常温下常规品系各组织的亚麻酸含量均高于HO品系,低温使HO品系各组织的亚麻酸含量显著升高。
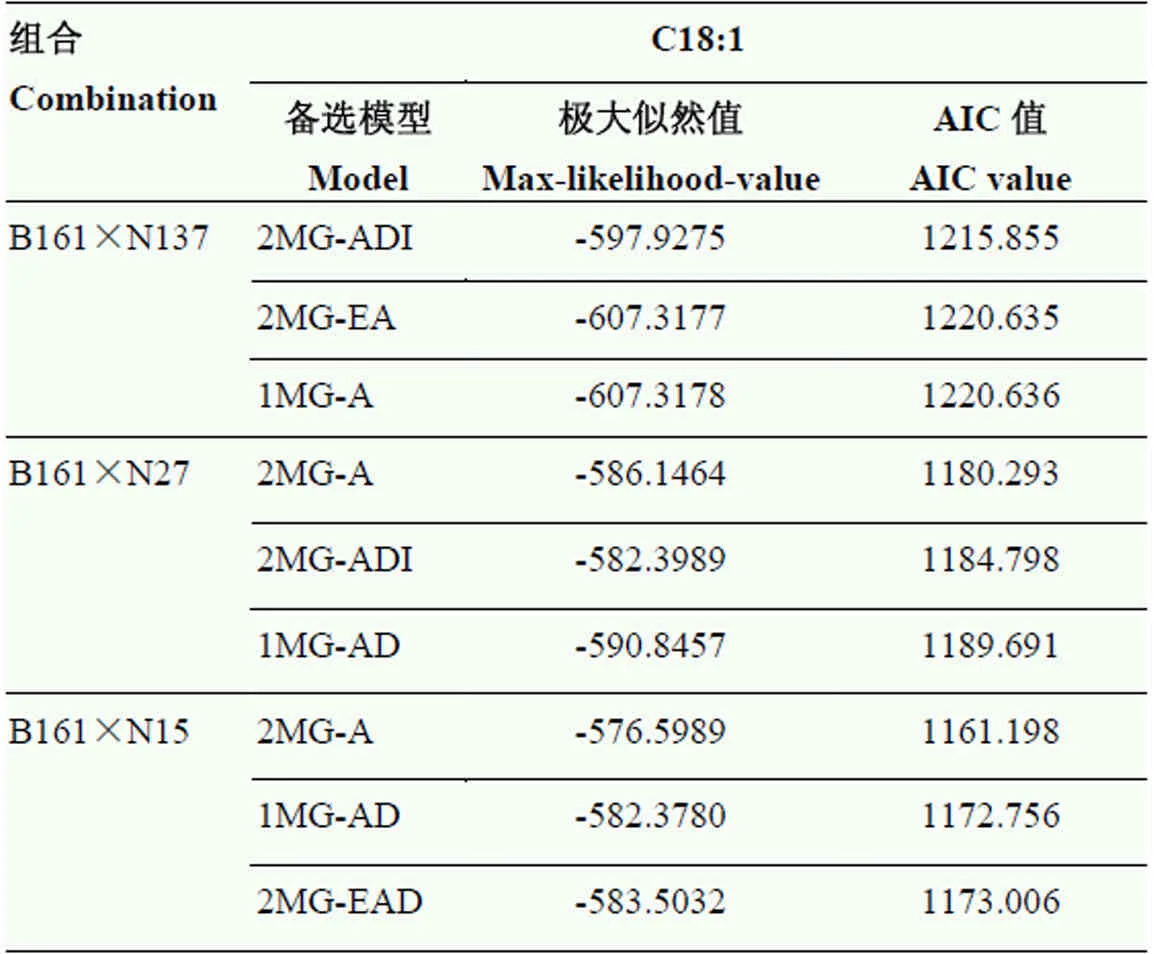
表4 3个组合的候选遗传模型及其极大对数似然函数值和AIC值
MG:主基因;MX:主基因+多基因;A:加性;D:显性;I:互作;E:相等
MG:Major gene; MX:Mixed major gene and polygene; A:Additive; D:Dominance; I:Interaction; E:Equal
2.4 HO种质B161油酸与亚麻酸含量的变化特征
2.4.1 种子成熟过程中HO种质B161与常规品系油酸与亚麻酸的积累HO品系B161与常规油菜品系N137在花后至种子成熟过程中油酸和亚麻酸含量的增减趋势一致,但含量具有显著差异。两品系油酸含量在此段时期内均表现为先迅速增长而后缓慢增长的趋势。B161在花后第15天油酸含量即达到约50%,至第35天达到约80%后仍继续缓慢增长;整个过程高于常规品系20个百分点。两品系亚麻酸含量在此过程中均表现为持续降低趋势,在花后15d时两者差异不大,但在此后差距持续拉大(图1)。

表5 最适遗传模型下3个杂交组合中油酸含量的遗传参数估计
/:无估计值 /:Means no value
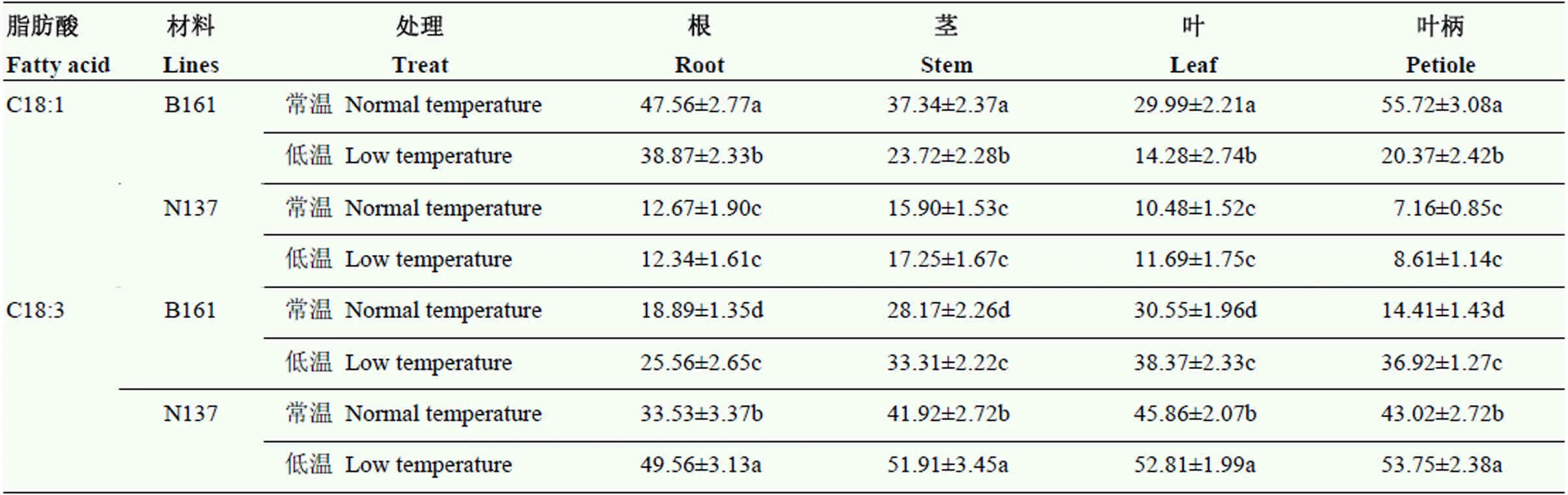
表6 不同温度下HO品系与常规品系苗期不同组织中油酸与亚麻酸的含量
不同小写字母表示在 0.05 水平差异显著 The different lowercase letters in the table indicate significance at 0.05 level

