高黎贡山南段长蕊木兰天然更新研究
2021-03-04李贵祥和丽萍赵欣凤马赛宇陈建洪
李贵祥,柴 勇,和丽萍,赵欣凤,毕 波,马赛宇,陈建洪
(云南省林业和草原科学院,云南 昆明 650201)
长蕊木兰Alcimandra cathcartii为木兰科Magnoliaceae长蕊木兰属Alcimandra常绿乔木,别名黄心树、黄泡心、团叶黄心,树高可达50 m[1],胸径可达80 cm[2],是稀有的单种属植物,《国家重点保护野生植物名录(第一批)》将其列为Ⅰ级保护植物,世界自然保护联盟(International Union for Conservation of Nature, IUCN)《濒危物种红色名录》将其列为濒危(EN)物种。长蕊木兰主要分布于中国云南西南部至东南部、西藏南部和东南部,印度东北部、锡金、不丹、缅甸北部,越南北部也有分布[3]。长蕊木兰喜温暖湿润的环境,多生长在砂页岩、石灰岩等发育的酸性黄壤、黄棕壤,土层深厚,有机质含量较高的土壤中,常成片或散生于海拔1 100~2 800 m的山地季风常绿阔叶林、半湿润常绿阔叶林或中山湿性常绿阔叶林中。长蕊木兰木材纹理通直,有光泽,是室内装修、建筑、家具等优良用材;其树形美观,是较好的园林景观绿化树种[2]。森林更新是一个极为重要的生态学过程[4−5],是森林得以繁衍,持续不衰的基础[6]。森林更新包括人工更新和天然更新,对天然更新加以人工辅助,则为人工促进天然更新。森林天然更新主要受到环境因子与林分因子等的影响[7],不同森林类型,其树种特性、更新过程和演替方向会有所差异。长蕊木兰的天然更新将对该种群的繁衍起着极其重要的作用。目前,对长蕊木兰的研究主要集中在形态特征、人工繁育、光合特性、分布格局、遗传多样性等方面[8−13],对其天然更新研究未见报道。本研究调查和分析了长蕊木兰种群的天然更新情况,旨在为长蕊木兰的天然更新和人工促进更新提供理论基础和科学依据,从而为长蕊木兰种群扩繁提供支撑。
1 研究区概况
