不同播深环境下玉米耐深播性状杂种优势及遗传效应解析
2021-02-22赵小强李文丽周文期
钟 源 赵小强 李文丽 张 丹 周文期
(1甘肃农业大学,甘肃省干旱生境作物学重点实验室, 甘肃 兰州 730070;2甘肃省农业科学院作物研究所, 甘肃 兰州 730070)
我国三分之二的玉米(Zea mays)主要种植分布于东北、华北及西北等依靠自然降水的旱作区[1],玉米播种时这些区域常年0 ~5 cm 表土蒸发量大,土壤含水量和降水量低,90%的年份因墒情不足严重影响了玉米适时播种及种子正常出苗生长[2-3]。目前,一般的玉米品种因其顶土能力和破土出苗能力差,在深播下即使发芽也不能把幼芽送出地表,造成玉米缺苗断垄达40%~50%[2,4],严重制约玉米稳产增产。因此,培育耐深播玉米品种并及时赶墒深播,是一项拓宽玉米耐深播抗旱育种的新策略和节本高效的轻简栽培措施。
玉米耐深播性属于典型的数量遗传性状,受复杂遗传体系控制[5-7],深入理解玉米耐深播性状的遗传机理,对指导玉米耐深播抗旱育种和提高玉米产量具有重要意义。目前已在玉米Bin7.04、 Bin9.03、Bin10.03 处找到调控中胚轴长的GRMZM2G133836[4]、GRMZM2G092174[8]、GRMZM2G149580[9]基因。Troyer[6]最早采用易位测交材料在玉米3、6 和9 号染色体上找到了3 个调控中胚轴长的显性位点;后来Zhang 等[10]用F2∶3群体在10 和20 cm 播深下共检测到40 个调控中胚轴长、胚芽鞘长、苗长及出苗率的加性数量性状基因座(quantitative trait loci, QTLs)位点;Liu 等[4]还用IBM Syn10 DH(doubled haploid)群体在2 cm 和12.5 cm 播深下共定位到55 个调控发芽率、苗长、中胚轴长及胚芽鞘长的加性QTLs 位点。此外,曹立勇等[11]用DH 系在2 种低温环境下共识别到7 个调控水稻(Oryza sativa)中胚轴长的加性QTLs 和2 对加性×加性上位性QTLs 位 点。Spielmeyer 等[12]还 用RILs(recombinant inbred lines)群体分析了小麦(Triticum aestivum)6A 染色体上调控胚芽鞘长的加性主效QTLs位点。从这些研究可以看出,玉米耐深播性状QTL 定位,特别是QTL 与环境互作及上位性互作位点的深入分析还不够深入。
