不同种源山桐子光合特性分析
2021-03-04徐自恒房丽莎刘震李志蔡齐飞耿晓东翟雯婧王炳焱詹志伟王艳梅
徐自恒, 房丽莎, 刘震, 李志, 蔡齐飞, 耿晓东, 翟雯婧, 王炳焱, 詹志伟, 王艳梅
(1.河南农业大学林学院,河南 郑州 450002;2.河南省漯河市森林病虫害防治检疫站,河南 漯河 462000;3.河南省国有鲁山林场,河南 平顶山 467300)
山桐子(Idesiapolycarpa)是大风子科(Flacourtiaceae)山桐子属的高大落叶乔木,生长迅速,树形美观,红色果实艳丽夺目,可作为城市绿化观赏树种[1]。山桐子果实中不饱和脂肪酸含量较高,其中亚油酸含量高达70%[2],是优良的木本油料树种,有“美丽的树上油库”之美称[3]。山桐子经济效益高,适应性强,分布广泛,侧根发达,耐贫瘠,是中国荒山造林的优良树种[4]。随着中国木本油料行业的快速发展,山桐子的栽培面积也在迅速增加,优良种源选择成为当前产业发展需要解决的关键问题。已有研究表明,同一物种不同种源间在同一地区光合性能表现差异明显[5-7],且光合性能优异的种源表现更优[8]。不同种源只有经过地区生长适应性评价才可用于推广应用,而光合性能是评价植物对环境适应性的重要指标[9],树木干物质积累的90%以上来源于光合作用[10],光合产物为林木生长与果实发育提供物质基础,使用高光效种源来提高林木产量和质量越来越受到重视[11-13]。黄跃宁等[14]和王小菲等[15]对不同生境的栓皮栎幼苗在同一地区栽植的光合特性研究中发现,南方种源比北方种源的光合潜能高,刘炳妤等[16]对不同种源的柚木无性系研究发现,不同种源间光合特性差异极显著,白雪卡等[17]认为,种源地气候会影响不同种源蒙古莸光响应特性,其光合能力的差异是适应环境的体现。目前山桐子研究主要在种子萌发[18-19]、品种栽培[20]、生理生态[21-22]和油脂加工利用[23]等方面,但关于山桐子光合特性研究较少。本试验对4个种源山桐子不同月份的光合生理参数进行测定,旨在研究山桐子在种植地域的光合特性,为后期山桐子的种源选择及高效栽培提供理论依据。
1 材料与方法
1.1 试验地概况
试验地选在河南省郑州市金水区河南农业大学林学试验站,地理位置113°38′E、34°47′N,海拔为108 m,四季分明,该地区最高气温为43 ℃,最低气温为-17.9 ℃,年平均气温为14.2 ℃;热量充足,年降雨量为650.1 mm,无霜期为215 d,全年光照时间约为2 400 h,属于典型的半干旱半湿润大陆性季风气候。土壤为潮土,pH值约为7.0。
