利用同源重组技术构建Chordin 基因载体及其在成纤维细胞中表达量的研究
2021-03-01刘开东贺建宁张梦瑶
刘开东,荣 恒,贺建宁,张梦瑶,赵 明,柳 楠*
(1.青岛农业大学动物科技学院,山东青岛 266109;2.青岛市畜牧兽医研究所,山东青岛 266000;3.青岛市动物疫病预防控制中心,山东青岛 266000)
羊毛是毛囊的衍生物,毛囊是表皮向内凹陷,通过相互作用,在各种信号通路的协作下形成的[1]。近年来随着人们对羊毛质量要求的提高,对毛囊的研究显得尤为重要[2]。敖汉细毛羊是我国知名的肉毛兼用羊,具有羊毛细、柔软度高、体格大等特点,是理想的研究对象[3]。在模式小鼠中,Puas 等[4]在20 年前对于毛囊的发生发育给出一套分类方法,将其分为3 个时期8 个阶段;随着近些年遗传学手段及二代测序技术的发展,Sexna[5]将原本的分类方法进行改进,划分为10 个阶段。关于羊毛囊的研究目前主要集中在出生后和成年绵羊身上[6]。Stenn 等[7]在成年绵羊身上对毛囊的周期性生长变化展开研究;柳楠等[8]在分子水平上研究了敖汉细毛羊羊毛性状形成的机理;王小佳等[9]在分子水平上探究了羊毛的生长部位。大多数研究都集中在分子水平上,而在细胞水平上与毛囊生长发育相关的研究甚少。因此,本研究通过对其质粒的构建及转染成纤维细胞来进行Chordin基因表达量的分析,为该基因在个体水平上的研究提供基础。
Chordin基因是骨形态发生蛋白信号通路中的拮抗剂[10],Chordin 蛋白与BMP2和BMP6的亲和力较强,能够与TSG、BMP 结合形成蛋白三聚体[11],进而阻止BMP2和BMP6与其受体的结合[12]。许多研究表明,BMP 家族中基因抑制毛囊发育[13],Chordin作为此家族的拮抗剂,能够间接促进毛囊发育。早期研究表明,BMP 信号通路对毛囊的形态发生及周期性发育有重要的调控作用,秦波等[14]研究发现在团头鲂胚胎发育的各个时期都可检测到Chordin基因,其对胚胎早期发育起到了重要的调控作用。Chordin基因在12 个组织中的脑组织表达量较高,推测其参与了脑组织发育过程,对神经系统的发育有较大影响[15]。由此推断Chordin基因可以通过与BMP 结合进而对毛囊发育过程起调控作用。
目前,关于Chordin调控机制研究主要集中在人类、小鼠、斑马鱼等动物中,在细毛羊中与毛囊相关的研究较少。因此,本实验以敖汉细毛羊为研究对象,采集40 日龄的胎羊,通过对Chordin基因表达载体的构建,利用实时荧光定量PCR 和Western Blot 等技术分析其转染成纤维细胞后表达量的变化,旨在为Chordin基因的功能研究奠定基础。
1 材料与方法
1.1 实验材料 选取40 日龄的健康胎羊(由青岛畜牧所奥特种羊场提供)并及时消毒处理。TRIZol(Roche)试剂、蛋白裂解液、EcoRI限制性内切酶、T4 DNA 连接酶、pcDNA3.1 质粒、DH5α感受态细胞、DMEM(基础培养基)、胰蛋白酶、标准胎牛血清、DPBS(磷酸缓冲盐溶液)、Lipofectamine 2000、反转录试剂盒、SoSo 试剂盒、切胶回收试剂盒、质粒提取试剂盒、培养皿等均购自青岛尚赛科贸有限公司。
1.2 实验方法
1.2.1 引物设计 从组织中提取RNA 并反转录成cDNA。在绵羊Chordin基因序列(登录号为XM-027957560.1)CDS 区外侧通过Primer 5.0 设计引物,并在上游引物和下游引物的5' 加入同源臂。完成后的引物序列为:F:5'-TTTAAACTTAAGCTCGGAATTCCC CCAGCTGTCCCGTTCG-3';R:5'-TGGATCCGAGCTC GGCTAGGAGCCTTCATCTTCTTTCCCAGAC-3'。通过PCR 扩增后进行琼脂糖凝胶电泳检测,体系25 μL:DNA 模板1 μL,上下游引物各1 μL,金牌Mix 22 μL。
1.2.2Chordin基因与pcDNA3.1 质粒同源重组对pcDNA3.1 质粒进行EcoRI单酶切,酶切体系为:EcoRI1 μL、pcDNA3.1 5 μL、Buffer 5 μL、ddH2O 39 μL,置于37℃金属浴中2 h。电泳检测后,通过切胶回收试剂盒纯化Chordin基因及切开的pcDNA3.1,通过T4 DNA连接酶进行连接,10 μL 体系:Chordin基因1 μL、pcDNA3.1 1 μL、SoSo 5 μL、ddH2O 3 μL,置于50 ℃金属浴10 min。之后转至DH5α感受态细胞,选取摇菌后的单菌落对其进行阳性率测定,经测序、质粒提取最终获得阳性质粒[16]。
1.2.3 成纤维细胞的培养 在羊场取40 日龄胎羊,用75% 酒精清洗2 遍,而后转至实验室,在培养皿中用75% 的酒精再次清洗3 遍,最后用含有双抗的PBS 反复清洗3 遍。用剪刀剪掉头部和四肢,然后将躯干在体视镜下剪成1.5 mm3左右(米粒大小),将其放在培养皿,加胎牛血清,在CO2浓度为0.05、温度37℃的培养箱中培养4 h,再加培养液。从培养箱拿出24 h 后,用PBS 冲洗干净,更换培养液,定时观察。细胞生长到95%左右进行传代培养[16]。
1.2.4 细胞转染 原代培养后传至第二代(P2)。用Lipofectamine 2000 进行瞬时转染,实验组、对照组分别加20 μL 脂质体试剂和480 μL DMEM(不含双抗),混匀,静置10 min,在实验组中加入40 μL 构建好的表达载体和460 μL DMEM(不含双抗),对照组仅加入500 μL DMEM,静置20 min,然后将实验组和对照组都移到培养皿中均加入4 mL DMEM,在37℃的温度条件下,培养6 h 后换液,之后培养36 h[16]。
1.2.5 细胞转染后的RT-PCR 扩大培养转染成功的单细胞,细胞在培养皿底部长到95%时,每个板加TRIZol(Roche)试剂3 mL,细胞裂解后进行RNA 提取。提取完成后,测定浓度和纯度符合标准后将其反转录成cDNA[17]。通过Primer Premier 5.0 软件设计Chordin和GAPDH基因的引物序列,完成后其引物序列见表1。
通过实时荧光定量PCR 反应测定表达量,实验组和对照组各3 次重复,Chordin相对于内参基因GAPDH的表达量用Ct(2-ΔΔCt)值法计算。利用SPSS 17.0 中t检验进行数据分析[17]。
1.2.6 细胞转染后Western Blot 将样品分为实验组和对照组,每组3 个生物学重复。先裂解提取细胞中的蛋白质,内参基因GAPDH、目的基因Chordin分别点3个胶孔,电泳后对蛋白质进行PVDF 转膜,用转膜电泳槽进行2~3 h,之后封闭蛋白质2 h;TBST 洗膜3 次,每次5 min;按1:2 000 的比例对一抗进行稀释,在4℃冰箱用一抗孵育PVDF 膜12 h;用TBST 洗膜3 次,每次5 min。以1:2 000 的比例稀释二抗,孵育1.5 h,在暗室中曝光。用ImageJ 软件分析条带,结果用SPSS软件进行分析[17]。