图1 花后HO品系和常规品系种子中油酸与亚麻酸的含量变化
2.4.2 B161与常规品系子叶中油酸与亚麻酸在发芽进程中的代谢HO品系与常规品系子叶中油酸与亚麻酸的相对含量在发芽过程中的变化(图2)具有明显差异。在种子萌发后5d之内,两品系子叶的油酸含量均呈持续下降趋势,下降速率接近,两品系差异值基本维持在20个百分点左右。相反的,在此进程中,两品系亚麻酸含量均呈持续上升趋势,但两品系差异经历了由小到大再变小的过程。
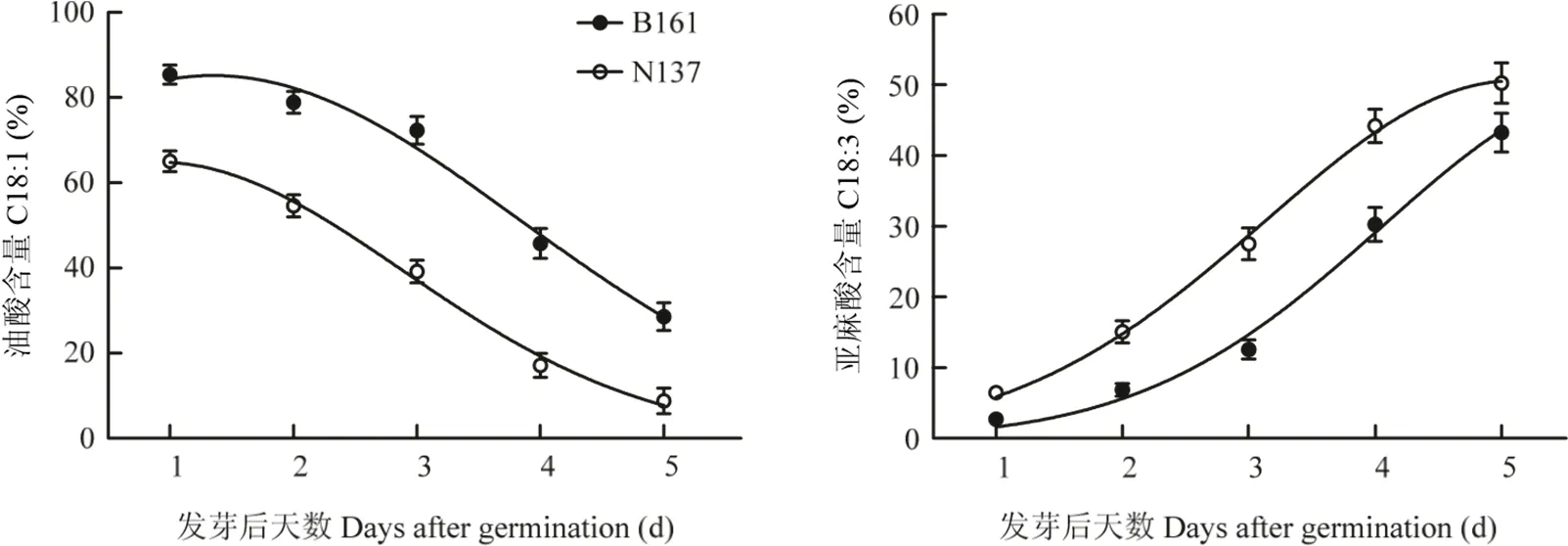
图2 发芽后HO品系和常规品系子叶中油酸与亚麻酸的含量变化
3 讨论
诱变是创建油菜遗传变异材料的有效方法。目前,已报道的非转基因油菜HO种质均通过诱变获得,但诱变方法以及诱变对象不尽相同。大部分HO材料都是通过化学诱变油菜种子获得的[7,9,11-13,20-21,29],也有通过诱变小孢子获得[10],而利用辐射处理油菜种子得到高油酸种质的案例较少[14-15]。本研究亦是采用辐射诱变,但在辐射对象上与前人有所不同。本研究辐射对象是处于萌动露白状态的油菜种子。此种状态下油菜种子中各项生理生化活动刚刚启动,对外界环境极度敏感[30]。从理论上说辐射处理具有较高的诱变率。本研究中在早期世代(M2代)即发现了油酸含量显著升高的株系,不但说明创建思路是可行的,而且也表明突变位点位于控制HO性状的主效基因内,同时这些主效基因可能位于相对容易诱变的染色体区域。此外,由于辐射诱变可能造成油菜染色体变异,若持续采用系谱法选择可能多代自交仍不能纯合稳定,因此,本例中采用小孢子培养技术迅速固定高油酸性状,效果很好,这与LORIN等[9]使用的方法一致。本研究获得的HO品系油酸和亚麻酸含量均与现有报道中的极端值相当,表明所集成的诱变及选择方法在油菜上具有实用性,对其他的类似性状可能也具有借鉴价值。