1.2 试验材料
选择张家界、成都、广元和东京4个地区引种的种子播种在郑州地区并生长4 a的山桐子为试验材料,株行距为4 m×4 m,采取山桐子常规栽培技术管理。
1.3 试验方法
1.3.1 不同种源山桐子叶片在不同月份光合参数测定 每个种源选取3株生长较好无病虫害且树体大小一致的山桐子,每株选择冠层中部向阳叶片作为测试叶,使用LCpro-SD便携式光合仪,5—10月,每月选择1 d晴朗、无风的天气,从8:00—18:00每隔2 h测定1次,主要测定指标为净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2摩尔分数(Ci)。
1.3.2 不同种源山桐子叶片光响应曲线和CO2响应曲线测定 采用LCpro-SD光合仪,利用自身配备的红蓝光源将光合有效辐射(PAR)设置为:0、50、100、200、400、600、800、1 000、1 200、1 500 μmol·m-2·s-1,叶室CO2摩尔分数控制为600 μmol·mol-1,测定4个种源不同光合有效辐射所对应的Pn值。
CO2响应曲线测定:气源采用CO2小钢瓶,CO2摩尔分数梯度为:0、8、150、200、400、600、800、1 000、1 200、1 400,PAR设定为1 000 μmol·m-2·s-1,测定4个种源不同CO2摩尔分数所对应的Pn值。
1.4 数据处理与分析
采用叶子飘模型[24]对光响应曲线和CO2响应曲线进行拟合,作图软件采用Origin 8.0。
2 结果与分析
2.1 不同种源山桐子净光合速率分析
不同种源的山桐子Pn在不同月份的日变化规律如图1所示,4个种源山桐子不同月份日变化趋势一致,均为双峰曲线,广元、东京、成都种源不同月份Pn差异不明显,张家界种源不同月份Pn差异较大,4个种源的净光合速率在上午10:00达到峰值后均逐渐下降,而后在16:00时又出现了一个高峰(东京种源7、8月份峰值出现在14:00),4个种源均有明显的“午休”现象。整体看来,4个种源5—8月份的山桐子Pn明显大于9、10月份,不同之处在于,张家界种源在6月的Pn值最高,为12.66 μmol·m-2·s-1,广元种源5月的Pn值最高,为9.41 μmol·m-2·s-1,东京和成都种源7月的Pn值最高,分别为9.36 和12.65 μmol·m-2·s-1,4个种源Pn均在10月最低。

