湿加松亲本间SNP遗传距离与生长性状杂种优势关系研究
2021-03-02李义良赵奋成钟岁英林昌明谭志强李福明廖仿炎吴惠姗郭文冰
李义良 赵奋成 刘 阳 钟岁英 林昌明 谭志强 李福明 廖仿炎 吴惠姗 郭文冰 王 哲
(1. 广东省森林培育与保护利用重点实验室,广州 510520;2. 广东省林业科学研究院,广州 510520;3. 广东省台山市红岭种子园,台山 529200)
湿加松(Pinus elliottii×Pinus caribaea)是湿地松(P.elliottiiEngelm. var.Elliottii)与加勒比松(P.caribaeaMorelet)的杂交后代,具有速生、丰产、抗风、抗病虫害、松脂产量高等优点,是我国华南地区重要的工业原料林树种。湿地松×洪都拉斯加勒比松(Pinus elliottiiEngelm var.elliottii×P.caribaeavar.hondurensis(Senecl.)Barrett. Et Golfari)存在显著杂种优势,我国获得的杂交子代与湿地松自由授粉子代相比,平均材积增益为221.36%[1]。因此,杂种优势高效利用已成为湿地松、加勒比松种间杂交育种中的核心关键问题,湿加松在我国华南地区推广面积达100 多万亩。高配合力亲本选择是湿加松杂种的关键,配合力的测定需要进行大量杂交制种,通过营建大规模子代测定林统计性状表现,从而获得配合力结果,过程复杂,费时费力,准确性也往往受到环境等因素影响。杂种优势的精准预测对提高湿加松杂交育种效率尤为重要。
李义良等[2]利用SSR 标记对湿加松17 个亲本遗传距离进行估算,发现杂交组合树高、胸径、材积的杂种优势与亲本间SSR 遗传距离(0.337 0~0.611 2)相关性达显著或极显著水平。张一等[3]利用ISSR标记分析马尾松亲本遗传距离与子代生长性状相关性分析,发现遗传距离在0.264~0.529时,亲本间遗传距离与子代表型值显著正相关,在0.158~0.657 时,亲本间遗传距离与子代表型值相关性降低。王茜等[4]采用ISSR分子标记对28个马尾松优良家系最优单株进行遗传分析,发现两两之间的遗传距离在0.212 1~0.565 6,遗传距离大于0.5的无性系对有11组,两两之间具有相对较远的亲缘关系,为马尾松高世代育种群体的选育提供分子水平上的依据。董虹妤等[5]利用SSR 标记分析马尾松杂交亲本间的遗传距离,除半双列Ⅱ中的胸径表型值和胸径杂种优势外,多数马尾松子代生长性状表型值及其杂种优势与GD(≤0.351 6)呈显著或极显著的正相关关系。以上在松树上的研究结果表明,SSR、ISSR 遗传距离与生长性状杂种优势存在一定的相关性。
随着全基因组测序及高通量测序技术的发展,开发覆盖全基因组海量SNP 位点已成分析物种遗传多样性和遗传进化有效手段。SLAF-seq(Specific length amplification fragment sequencing)技术通过对限制性内切酶酶切后的特异性DNA特段,以高通量测序技术获得海量序列来代表物种全基因组信息,不依赖基因组序列,已成为SNP标记开发的首选方法。应用的物种有芝麻(Sesamum indicumLinn.)[6]、大 豆(Glycinemax(L.)Merr)[7]、猕 猴 桃(Actinidia chinensisPlanch)[8]等。该技术可用于单体型图谱、遗传图谱、关联性图谱、多态性图谱的构建,为分子育种、物种进化、种质资源鉴定提供重要技术保障[9]。但在基于海量SNP 预测湿加松种间遗传距离与杂种优势关系还未见报道。利用SNP 芯片预测玉米复杂性状的杂种优势,精度达到了0.72~0.81[10]。利用SNP 芯片分析油菜遗传距离与杂种优势发现,株高、分枝部位高度和单株产量的杂种优势与遗传距离呈显著正相关关系[11]。