菜籽油中的油酸含量主要受遗传控制,环境影响较小[21,31]。本研究也发现HO新种质高油酸性状的遗传贡献率高,与前人结果一致。在控制该性状的基因数目上,研究结果有所不同。HU等[11]、官春云等[14]、、YANG等[23]利用QTL定位或基因克隆等方法发现高油酸性状受一个主效基因位点控制。而SCHIERHOLT等[15]和刘列钊等[21]分别利用遗传试验和分子标记筛选表明高油酸性状主要受2个主效位点控制。随后WELLS等[29]利用EMS诱变获得了一批油菜减饱和酶突变体,其不同位点突变对油酸含量的增量效应不同。ZHAO等[32]还发现在A9染色体也有一个位点控制油酸含量。这表明油菜中油酸含量的控制位点较为丰富。本研究中的高油酸新种质虽然也受2对基因控制,但其基因位点需要进一步确认。从育种实践而言,本研究创建的新种质高油酸性状遗传模式相对简单,育种利用较为容易。自2010年开始将HO种质的高油酸性状转育进入萝卜质不育系统并进行了杂交组合配置工作,品比试验发现一些组合的全生育期表现和产量与对照品种相当,表明本研究发现的高油酸种质具有育种利用潜力(电子附表1)。
脂肪酸是植物细胞质膜的主要组成成分,是细胞与环境互作的前哨,因此,脂肪酸的合成与代谢对油菜的非生物胁迫抗性至关重要[33-35]。油菜生育期长,无论是冬油菜还是春油菜在营养生长阶段都会受到温度影响。本研究发现,常温下HO种质营养器官以及种子中的油酸含量均高于常规品系,而亚麻酸含量则低于常规品系,表明(1)HO种质中控制油酸的基因是组成型表达,可能是油菜中2个组成型表达的发生了突变。