图1 不同种源不同时期山桐子净光合速率日变化Fig.1 Diurnal variation of net photosynthetic rate of Idesia policarpa in different provenances and periods
2.2 不同种源山桐子气孔导度分析
不同种源山桐子气孔导度在不同月份的日变化规律如图2所示,不同种源山桐子每日测定的叶片Gs随时间变化呈双峰曲线,4个种源山桐子不同月份Gs差异较小,均在10:00出现最大值,下午16:00出现第2个峰值。除张家界种源外,其余3个种源均是上午Gs峰值大于下午。张家界、成都种源在6月达到峰值,分别为0.26和0.27 μmol·m-2·s-1,广元种源在5月达到峰值,为0.26 μmol·m-2·s-1,东京种源在7月达到峰值,为0.35 μmol·m-2·s-1。4个种源Gs在10月均最小。
2.3 不同种源山桐子胞间CO2摩尔分数分析
不同种源山桐子胞间CO2摩尔分数在不同月份的日变化规律如图3所示,4个种源不同月份Ci变化趋势基本一致,但不同月份之间差异均较大,4个种源不同月份Ci整体呈先下降,在10:00达到最低值后逐渐趋于平稳的趋势。张家界、成都、广元种源在5月Ci整体呈现最高趋势,东京种源在6月呈现最高趋势。成都、广元、东京3个种源在8月Ci均呈现最低趋势,张家界种源Ci在6月整体呈现最低趋势。
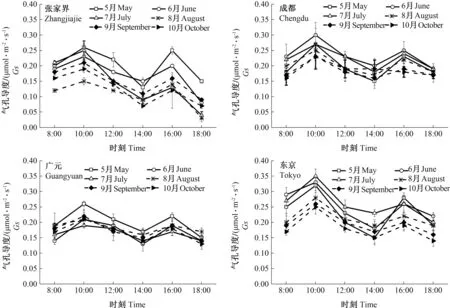
图2 不同种源山桐子气孔导度变化

图3 不同种源山桐子胞间CO2摩尔分数变化
2.4 不同种源山桐子蒸腾速率分析
不同种源山桐子蒸腾速率在不同月份的日变化规律如图4所示,不同种源山桐子Tr的总体变化趋势呈先上升后下降的倒“V”形,4个种源不同月份Tr均在12:00达到最大值,在12:00—14:00的Tr也明显较高,除张家界种源在6月Tr最高外(7.38 μmol·m-2·s-1),成都、广元、东京3个种源均在7月份Tr最高,分别为7.23、7.75和7.88 μmol·m-2·s-1。4个种源Tr的最小值均出现在10月。
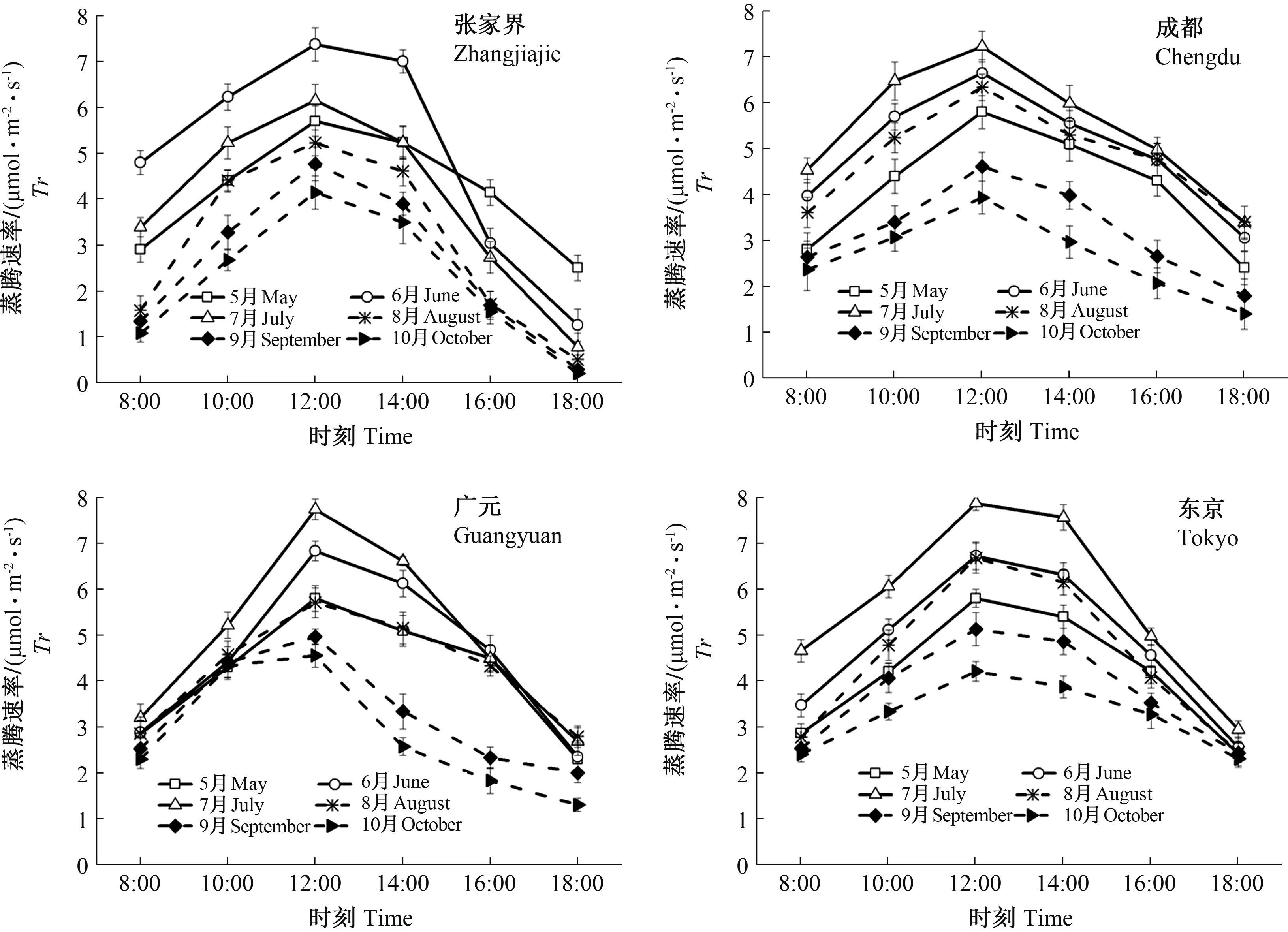
图4 不同种源山桐子叶片蒸腾速率
2.5 不同种源山桐子光响应曲线分析