在Cockerham 遗传模型和MINQUE 法基础上,Zhu[13]提出了研究动植物基因型与环境互作的数量性状加性-显性-母体遗传模型(additive-dominantmaternal effect genetic model, ADM),为调控玉米耐深播性遗传效应的系统解析提供了可行方法。基于以上考虑,本研究以耐深播性不同的22 份玉米亲本及组配的22 份F1为试材,采用ADM 模型,分析3 种播深下玉米13 个耐深播性状的遗传效应;根据各性状的杂种优势表型及遗传效应预测值,选用遗传主效应表现良好并在不同播深下互作效应表现稳定的优良亲本有目的地组配F1,旨在提高育种选择效率,为玉米耐深播抗旱育种奠定理论依据。
1 材料与方法
1.1 试验材料
以耐深播性不同的黄早四(P1)、昌7-2(P2)、H21(P3)、JW183(P4)、A413(P5)、K12(P6)、N192(P7)、W182BW(P8)、 掖478 (P9)、65232 (P10)、 Ca317(P11)、8723(P12)、TS141(P13)、TS161(P14)、廊H(P15)、A654(P16)、齐319(P17)、A154(P18)、XZX227(P19)、Q1723(P20)、8802A(P21)、综31(P22)共22 份亲本,及按不完全双列杂交设计组配的22 份F1为试验材料。其中P1~P8为Tangsipingtou(TSPT)种质,P15~P16为Lancaster (Lan)种质,P21~P22为Luda-Red -Cob(LRC)种质,P17~P20为P 群种质,其余为Reid 种质[3,5,14-16]。
1.2 试验方法
采用本研究设计的种子深播试验装置(http:/ /epub.cnipa.gov.cn/patentoutline.action)进行深播试验。试验时先往该种子深播试验装置(高50 cm,内径15 cm)中分层装入灭菌蛭石,再将提前浸种12 h 的亲本及F1种子各15 粒播于灭菌蛭石上,最后分别盖3、15和20 cm 灭菌蛭石使该种子深播试验装置蛭石达50 cm,每隔3 d 浇20 mL 超纯水,3 次重复。待种子在人工气候室(每天光照12 h,光照强度600 μmol.s-1.m-2, 恒温20±0.5℃)萌发10 d 后统计出苗率(seeding emergence rate,RAT)。