(2)油菜的脂肪酸合成途径中油酸有2个去向,绝大部分进入减饱和途径生成亚油酸和亚麻酸,少部分进入加碳途径生成二十碳烯酸和芥酸[36]。基于油酸与亚麻酸之间的强烈负相关关系,亚麻酸降低是油酸升高的必然结果。也有可能是HO种质中亚麻酸合成酶也产生突变,导致该酶活性降低,不能合成更多的亚麻酸。本研究还发现,低温下HO种质油酸含量降低和亚麻酸含量增加,但均低于常规品系,这表明(1)低温能够增加突变油酸控制基因的表达,提高油酸的转化率,这与的特性符合[37]。(2)亚麻酸是高不饱和脂肪酸,易于氧化。其含量的增加,可能是油菜进行呼吸作用时更偏好于将亚麻酸氧化从而为其自身提供足够能量;也有可能是因为亚麻酸合成酶活性的变化所致。另外,发现HO种质与常规品系在种子成熟过程和发芽过程中的亚麻酸含量并非持续保持平行差异,说明在HO种质中除了油酸含量变化影响亚麻酸含量之外,亚麻酸本身的合成控制酶也在起作用。值得提出的是,低温下常规品系较HO种质能合成更多的高不饱和度脂肪酸,这意味着常规品系在低温下细胞膜具有更好的流动性,在表型上则可能具有更强的耐寒性。因此可以推断,在较寒冷的油菜产区,HO品系可能耐寒性较常规品系差,需要关注。
4 结论
获得双低甘蓝型油菜HO新种质B161,其高油酸性状受2个主效基因控制。B161中控制高油酸性状的基因为组成型表达,低温对其有诱导增强作用。高油酸种质B161具有育种利用潜力。
[1] 胡忆雨,朱颖璇,杨雨豪,邹军,陈阜,尹小刚.1951-2015年中国主要粮食与油料作物种植结构变化分析.中国农业大学学报,2019,24(11):183-196.