4个种源山桐子的光响应曲线变化趋势基本一致(图5),在PAR≤400 μmol·m-2·s-1时,4个种源山桐子的Pn随着PAR增加,基本呈现线性增加的趋势,在400 μmol·m-2·s-1≤PAR≤1 200 μmol·m-2·s-1时,4个种源的Pn随着PAR增加较慢,当PAR≥1 200 μmol·m-2·s-1时,4个种源山桐子逐渐达到光饱和点,Pn随着PAR增加小幅度降低。此时,4个种源山桐子出现光抑制现象。4个种源山桐子光饱和点(LSP)为939.26~1 464.23 μmol·m-2·s-1,光补偿点(LCP)为11.38~87.76 μmol·m-2·s-1。不同种源山桐子Pn随着PAR增加的变化过程中,4个种源山桐子均在7月Pn最高,而在9月Pn最低。
2.6 不同种源山桐子叶片CO2响应曲线分析
从张家界、成都、广元、东京4个种源山桐子叶片CO2响应曲线(图6)可以看出,在0~800 μmol·mol-1的CO2摩尔分数范围内,4个种源叶片Pn随着CO2摩尔分数的升高几乎呈直线上升趋势,当CO2摩尔分数大于800 μmol·mol-1时,随着CO2摩尔分数升高,叶片Pn上升缓慢,逐渐趋于平稳,当CO2摩尔分数大于1 200 μmol·mol-1时,Pn最大,此后随着CO2摩尔分数升高,Pn开始出现小幅度下降趋势,出现CO2饱和点。4个种源山桐子CO2饱和点为1 385.36~1 527.29 μmol·m-2·s-1,CO2补偿点为19.12~146.42 μmol·m-2·s-1。不同种源山桐子Pn随着CO2摩尔分数增加的变化过程中,4个种源山桐子Pn均在7月最高,而在9月最低。
3 讨论与结论
植物光合能力的强弱反映了其对环境的适应能力[25],光合作用不仅是物质生产更是产量形成的重要因素[26],可以作为评价优良种源选择的重要指标之一[27]。本研究中4个种源山桐子间Pn均有差异,但张家界种源不同月份Pn差异较大,可能与其Pn与Tr、Gs、Ci、PAR相关性程度不同,影响Pn的主要因素差别较大。这与不同种源木棉的光合特性在种源间差异显著,且影响其Pn的因素不同[28],与不同种源羊草引种到同一地区,表现差异显著[7]的研究一致,同一树种不同种源光合特性是受温度、光照、CO2摩尔分数、经度、纬度等几个或多个因子共同作用的结果[29-30]。不同月份广元种源和成都种源Pn日变化规律相似却和张家界、东京种源日变化有差异,不同种源山桐子Pn差异可能是由于生境不同而导致的地域差异,这种差异还具有遗传效应[31]。这也为后期山桐子南北区域引种提供了理论依据,栽培方式还要考虑种源地气候。综合4个种源山桐子Pn日变化和光响应曲线,可以得出,4个种源中张家界种源对光的适应范围最大,而东京种源的Pn最高,在郑州地区东京种源可能有较强的光合能力。多种树种在不同种源间光合性能均有差异[32-33],衡量其是否为优质种源,还应结合树体自身呼吸消耗、光合产物分配、叶面积、叶面指数等指标综合判定。