每处理选择长势整体一致的5 株幼苗测定中胚轴长(mesocotyl length,MESL)、胚芽鞘长(coleoptile length, COLL)、苗长(seeding length, SDL)、根长(root length, RL)、中胚轴粗(mesocotyl coarse, MESC)、胚芽鞘粗(coleoptile coarse,COLC)、苗重(seeding weight,SDW)、中胚轴重(mesocotyl weight, MESW)、胚芽鞘重(coleoptile weight, COLW)和根重(root weight, RW),并计算中胚轴与胚芽鞘和(mesocotyl and coleoptile total length,MESL+COLL)及中胚轴与胚芽鞘比(mesocotyl length/coleoptile length ratio, MESL/COLL)。
1.3 数据统计与分析
采用IBM SPSS 19.0 (https:/ /www. ibm. com/products/spss-statistics) 中 的 GLM、 Reduction 及Correlate 模块对3、15 和20 cm 播深下亲本及F1的上述13 个性状进行ANOVA、Pearson 及PCA(principal component analysis)分析。参照赵小强等[17]的方法估算13 个性状的杂种优势,即:

式中,F1为杂交种值,MP 为双亲均值,PH为高值亲本值。采用隶属函数法[5]计算3 种播深下亲本及F1的13 个性状耐深播隶属值,并进行耐深播性综合评价,即:
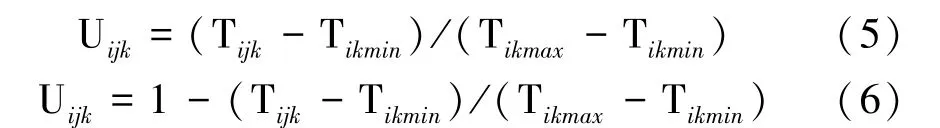
式中,Uijk为第i播深下第j材料第k性状的耐深播隶属值,Tijk为第i播深下第j材料第k性状值,Tikmax为第i播深下第k性状最大值,Tikmin为第i播深下第k性状最小值。若所测性状与玉米耐深播性呈正相关,则用式(5),反之用式(6)。计算各播深下所有性状的算术耐深播隶属值进行比较,值越大玉米耐深播性越强。参照ADM 模型分析玉米13 个性状的遗传效应。采用QGA Station(http:/ /ibi.zju.edu.cn/software/qga)无偏估算各性状的方差分量占表型方差的比率及遗传力。采用AUP(Adjusted-Unbiased-Prediction)估算各性状的遗传效应值[18];采用基因型为抽样单位的Jackknife 抽样方法[18]估算各性状统计量的标准误,并检验各遗传参数的显著性。