HU Y Y,ZHU Y X,YANG Y H,ZOU J,CHEN F,YIN X G.Changes of the planting structure of major food and oil crops in China from 1951 to 2015.Journal of China Agriculture University,2019,24(11):183-196.(in Chinese)
[2] 刘成,冯中朝,肖唐华,马晓敏,周广生,黄凤洪,李加纳,王汉中.我国油菜产业发展现状、潜力及对策.中国油料作物学报,2019,41(4):485-489.
LIU C,FENG Z C,XIAO T H,MA X M,ZHOU G S,HUANG F H,LI J N,WANGhZ.Development,potential and adaptation of Chinese rapeseed industry.Chinese Journal of Oil Crop Sciences,2019,41(4):485-489.(in Chinese)
[3] 王汉中.以新需求为导向的油菜产业发展战略.中国油料作物学报,2018,40(5):613-617.
WANGhZ.New-demand oriented oilseed rape industry developing strategy.Chinese Journal of Oil Crop Sciences,2018,40(5):613-617.(in Chinese)
[4] SCARTH R,MCVETTY P B E.Designer oil canola:A review of food-gradeoils with focus on high oleic,low linolenic types//WRATTEN N,SALISBURY P A (eds)..Canberra,Australia:GCIRC,1999:26-29.
[5] STANISŁAW S,KATARZYNA M,HANNA C,TERESA P,KRYSTYNA K,MARCIN M,JOANNA N,KRZYSZTOF M,IWONA B.Marker assisted selection of new high oleic and low linolenic winter oilseed rape (L.) inbred lines revealing good agricultural value.PLoS ONE,2020,15(6):e0233959.
[6] REZA F,MOHAMMADhR.Oxidative stability of canola oil bybioactive components during storage at ambient temperature.Food Science and Nutrition,2018,6(2):342-347.
[7] AULD D L,HEIKKINEN M K,ERICKSON D A,SERNYK J L,ROMERO J E.Rapeseed mutants with reduced levels of polyunsaturated fatty acids and increased levels of oleic acid.Crop Sciences,1992,32:657-662.
[8] RUCKER B,ROBBELEN G.Development of high oleic acid rapeseed//Proceeding of 9th International Rapeseed Congress.Cambridge,UK:GCIRC,1995:389-391.
[9] DEBONTE L R,,FAN Z G,MIAO G H.Fatty acid desaturases and mutant sequences thereof:US 6967243 B2 [P].2001-1-29
[10] 和江明,王敬乔,陈薇,李根泽,董云松,寸守铣.用EMS诱变和小孢子培养快速获得甘蓝型油菜高油酸种质材料的研究.西南农业学报,2003,16(2):34-36.
HE J M,WANG J Q,CHEN W,LI G Z,DONG Y S,CUN S X.Studies on rapidly obtaining high oleic acid germplasm ofby mutagen EMS and microspore culture.Southwest China Journal of Agricultural Sciences,2003,16(2):34-36.(in Chinese)
[11] HU X,SULLIVAN GILBERT M,GUPTA M,THOMPSON S A.Mapping of the loci controlling oleic and linolenic acid contents and development ofandallele-specific markers in canola (L.).Theoretical and Applied Genetics,2006,113:497-507.
[12] 张宏军,肖钢,谭太龙,李栒,官春云.EMS 处理甘蓝型油菜()获得高油酸材料.中国农业科学,2008,41(12):4016-4022.