本研究利用SLAF 技术对湿加松亲本种质资源进行测序,开发SNP 标记,基于海量SNP 估算13个杂交组合亲本间遗传距离,分析杂交组合生长性状的杂种优势,进而探讨湿加松亲本间SNP 遗传距离在杂种优势关系,为湿加松分子辅助交配设计育种提供参考。
1 试验材料方法
1.1 试验材料
131个湿加松亲本种质资源,包括湿地松种质54 个、洪都拉斯加勒比松种质59 个、古巴加勒比松种质12 个、巴哈马加勒比松种质6 个,其中湿地松种质A06、B20、B118、SS05-2,洪都拉斯加勒比松种质ZH03、HL17、HQ09、HQ27 是13 个湿地松×洪都拉斯加勒比松杂交组合(A06×HL17、A06×HQ09、A06×ZH3、B118×HL09、B118×HL17、B118×HQ09、B118×ZH3、B20×HL17、B20×HQ09、B20×ZH3、SS05-2×HL17、SS05-2×HQ09、SS05-2×ZH3)的亲本。试验材料取自台山市红岭种子园,取成熟针叶,于液氮中速冻后放-80℃冰箱保存备用。
1.2 子代测定
湿加松子代测定林1999年种植于台山,随机完全区组设计,5株单行小区,8次重复,株行距3 m×3 m。2006年对湿加松子代测定林进行每木调查,测量了13 个湿地松×洪都拉斯加勒比松杂交组合树高(H/m)、胸径(D/cm)。利用树高、胸径计算单株材积(V)(V/m3):
式中:π为圆周率;f为形数,取f=0.5。
杂种优势程度(H)计算公式为:

1.3 DNA的提取
采集131个湿加松亲本种质当年生针叶,基因组DNA 提取采用DNeasy Plant Mini Kit 试剂盒(QIAGEN),DNA 的完整性用1%的琼脂糖凝胶电泳进行检测,紫外吸收法检测基因组DNA 的浓度及纯度。DNA 样品送至百迈客生物科技有限公司利用SLAF 技术进行SNP 标记开发,包括酶切、测序数据统计与评估、实验建库评估、SNP 标记开发等步骤。
1.4 数据分析
通过完整度0.5 次要基因型频率0.05 对群体SNP 进行过滤,质量控制后得到的SNP 位点用于遗传进化分析。利用MEGA5.0软件进行亲本间的遗传距离和亲本聚类分析。利用SAS 软件计算株高、胸径、材积生长表现及杂种优势,SPSS22.0 软件进行遗传距离与株高、胸径、材积的相关性分析。
2 试验结果
2.1 湿加松亲本SNP标记开发
对火炬松参考基因组进行电子酶切预测,确定限制性内切酶切组合为EcoRV+ScaI,对131 个湿加松亲本种质资源进行测序,共获得110.97 M reads 数据,测序平均Q30 为86.68%,平均GC 含量为36.84%,达到测序要求。通过SOAP 软件将Control(水稻日本晴)的测序reads与其参考基因组进行比对,双端比对效率在95.89%,酶切效率为98.88%,SLAF 建库正常。共开发668 656 个SLAF标签,平均测序深度为128.62 x,多态性SLAF 标签有53 952 个。SNP 标记的开发是以每个SLAF 标签中深度最高的序列类型作为参考序列,利用BWA 将测序reads 比对到参考基因组上,并使用GATK 和samtools 两种方法开发SNP,以两种方法得到的SNP 标记交集作为最终可靠的SNP 标记数据集,共得到206 219个群体SNP。通过完整度0.5次要基因型频率0.05 对206 219 个群体SNP 进行过滤,最终得到96 736 个SNP 位点用于遗传进化分析。