2 结果与分析
2.1 播深对玉米耐深播性状的响应
由表1 可知,随着播深增加,22 份亲本及22 份F1的RAT、SDL、RL、MESC 和RW 整体呈降低趋势,其余性状整体呈升高趋势。但不同亲本及F1各性状间的变化幅度不同,与3 cm 播深相比,15 cm 播深下亲本及F1的RAT、MESL、COLL、MESL + COLL、MESL/COLL、SDL、RL、MESC、COLC、SDW、MESW、COLW 及RW 分别平均变化了22.94%、-114.12%、-48.02%、-81.32%、 - 56.00%、 14.71%、 16.26%、 8.99%、 -8.89%、-20.17%、-132.56%、-172.11%和10.02%,20 cm 播深下亲本及F1的这13 个性状分别平均变化了44.93%、 - 139.47%、 - 61.27%、 - 100.47%、 -58.94%、 15.65%、 16.73%、 6.14%、 - 15.37%、 -40.28%、-131.93%、-94.00%和16.46%。

表1 3 种播深下亲本及F1 耐深播性状比较Table 1 Comparison of deep-seeding traits of parents and F1 at three depths
2.2 玉米耐深播性状差异分析
3 种播深下所有亲本及F1的13 个性状联合方差分析表明(表2),亲本及F1的13 个性状在校正模型间均达到极显著差异水平(P<0.01),说明这13 个性状整体方差分析有效。此外,亲本及F1所有性状在基因型间及播深间均存在显著或极显著差异,而除SDL、RL、MESW、COLW 外,其余性状均受基因型×播深互作的显著或极显著影响。说明这13 个性状总体显著受玉米基因型自身遗传基础、播深及基因型×播深互作的影响,可能这些性状间的相互作用决定玉米耐深播性的强弱。
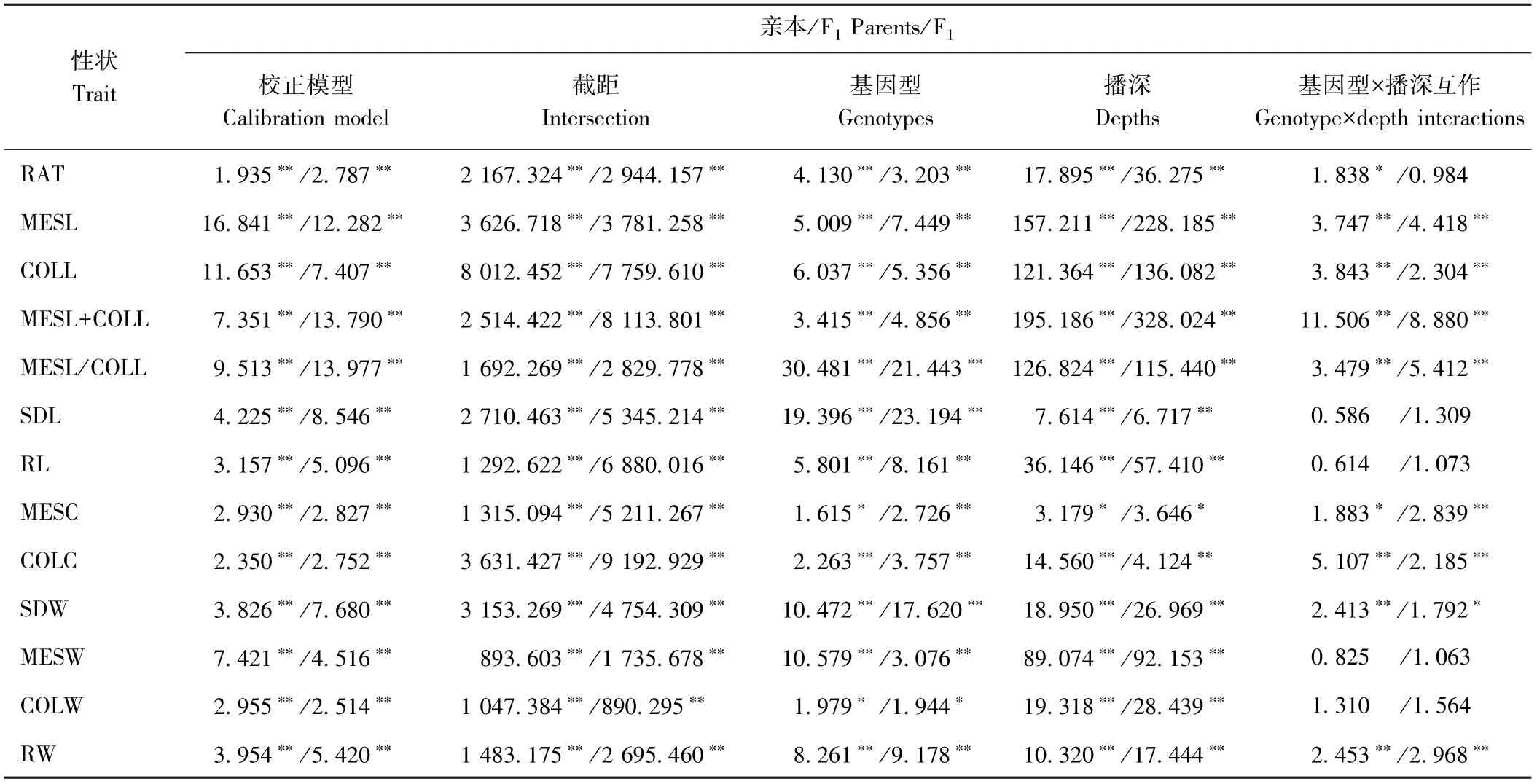
表2 3 种播深下亲本及F1 耐深播性状联合方差分析(F 值)Table 2 Combine variance analysis of deep-seeding traits of parents and F1 at three depths (F value)
2.3 玉米耐深播性状间的关系