ZHANGhJ,XIAO G,TAN T L,LI X,GUAN C Y.High oleate material of rapeseed () produced by EMS treatment.Scientia Agricultura Sinica,2008,41(12):4016-4022.(in Chinese)
[13] 黄永娟,张凤启,杨甜甜,刘葛山,蒋守华,陈健美,管荣展.EMS 诱变甘蓝型油菜获得高油酸突变体.分子植物育种,2011,9(5):611-616.
HUANG Y J,ZHANG F Q,YANG T T,LIU G S,JIANG S H,CHEN J M,GUAN R Z.High oleate mutants ofproduced by EMS inducement.Molecular Plant Breeding,2011,9(5):611-616.(in Chinese)
[14] 官春云,刘春林,陈社员,彭琦,李栒,官梅.辐射育种获得油菜()高油酸材料.作物学报,2006,32(11):1625-1629.
GUAN C Y,LIU C L,CHEN S Y,PENG Q,LI X,GUAN M.High oleic acid content materials of rapeseed () produced by radiation breeding.Acta Agronomica Sinica,2006,32(11):1625-1629.(in Chinese)
[15] 刘列钊,王欣娜,阎行颖,王瑞,徐新福,卢坤,李加纳.航天诱变高油酸甘蓝型油菜突变体分子标记的筛选.中国农业科学,2012,45(23):4931-4938.
LIU L Z,WANG X N,YAN X Y,WANG R,XU X F,LU K,LI J N.Molecular marker screen for high oleic acid in space flight mutant.Scientia Agricultura Sinica,2012,45(23):4931-4938.(in Chinese)
[16] STOUTJESDIJK P A,HURLESTONE C,SINGH S P,GREEN A G.High-oleic acid Australianandvarieties produced by co-suppression of endogenous D12-desaturases.Biochemical Society Transactions,2000,28:938-940.
[17] 陈苇,李劲峰,董云松,李根泽,寸守铣,王敬乔.甘蓝型油菜基因的 RNA 干扰及无筛选标记高油酸含量转基因油菜新种质的获得.植物生理与分子生物学学报,2006,32(6):665-671.
CHEN W,LI J F,DONG Y S,LI G Z,CUN S X,WANG J Q.Obtaining new germplasm of Brassica napus with high oleic acid content by RNA interference and marker-free transformation ofgene.Journal of Plant Physiology and Molecular Biology,2006,32(6):665-671.(in Chinese)
[18] 田保明,廉玉利,凌华,杨光圣,赵珍,范兴福,李旭娇,师恭曜.RNAi干扰油菜Δ12脂肪酸脱饱和酶基因的表达效果.中国油料作物学报,2009,31(2):132-136.
TIAN B M,LIAN Y L,LING H,YANG G S,FAN S F,LI X J,SHI G Y.Introduction ofgene fragment intovia RNAi plasmid and enhanced oleic acid composition.Chinese Journal of Oil Crop Sciences,2009,31(2):132-136.(in Chinese)
[19] 陈松,浦惠明,张杰夫,高建芹,陈锋,龙卫华,胡茂龙,戚存扣.转基因高油酸甘蓝型油菜新种质的获得.江苏农业学报,2009,25(6):1234-1237.
CHEN S,PUhM,ZHANG J F,GAO J Q,CHEN F,LONG W H,HU M L,QI C K.Identification of high oleic acid germplasm from the T2progency of the transgenicL..Jiangsu Journal of Agriculture Sciences,2009,25(6):1234-1237.(in Chinese)
[20] PENG Q,HU Y,WEI R,ZHANG Y,GUAN C,RUAN Y.Simultaneous silencing ofandgenes affects both oleic acid and erucic acid contents inseeds.Plant Cell Report,2010,29:317-325.
[21] SCHIERHOLT A,BECKERhC.Environmental variability and heritability of high oleic acid content in winter oilseed rape.Plant Breeding,2001,120:63-66.
[22] 费维新,吴新杰,李强生,陈凤祥,侯树敏,范志雄,江莹芬,雷伟侠,荣松柏,段晓莉,胡宝成.甘蓝型油菜高油酸材料的遗传分析.中国农学通报,2012,28(1):176-180.