表1 引物信息
2 结果
2.1Chordin目的基因扩增 图1 为PCR 扩增后凝胶电泳分析结果(2%琼脂糖凝胶),扩增条带位于2 874 bp 处,与已知大小一致。
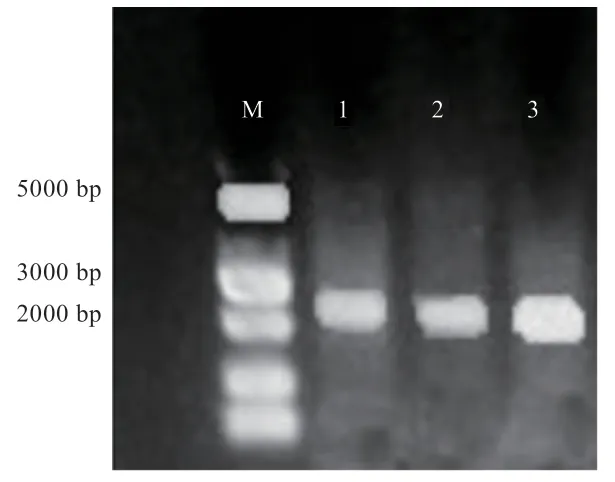
图1 Chordin 基因PCR 扩增结果
2.2 pcDNA3.1 载体单酶切及纯化 图2 为pcDNA3.1 单酶切结果,1~2 酶切效果良好,pcDNA3.1 载体5 428 bp,与图中条带所吻合,可用于后续实验。

图2 pcDNA3.1 载体单酶切鉴定
2.3 表达载体pcDNA3.1 与目的片段重组及鉴定 如图3 所示,在2 874 bp 处有单一明亮的条带,为Chordin基因。菌液测序后用BLAST 比对,结果如图4 所示,碱基未发生突变,可用于后续实验。提取重组质粒,结果如图5 所示,1~2 为pcDNA3.1-Chordin重组质粒,重组后质粒大小为8 302 bp,与图中条带相符,表明重组质粒构建成功。
2.4 细胞培养 电子倒置显微镜下,定时观察。24 h 后细胞逐渐贴壁,组织块的四周逐渐有细胞游出。传代培养在细胞密度约为95%时进行,采用胰蛋白酶消化法,2 000 r/min 离心处理分离成纤维细胞,之后扩大培养,观察结果如图6 所示。