2.2 种间杂交亲本间遗传关系分析
基 于96 736 个SNP 位 点,通 过MEGA 5.0 软件,计算13 个湿地松×洪都拉斯加勒比松杂交组合亲本间遗传距离(见表1),结果表明,4个湿地松和4 个洪都拉斯加勒比松间的遗传距离变幅为0.084 6~0.490 6,平均为0.319 4,其中两个湿地松B20 与B118 遗传距离最小为0.084 6,湿地松A06与洪都拉斯加勒比松HL17 的遗传距离最大为0.490 6。湿地松间的平均遗传距离为0.110 6明显小于与洪都拉斯加勒比松间的平均遗传距离为0.244 0,但均小于湿地松与加勒比松间的平均遗传距离为0.460 2。在13 个种间杂交组合中,亲本间遗传距离介于0.425 1~0.490 6,A06 与HL17 的遗传距离最大为0.490 6,SS05-2与HQ09的遗传距离最小为0.425 1,平均遗传距离为0.460 2,遗传距离的变异系数为10.59%,表明不同杂交组合的亲本间遗传距离存在明显差异。

表1 8个亲本间SNP遗传距离Table 1 SNP genetic distance among eight parents
利用MEGA5.0 软件,基于SNP 位点对8 个亲本进行聚类分析,8 个亲本被分为2 个组群(见图1)。4 个湿地松A06、B20、B118、SS05-2 聚为一类,4 个洪都拉斯加勒比松HL09、HL17、HQ09、ZH03聚为一类。反映了亲本间的亲缘关系,与树种实际情况相符。
2.3 杂交组合生长性状表现
13 个杂交组合3 个生长性状的表现如表2,树高变幅为8.88~9.91 m,A06×HL17 组合最高,SS05-2×HQ09 组合最低;胸径变幅为14.36~16.73 m,A06×HL17 组合最高,SS05-2×HQ09 组合最低;材积变幅为0.080 9~0.111 8 m3,A06×HL17 组合最高,B20×HL17组合最低。
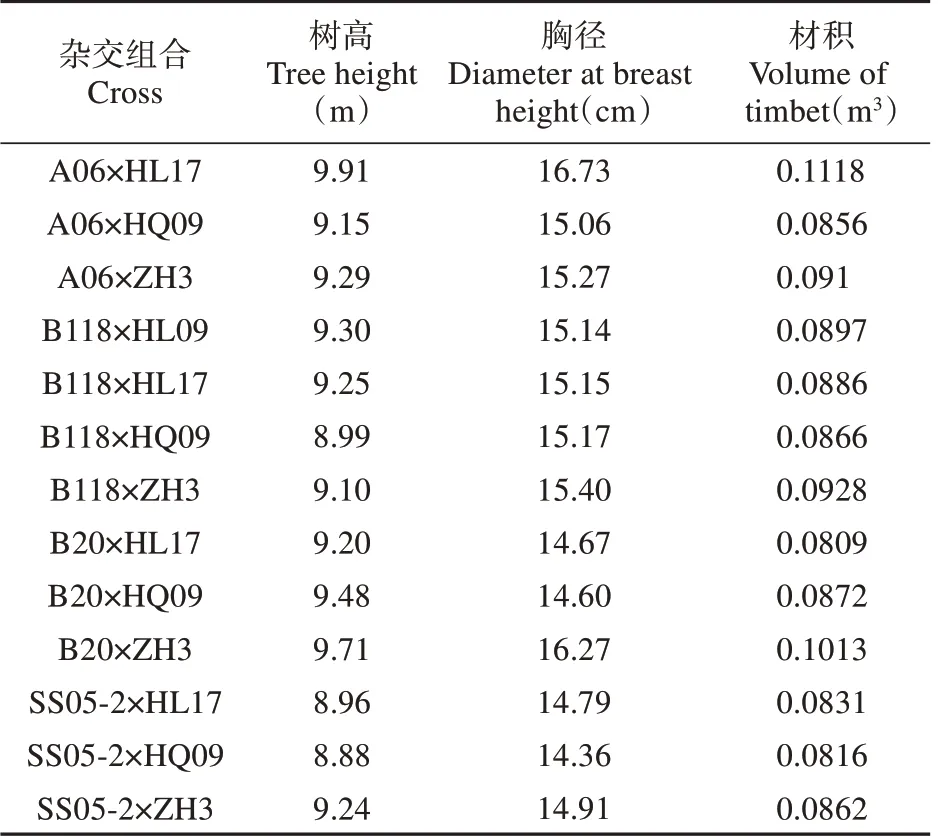
表2 13个杂交组合生长性状平均表现Table 2 The average performance of thirteen hybrid combinations
2.4 杂交组合生长性状杂种优势分析
对13 个杂交组合3 个生长性状杂种优势进行分析,发现13 个杂交组合树高、胸径、材积均具有正向杂种优势,均值分别21.92%、57.65%、190.30%(见表3)。3 个性状中材积杂种优势最明显,变异范围为,变幅为161.85%-261.74%,A06×HL17杂种优势最高为261.74%;其次是胸径,变幅为49.03%~73.58%,A06×HL17 杂种优势最高为73.58%;树高杂种优势最低,变幅为16.87%~30.45%,依然是A06×HL17杂种优势最高为30.45%。

表3 13个杂交组合生长性状杂种优势分析Table 3 Analysis of heterosis in growth traits of thirteen hybrid combinations