研究区位于高黎贡山南段,地处云南省保山市隆阳区的赧亢、腾冲市的小地方和龙陵县的小黑山交汇地段。该区域为长蕊木兰较为集中分布的地带,面积约4 km2,为高黎贡山自然保护区隆阳管护分局赧亢管护站、腾冲市管护分局整顶管护站和小黑山省级自然保护区管辖区域。地势东高西低,地处中国西南部亚热带高原季风气候,气候主要受西南季风和西风南支急流2支风向基本相同但物理性质不同的气流交替控制。全年盛行西南风,四季不分明,干湿季显著,气温日较差大,年较差小,兼有大陆性和海洋性气候特征。年平均气温12.0~17.0 ℃,最冷月为1月,气温为7.0 ℃左右,年降水量1 200.0 mm左右。11月下旬至翌年4月为干季,日照充足,降水少,相对湿度50%~60%。5−10月为湿季,降水量占全年的87%,相对湿度比干季高20%左右。研究区土壤为黄棕壤、棕壤,主要植被类型为中山湿性常绿阔叶林。
2 研究方法
2.1 野外调查
2.1.1 调查母树选择 在长蕊木兰分布的区域,通过踏查,选择具有天然下种能力的长蕊木兰单株作为母树,在所选母树半径为30 m范围内无其他母树,以保证在调查样圆内母树天然更新的唯一性。共选择8株母树的所有更新植株进行统计分析,分别记录母树的位置、海拔、郁闭度、胸径、树高和冠幅等。所选择母树基本情况见表1。
2.1.2 调查方法 以母树为中心向外辐射,按 0~5、5~10、10~15、15~20、20~25、25~30 m 的环形样带进行调查,并在相应的距离范围内,按方位角 0°~90°、90°~180°、180°~270°、270°~360°分别调查长蕊木兰更新的株数。调查时,记录更新植株与母树的相对位置、树高、地径、冠幅等指标。
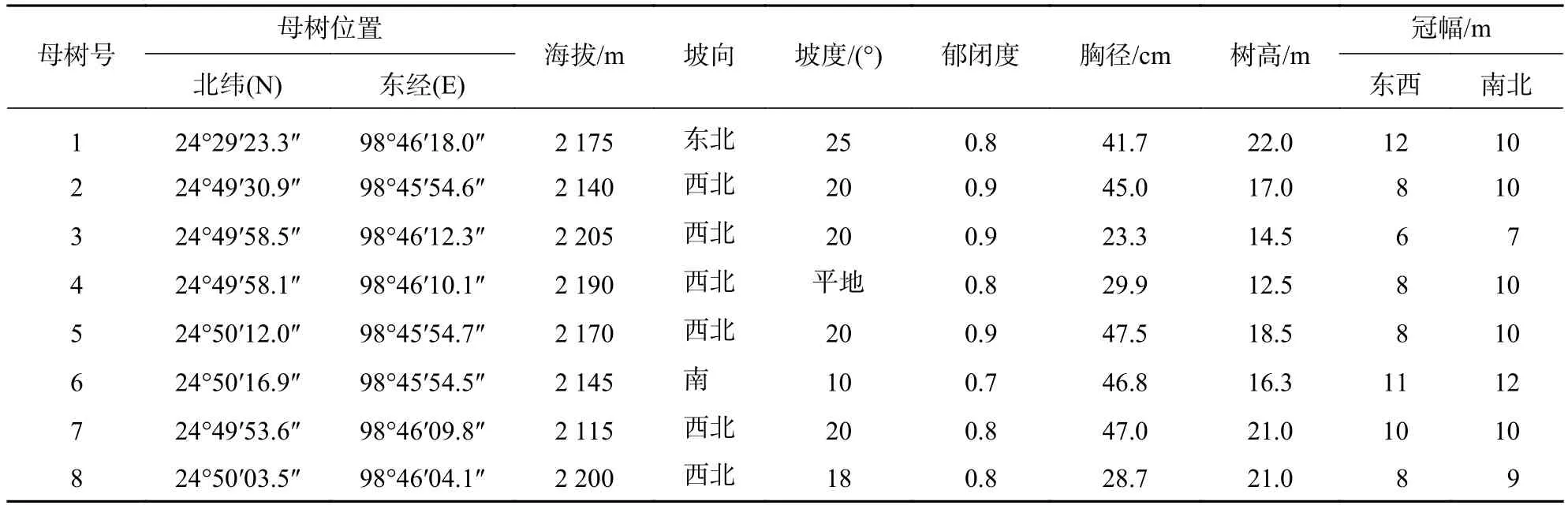
表1 长蕊木兰天然更新调查母树基本情况Table 1 Investigation of seed trees of natural regeneration of A. cathcartii
2.2 数据处理
2.2.1 种群分布格局 密度计算公式:D=x/s。其中:x为样方中某个种的个体数;s为样地面积;D为密度。种群分布格局由下列公式计算[14]:其中:xi为样方中某个种的个体数;m为各样方中某个种个体平均数;n为取样个体数;S2为分散度(方差)。
2.2.2 年龄结构划分 采用径级大小结构代替年龄结构的方法来分析其结构特征[15]。以长蕊木兰的树高(H)对天然更新苗木进行分级:H<1 m划为幼苗幼树,1 m≤H<5 m划为小树,H≥5 m划为中树。母树作为大树不计入更新分析。