FEI W X,WU X J,LI Q S,CHEN F X,HOU S M,FAN Z X,JIANG Y F,LEI W X,RONG S B,DUAN X L.Genetic analysis of high oleic acid mutation materials in.Chinese Agricultural Science Bulletin,2012,28(01):176-180.(in Chinese)
[23] YANG Q,FAN C,GUO Z,QIN J,WU J,LI Q.Identification ofandgenes ingenome and development of allele-specific markers for high oleic and low linolenic acid contents.Theoretical and Applied Genetics,2012,125:715-729.
[24] LONG W,HU M,GAO J,CHEN S,ZHANG J,CHENG L,PU H.Identification and functional analysis of two new mutantAlleles that confer elevated oleic acid content in rapeseed.Frontiers in Genetics,2018,9:399.
[25] 傅寿仲,吕忠进,戚存扣,陈爱华.甘蓝型油菜十八碳烯酸的进一步改良.江苏农业学报,1995,11(1):16-20.
FOU S Z,LÜ Z J,QI C K,CHEN A H.Further modification of levels of C18 unsaturated fatty acids in rapeseed.Jiangsu Journal of Agriculture Sciences,1995,11(1):16-20.(in Chinese)
[26] 徐华军,贺源辉,陈秀芳.60Co 射线对双低甘蓝型油菜(L.)的辐射效应.核农学报,1992,6(4):199-206.
XUhJ,HE Y H,CHEN X F.The effects of gamma irradiation on morphological cytological and biochemical characters of double-low rape (L.).Acta Agriculturae Nucleatae Sinica,1992,6(4):199-206.(in Chinese)
[27] 高建芹,浦惠明,戚存扣,张洁夫,陈新军,龙卫华,傅寿仲.应用气相色谱仪分析油菜脂肪酸含量.江苏农业学报,2008(5):581-585.
GAO J Q,PUhM,QI C K,ZHANG J F,CHEN X J,LONG W H,FU S Z.Analysis of fatty acid content in rapeseed by gas chromatography.Jiangsu Journal of Agriculture Sciences,2008(5):581-585.(in Chinese)
[28] ZHANG Y M.Segregation Analysis of Quantitative Traits and Its R Software.Beau Bassin,Mauritius:Golden Light Academic Press,2017.
[29] WELLS R,TRICK M,SOUMPOUROU E,CLISSOLD L,MORGAN C,WERNER P.The control of seed oil polyunsaturated content in the polypoid crop species.Molecular Breeding,2014,33:349-362.
[30] 曾新华.不同诱变方法对油菜种子诱变效果及突变体的研究[D].武汉:华中农业大学,2010.
ZENG X H.Comparing effectiveness of different mutagens for seed quality and analysis of mutants in[D].Wuhan:Huazhong Agriculture University,2010.(in Chinese)
[31] ZHANGhZ,SHI C H,WU J G,REN Y L,LI C T,ZHANG D Q,ZHANG Y F.Analysis of genetic and genotype × environment interaction effects from embryo,cytoplasm and maternal plant for oleic acid content ofL..Plant Science,2004,167(1):43-48.
[32] ZHAO Q,WU J,CAI G,YANG Q,SHAHID M,FAN C,ZHANG C,ZHOU Y.A novel quantitative trait locus on chromosome A9 controlling oleic acid content in.Plant Biotechnology Journal,2019,17(12):2313-2324.
[33] LI Q,ZHENG Q,SHEN W Y,CRAM D,FOWLER D B,WEI Y D,ZOU J T.Understanding the biochemical basis of temperature-induced lipid pathway adjustments in plants.The Plant Cell,2015(27):86-103.
[34] HOU Q C,UFER G,BARTELS D.Lipid signaling in plant responses to abiotic stress.Plant,Cell and Environment,2016,39:1029-1048.
[35] STACY D S,ZOU J T,RANDALL J W.Abiotic factors influence plant storage lipid accumulation and composition.Plant Science,2016,243:1-9.