2.5 亲本SNP 标记遗传距离与杂交组合生长性状杂种优势相关性分析
利用SPSS 软件计算13 个杂交组合3 个生长性状杂种优势与遗传距离相关系数,发现湿加松亲本间SNP 遗传距离与树高、胸径、材积生长性状杂种优势相关性均达到显著正相关水平,遗传距离与树高相关性最高为0.680;与材积相关性最低为0.624。

表4 生长性状杂种优势与遗传距离之间的相关关系Table 4 Correlation between heterosis of growth traits and genetic distance
3 讨论
SLAF-seq 等简化基因组测序技术为无参考基因组物种标记开发提供了一条新的途径,目前已应用于多个物种标记开发,这些技术通过酶切后测序,获得海量SNP。本研究利用SLAF-seq 技术对131个湿加松亲本种质资源进行测序,筛选到有效多态性SNP 位点96 736 个,基于海量SNP 位点获得湿加松亲本间遗传距离,并进行聚类,与树种实际情况相符,即4 个湿地松A06、B20、B118、SS05-2 聚为一类,4 个洪都拉斯加勒比松HL09、HL17、HQ09、ZH03 聚为一类。李义良等[2]利用SSR分析的17个湿加松亲本间遗传距离在0.233 6~0.611 2,平均遗传距离为0.429 0,遗传距离的变异系数为20.90%。本研究SNP遗传距离变幅(0.425 1~0.490 6)低于SSR 遗传距离变幅,SNP 遗传距离变异系(10.59%)数也低于SSR 遗传距离变异系数。与SSR 相比SNP 聚类结果也更加清晰,表明在湿加松亲本间遗传距离分析方面,SNP 标记更准确,与SSR标记相比SNP标记更加有效,优势明显。
近年来,利用SSR、ISSR、AFLP、RAPD 等分子标记估算遗传距离预测杂种优势开展了广泛研究,出现了相关性显著[12]、一定范围内相关[3,5,13~14]、相关系数较小[3,15~16]、相关性不显著[17]、预测性较差[18]、不能有效预测[19~20]等结论,出现以上结果可能与所用种质材料有关[21],也可能与标记覆盖基因组的范围、标记与预测性状的关联度、随机标记的分布等因素有关[21~23]。张一等[3]等认为马尾松亲本间ISSR分子遗传距离对于子代生长性状相关系数较小,预测作用较弱。李义良等[2]认为SSR 标记遗传距离与树高、胸径、材积的杂种优势相关程度较高(相关系数分别为0.542 0、0.641 2、0.639 3),利用SSR 标记预测湿加松杂种优势是可行的。董虹妤等[5]发现SSR 标记估算的遗传距离与马尾松子代生长性状杂种优势相关性较高,可以利用一定水平的遗传距离对马尾松子代生长性状杂种优势进行有效预测。在作物研究中发现,SNP 芯片能够高效预测玉米复杂性状杂种优势[10],利用60K SNP 芯片分析亲本的遗传关系预测油菜杂种优势的效果显著[11],SNP 芯片比其他标记预测效果好。本研究基于96 736 个SNP 位点估算出13个杂交组合亲本间遗传距离,与树高、胸径、材积生长性状杂种优势呈显著正相关关系,相关系数分别为0.680、0.648、0.624。与SSR 标记相比,本研究所用SNP 标记不仅数量多,而且是在整个基因组范围内筛选获得的多态性标记,比传统标记覆盖度高,利用海量SNP 位点估算湿加松亲本间遗传距离更加准确,能够有效预测树高、胸径、材积杂种优势。因此,应选择SNP 遗传距离相对较远的湿加松亲本配置杂交组合,提高良种选育效率。