2.2.3 数据整理分析 采用Excel 2003对长蕊木兰更新苗木进行统计整理,并在不同方位角和不同距离上进行方差分析。
3 结果与分析
3.1 长蕊木兰空间格局
种群分布格局是指种群中的个体在其水平空间的分布状况,反映了种群内的个体间在水平空间上的相互关系[16]。不同植物群落中的每个种群都有自己的分布格局,其表现的方式主要有:聚集分布、随机分布和均匀分布。通过对植物群落中种群分布格局的测定,可以揭示该群落空间结构特征。
从表2可见:8个样圆中,共有长蕊木兰75株。长蕊木兰种群密度为10~81株·hm−2,平均密度为32株·hm−2。长蕊木兰种群内分布型的指标方差和平均数的比率(S2/m)为4.14,显著大于1.00,说明长蕊木兰在调查样圆内为集群型分布,其种群个体分布不均匀,幼树的分布是集群型的。
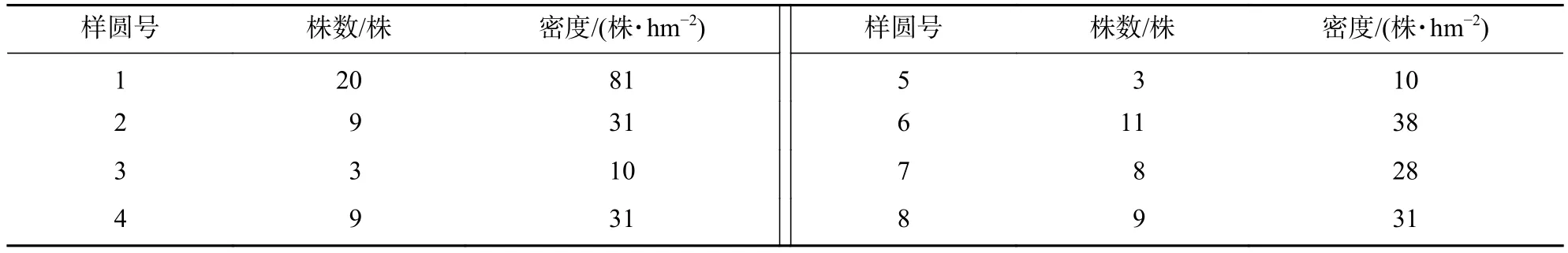
表2 长蕊木兰在不同样圆中的更新数量Table 2 Number of regeneration of A. cathcartii in different sample circle
3.2 长蕊木兰更新分析
3.2.1 不同龄级上的植株状况 长蕊木兰的天然更新能力是该种群得以繁衍的基础。从长蕊木兰天然更新调查结果来看(表3):在8个样圆分布的75株长蕊木兰植株总数中,母树8株,更新苗67株,树高H<1 m的幼苗幼树31株,占总株数的46.27%;1 m≤H<5 m的小树22株,占总株数的32.83%;H≥5 m的中树14株,占总株数的20.90%。从幼苗幼树、小树、中树更新数量来看,苗木数量依次递减,呈金字塔分布结构,说明长蕊木兰更新数量虽然较少,但其结构相对较为稳定。
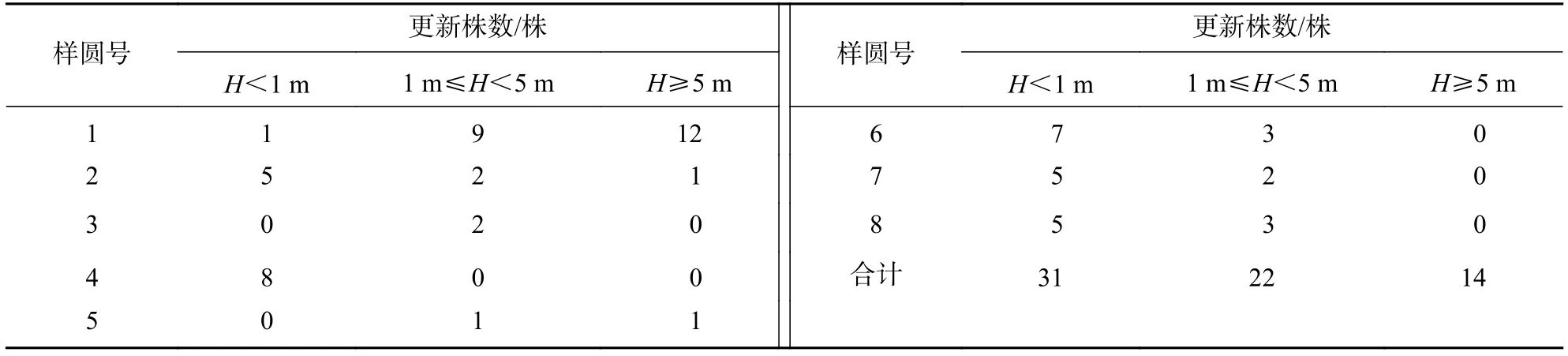
表3 长蕊木兰在不同龄级的更新状况Table 3 Natural regeneration of A. cathcartii in different ages