[36] KONG Q,YANG Y Z,GUO L,YUAN L,MA W.Molecular basis of plant oil biosynthesis:insights gained from studying the WRINKLED1 transcription factor.Frontiers in Plant Sciences,2020,11:24.
[37] DAR A A,CHOUDHURY A R,KANCHARLA P K,ARUMUGAM N.Thegene in plants:Occurrence,regulation and role.Frontiers in Plant Sciences,2017,8:1789.
Creation of High-Oleic (HO) Canola Germplasm and the Genetic and Physiological Analysis onHOTrait
LONG WeiHua,PU HuiMing,GAO JianQin,HU MaoLong,ZHANG JieFu,CHEN Song
Institute of the Industrial Crops,Jiangsu Academy of Agriculture Sciences/Key Lab of Cotton and Rapeseed (Nanjing) of Ministry of Agriculture,Nanjing 210014
【】 This study is to create the new high oleic (HO) canola germplasm,to explore its genetic mode and the physiological characters of theHOtrait,which will lay a foundation for breedingHOcanola varieties.【】 The primary mutation population with was obtained by radiation treatment of germinating canola seeds.The newHOgermplasm was screened by extreme selection method combined with microspore culture technology in subsequent generations.The genetic populations of six generations (P1,P2,F1,BC1P1,BC1P2and F2) were constructed by crossing theHOGermplasm with three conventional canola lines with different genetic background.After the fatty acid content of each population was determined,the genetic analysis of high oleic acid content in the genetic population was analyzed by the mixed major-gene plus polygene inheritance model.The oleic acid content in cotyledons during seed germination,vegetative organs at seedling stage in different temperature regimes and seeds during silique ripening process of theHOgermplasm were detected to explore their change patterns and physiological effects.【】 The primary mutation population with significantly increased oleic acid content was obtained by radiation treatment,and then the high generation population with 20-percent-increased oleic acid content was obtained by using extreme selection method in subsequent generations and the double haploid population was obtained by microspore culture.Finally,a newHOgermplasm B161 (C18:1=85%,C18:3=3%) was successfully screened according to the quality traits.Three genetic populations with different genetic background were obtained by crossing B161 asHOparent with three other conventional lines.The correlation analysis of fatty acid contents showed that there was a significant negative correlation between oleic acid content,linoleic acid content and linolenic acid content.The results of genetic analysis showed that the high oleic acid content was controlled by two major genes with additive effect,and their effect values were close.Physiological analysis showed that the contents of oleic acid in vegetative organs (root,stem,leaf and petiole) ofHOline were significantly higher than those of the conventional strain,and the linolenic acid contents ofHOline were significantly lower than those of the conventional line.The contents of oleic acid in vegetative organs ofHOline decreased at low temperature,but they were still higher than those of the conventional line.The linolenic acid contents in vegetative organs of the two lines increased significantly at low temperature,but the linolenic acid content ofHOline was still lower than that of the conventional line.During the process of seed ripening and seed germination,the oleic acid content ofHOline was significantly higher than that of conventional line,while the linolenic acid content was significantly lower than that of conventional line.【】The newHOgermplasm was successfully created and the genetic mode and physiological characters were confirmed.ThisHOgermplasm has the potential value in breeding.
L.; radiation mutagenesis; germplasm creation; high oleic trait; genetic analysis; response to low temperature
附表1 高油酸低亚麻酸杂交组合产量和菜籽品质

Supplemental table 1 The yields and the seed-oil quality of the HOLL hybrid combinations
*表示在0.05水平上差异显著 * Significance at the 0.05 level

10.3864/j.issn.0578-1752.2021.02.003
2020-07-04;
2020-09-01
国家重点研发项目(2016YFD0101300)、江苏省现代作物生产协同创新中心(JCIC-MCP)
龙卫华,Tel:025-84390368;E-mail:long-weihua@163.com。通信作者浦惠明,Tel:025-84390370;E-mail:puhuiming@126.com
(责任编辑 李莉)