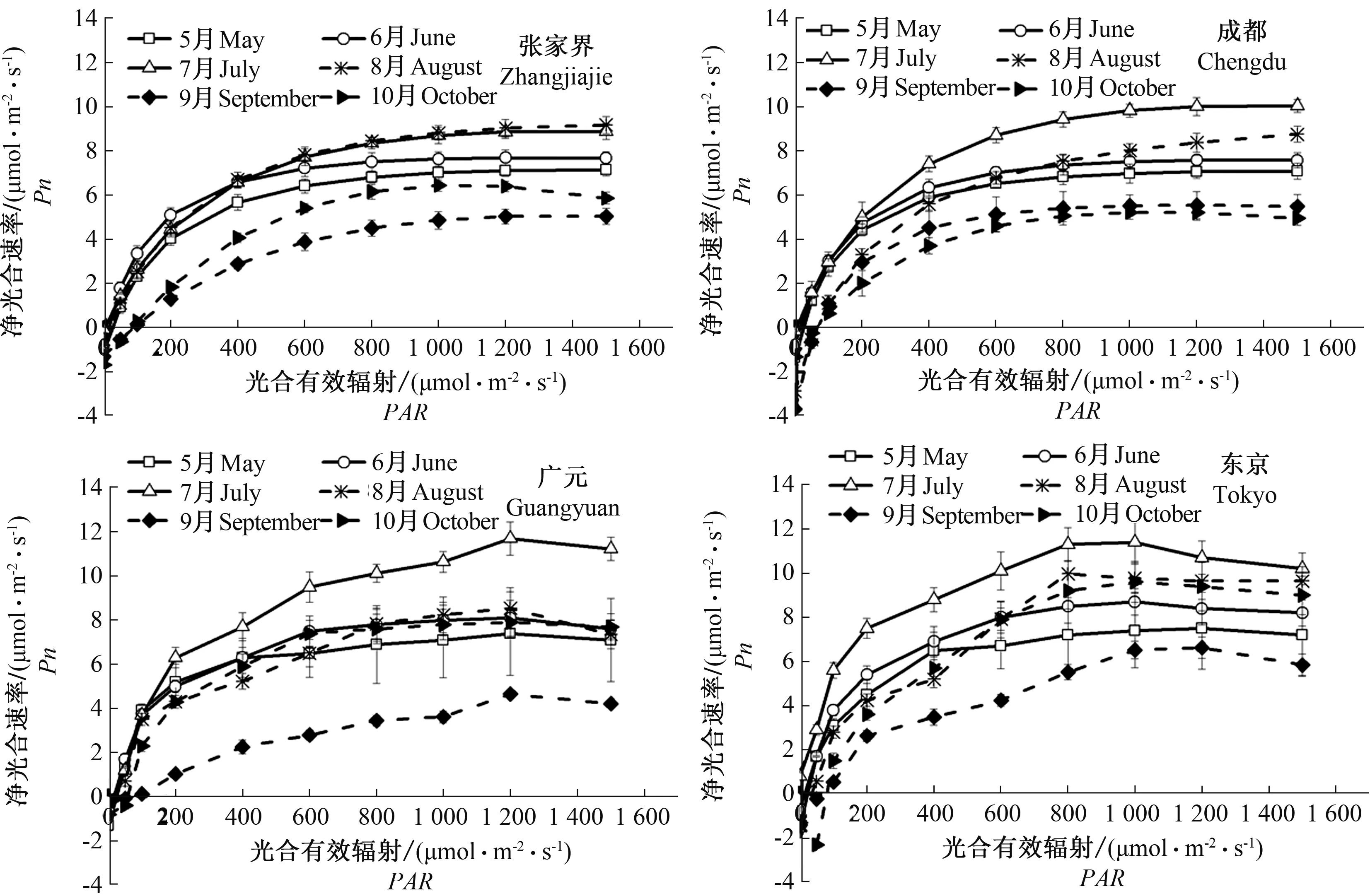
图5 不同种源山桐子Pn-PAR曲线
多种植物有光合“午休”现象,它是在植物进化过程中产生的生理现象,有利于植物适应逆境[34]。“午休”期间Pn降低的原因有气孔因素和非气孔因素[35],前者部分气孔关闭后,Ci会降低,非气孔因素一般是由于Rubisco关键酶含量和活性降低[36],此时,Pn降低后Ci会有所升高,从4个种源山桐子Pn来看,4个种源山桐子Pn不同月份日变化规律皆为双峰曲线,不同种源不同时期均有明显的“午休”现象,这与张磊等[37]在研究1 a生苗木山桐子7月份光合特性结果一致。本研究中山桐子产生“午休”现象的原因是气孔因素导致的,可能是由于夏季中午高温和强光使气孔关闭,山桐子光合作用能力减弱。高温时适当喷灌促进气孔开放以维持较高性能的光合,同时非气孔因素也有一定的影响。
植物的光响应曲线反映了植物的光化学效率[38],LSP反映了植物对强光照的利用能力,LSP值越高,说明植物利用强光照的能力越强,而LCP反映了植物在弱光条件下的光合性能,LCP值越低,说明植物在弱光下就能进行光合物质积累,一般木本植物阳性叶片光饱和点范围为600~1 500 μmol·m-2·s-1[39],光补偿点范围为10~50 μmol·m-2·s-1[40]。本研究中4个种源山桐子不同生长时期LSP均为939.26~1 464.23 μmol·m-2·s-1,LCP为11.38~87.76 μmol·m-2·s-1。与阳性植物特征研究一致,表明山桐子为阳性植物,光饱和点较高而光补偿点相对较低,因此,山桐子栽培时应选择光照充足的地方。
CO2饱和点和CO2补偿点是光合作用中植物对CO2利用能力的表现[14],CO2饱和点是反映植物利用CO2能力的重要指标,CO2补偿点越低表明植物利用低摩尔分数CO2进行光合作用干物质积累能力越强。4个种源山桐子CO2饱和点为1 385.36~1 527.29 μmol·m-2·s-1,CO2补偿点为19.12~146.42 μmol·m-2·s-1。4个种源山桐子CO2补偿点相对其他植物较高,而CO2饱和点也较高,说明山桐子利用低摩尔分数CO2的能力相对较差,而利用高摩尔分数CO2的能力相对较强。
综上所述,4个种源中张家界种源对光的适应范围最大,而东京种源的Pn最高。山桐子存在光合“午休”现象。山桐子为阳性树种,在山桐子的栽培中应选择光照充足的地方。针对不同种源的栽培管理方式还应结合种源地的气候条件进行综合评价。