3.2.2 不同方位角上的更新状况 从长蕊木兰不同方位角更新株数来看(表4):在方位角270°~360°的更新株数最多,占32.84%;0~90°的更新株数18株,占26.86%;在90°~180°和180°~270°更新株数分别为13和14株,所占比例分别为19.40%和20.90%。从不同方位角的更新状况来看:长蕊木兰更新有一定差异,但从其更新分布的株数方差分析来看(表5):检验统计量F=0.383 319,显著性水平P=0.765 836>0.05,不同方位角长蕊木兰更新分布株数差异不显著。
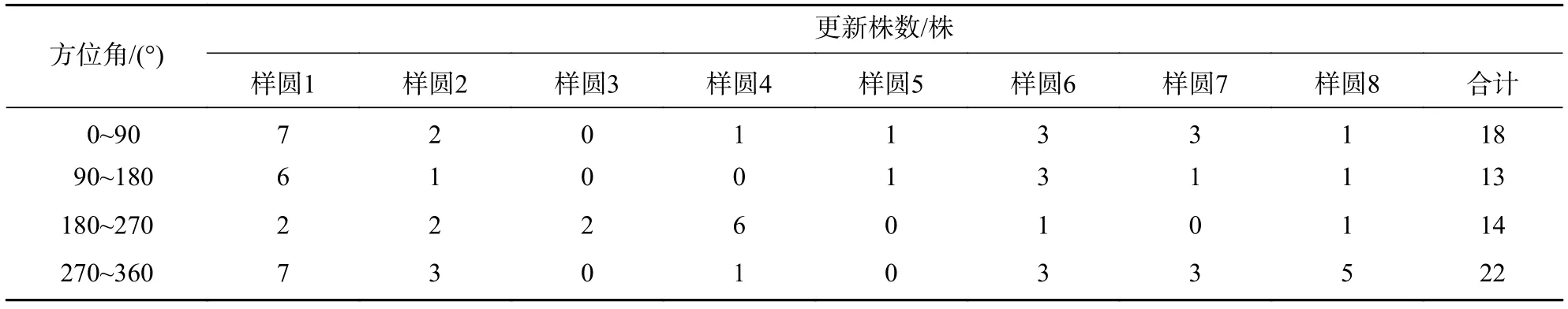
表4 长蕊木兰不同方位角上的天然更新状况Table 4 Natural regeneration of A. cathcartii in different azimuths

表5 长蕊木兰在不同方位角天然更新数量的方差分析Table 5 Analysis of variance of natural regeneration numbers of A. cathcartii in different azimuths
3.2.3 不同距离的更新状况 长蕊木兰的更新与母树距离具有一定的关系。从母树不同距离的长蕊木兰更新情况看 (表6):在 0~5、5~10、10~15、15~20、20~25、25~30 m 距离范围内所占的株数比例分别为11.94%、34.33%、23.88%、22.39%、5.97%、1.49%。表明离母树的距离不同,长蕊木兰更新株数不一样,更新株数最多是 5~10 m,其余依次为 10~15、15~20、0~5、20~25 和 25~30 m,5~20 m 距离更新株数所占比例为总更新株数的80.60%。从离母树不同距离的长蕊木兰更新株数方差分析来看(表7):检验统计量F= 3.376 084,P<0.05,离母树不同距离的长蕊木兰更新株数差异显著。
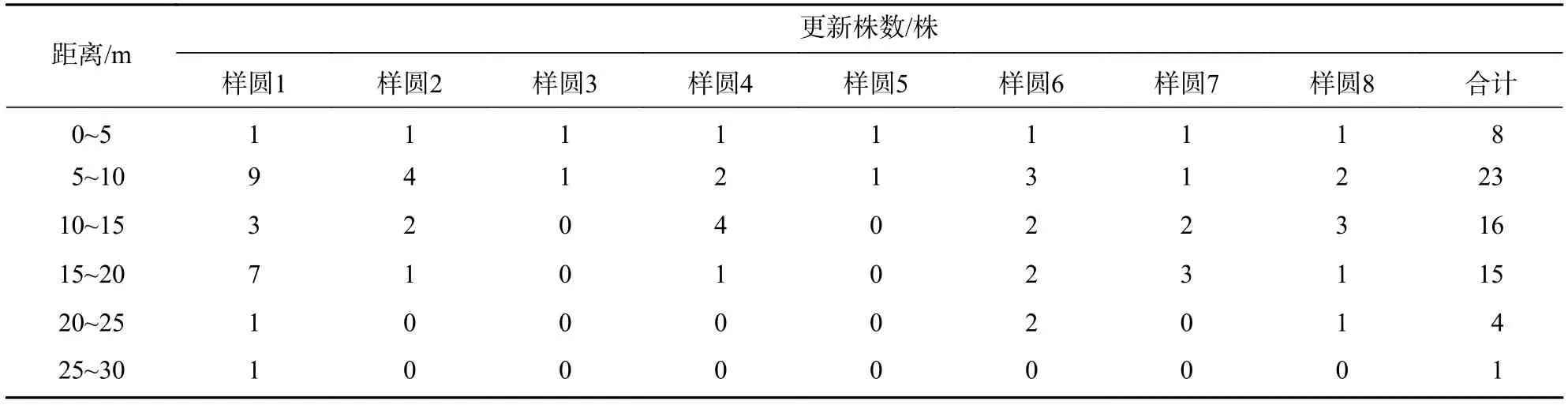
表6 长蕊木兰不同距离范围内的天然更新状况Table 6 Natural regeneration of A. cathcartii in different distances

表7 长蕊木兰不同距离范围内的天然更新方差分析Table 7 Analysis of variance of natural regeneration numbers of A. cathcartii in different distances