图3 菌液PCR 扩增结果

图4 测序比对

图5 重组质粒的提取
2.5 实时荧光定量PCR 检测mRNA 的表达 琼脂糖凝胶电泳检测结果表明,扩增的条带单一明亮,GAPDH产物大小为125 bp、Chordin产物大小为172 bp(图7),与已知大小均相同,可继续实时荧光定量PCR 实验。

图6 成纤维细胞原代、传代培养

图7 内参基因GAPDH(A)和目的基因Chordin(B)扩增产物
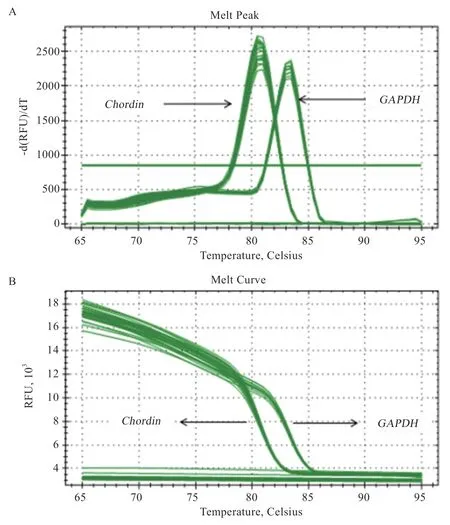
图8 Chordin 和GAPDH 熔解曲线(A)和扩增曲线(B)图
如图8 所示,Chordin和内参基因GAPDH的扩增曲线、熔解曲线清晰且只有一个峰值,结果表明在实时荧光定量PCR 过程中无二聚体且得到特异性的扩增产物,可用来进行定量分析。Chordin基因经过质粒构建转染成纤维细胞后其表达量极显著高于对照组(图10)。
2.6 Westren Blot 检测蛋白的表达 由图10 可知,转染的实验组条带明显比未转染的对照组粗,其灰度值由Image J 软件进行分析,结果用软件SPSS17.0 分析。由图11 可知,转染组Chordin基因蛋白的表达显著高于未转染的对照组,表明质粒pcDNA3.1-Chordin能高效表达Chordin基因。

图9 Chordin 基因mRNA 相对表达量

图10 Chordin 蛋白表达条带

图11 Chordin 蛋白相对表达量
3 讨 论
多种研究迹象表明,Chordin可作为BMP 基因家族的拮抗剂存在。Elisa[18]在牛中发现Chordin可以被BMP1和tolloid调节,其中BMP1对Chordin的调节具有抑制作用,但不能诱导骨形态发生过程。Kane 等[19]研究发现缺氧缺血性视网膜病变是视网膜新生血管形成的主要原因,低氧可诱导人视网膜周细胞中Chordin的表达,BMP6有效地参与了血管生成,Chordin在缺氧条件下可通过调节BMP6对内皮细胞的作用而对视网膜周细胞发挥作用,从而可以促进新生血管的形成。Fabian 等[20]实验发现可通过刺激BMP 通路保护肾脏免受急性或慢性损伤,Chordin特异性拮抗BMP7,BMP7在相邻的远端肾单位中表达,Chordin在人的缺血敏感S3 节段中表达,当该节段退变时,Chordin表达减少,BMP 信号的表达也相应减少。结果表明,Chordin与健康近端小管中的BMP7信号拮抗,降低其表达量,同时导致上皮损伤,损伤的上皮脱落能促进BMP7信号在上皮中再次表达,进而保护肾脏。Mikawa[21]等发现Chordin在大多数神经元以及它们的树突和轴突中被强烈表达,在高可塑性灰质的神经纤维中也观察到丰富的Chordin表达,例如小脑的分子层和上丘的浅层。Piccolo 等[22]在非洲蟾蜍体中发现Chordin 蛋白与BMP2和BMP6的亲和力较强,进而阻止了BMP2和BMP6与其受体的结合 。
Petrky 等[12]研究发现Chordin通过与BMP 结合而阻断其与受体结合,进而阻断BMP 信号通路。综上所述,Chordin可与BMP 特异性结合,干扰BMP 与其受体结合进而发挥调控作用,因此推测Chordin可以通过与BMP 结合进而对毛囊发育过程起调控作用。鉴于此,本实验通过对pcDNA3.1-Chordin载体的构建,通过其表达量的变化,在细胞层面上检测Chordin的功能,推断其与毛囊生长发育间的关系。
4 结 论
本研究通过对pcDNA3.1-Chordin载体的构建,通过实时荧光定量PCR、Western Blot 技术检测其转染成纤维细胞后的表达量,证实Chordin基因过表达,其mRNA 表达量和蛋白的表达量与对照组相比显著提高。