4 结论与讨论
植物种群分布格局一方面与种群本身的特性有关,另一方面还受生境条件以及种群间相关性的影响[16]。从高黎贡山南段长蕊木兰分布格局来看,其种群内分布型的指标方差和平均数的比率(S2/m)为4.14,显著大于1,表明长蕊木兰在调查样圆内为集群型分布,种群个体分布不均匀,幼树的分布是集群型的。聚集分布格局会满足幼苗的生长需要,可以群聚的形式来增强对其他植物种竞争的能力[17]。
长蕊木兰的更新与母树的结实量、种子的成熟情况以及气候等环境因子有关。长蕊木兰在8个样圆内的更新数量为67株,更新数量较少,这可能与长蕊木兰种子多不饱满,发芽率为50%~60%有一定关系[2]。另外,调查中发现,长蕊木兰种子是长吻松鼠Dremomys pernyi及其他动物较为喜好的食物,在一定程度上影响了长蕊木兰的更新。从所更新的幼苗径级分布来看,幼苗幼树、小树和中树更新比例分别为46.27%、32.83%和20.90%,其更新数量呈金字塔结构,说明长蕊木兰更新数量虽少,但其结构相对较为稳定。
不同方位角更新数量可能和坡度、不同范围分布的枝条有关。从长蕊木兰不同方位角更新株数来看,在 0°~90°、90°~180°、180°~270°和 270°~60°更新株数比例分别为 26.86%、19.40%、20.90% 和32.84%。各个方位角的长蕊木兰更新株数有一定差异,但从不同方位角的分布株数的方差分析来看,长蕊木兰更新分布株数差异极不显著。
长蕊木兰的更新与母树距离具有一定的关系。在 0~5、5~10、10~15、15~20、20~25、25~30 m距离范围内更新株数所占的比例分别为11.94%、34.33%、23.88%、22.39%、5.97%、1.49%。长蕊木兰更新株数最多的是距离母树 5~10 m 范围,其余依次为 10~15、15~20、0~5、20~25 和 25~30 m。距离长蕊木兰母树5~20 m范围更新株数所占比例为80.60%,说明长蕊木兰更新主要分布在距离母树5~20 m。方差分析表明:离母树不同距离的长蕊木兰更新株数存在显著差异。对8个样圆中距离母树30~35 m的范围进行了更新调查,均没有长蕊木兰更新,说明长蕊木兰更新较窄,难以扩大区域,种群的扩大需要人工促进更新或人工更新。