3 种播深下所有亲本和F1的13 个性状的Pearson相关分析表明(图1),1 个单-性状至少与其余6 ~12性状呈显著或极显著正/负相关。表明玉米这13 个性状间的相互协同或拮抗作用共同决定了玉米的耐深播性强弱。此外,除SDL 外,其余性状均与播深呈极显著或显著正/负相关(图1)。说明玉米的这些性状受播深影响显著。
进一步对3 种播深下所有亲本及F1的13 个性状进行PCA 分析,结果表明(图2),13 个主成分中前3个主成分的特征值均大于1,总体方差累计贡献率达77.738%。表明前3 个主成分能代表玉米耐深播特性的绝大部分信息。其中决定第1 主成分的主要是MESL+COLL、MESL、MESW、SDW、RW 及RAT,可概括为玉米中胚轴、幼苗及根系综合生长因子,解释47.550%的贡献率。决定第2 主成分的主要是COLL、COLC 及COLW,可概括为玉米胚芽鞘生长因子,解释19.057%的贡献率。决定第3 主成分的主要是SDL 和RL,可概括为玉米幼苗及根系综合生长因子,解释11.131%的贡献率。
2.4 玉米耐深播相关性状的杂种优势分析
3 种播深下玉米13 个性状杂种优势分析表明(图3),13 个性状的平均F1杂种优势指数介于90.97%~175.64%,CV 为16.67%;平均相对杂种优势介于-16.49%~34.58%,CV 为99.77%;平均中亲优势介于-9.03%~75.65%,CV 为85.74%;平均超亲优势介于-17.78%~53.48%,CV 为212.96%。说明玉米这13 个耐深播性状间的杂种优势差异较大,今后育种应充分利用玉米这些耐深播性状间的杂种优势表现,尽可能地发挥相应性状的正向杂种优势,以便组配出强耐深播杂交种。
2.5 玉米耐深播性状的遗传方差分量和遗传力分析
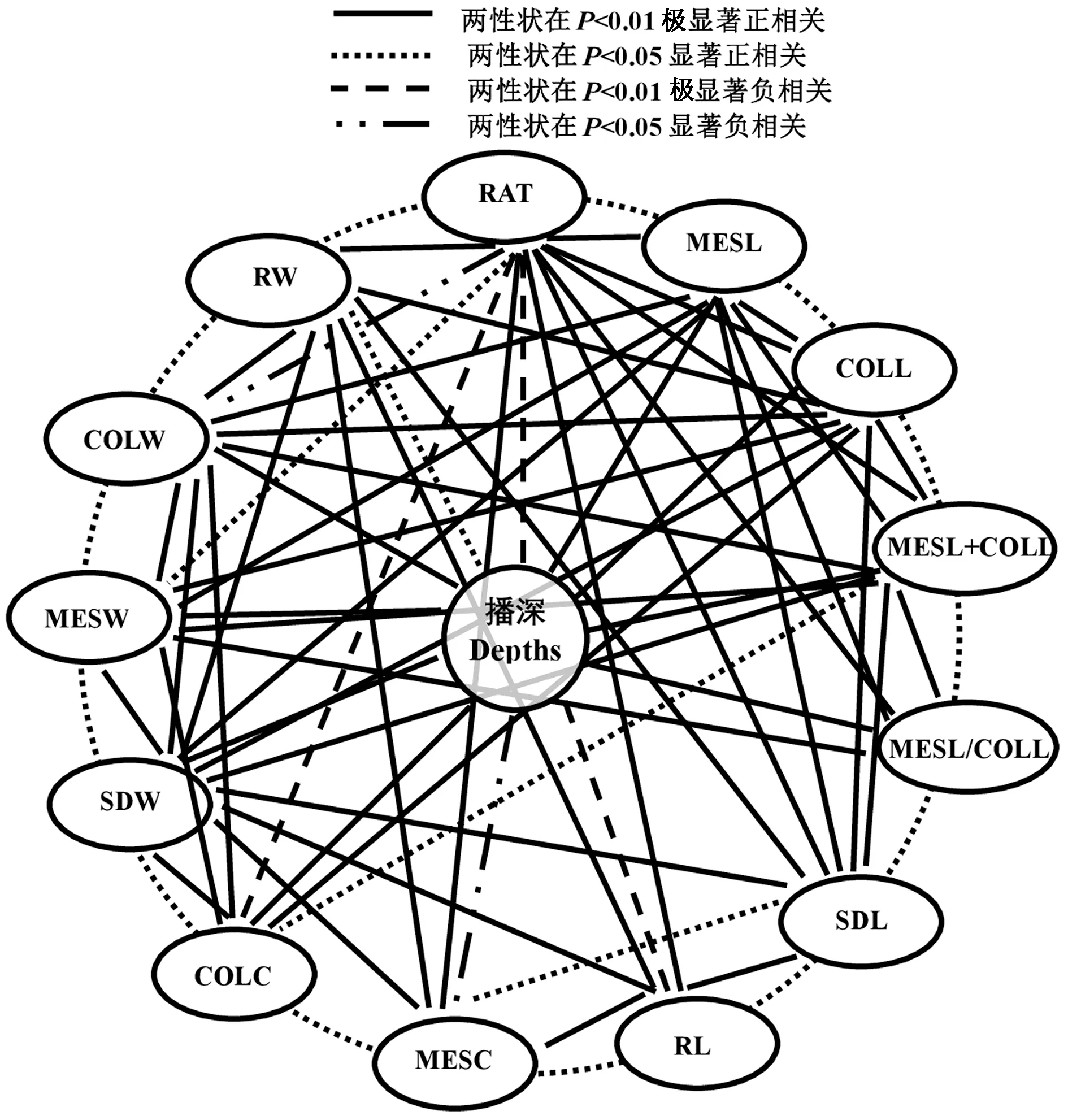
图1 玉米耐深播性状Pearson 分析Fig.1 Pearson correlation analysis of deep-seeding traits in maize
13 个性状各遗传方差分量分析表明(图4-A),除RAT 的母体方差比率及母体×环境互作方差比率不显著外,其余各遗传效应方差分量比率均在P<0.01/0.05 水平下显著,但所占比重不同。说明玉米这13个耐深播性状同时受多种遗传效应影响。其中MESL、COLL、MESL+COLL、MESL/COLL、RL 的加性遗传效应占主导地位,其加性方差比率均高于显性方差比率。此外,这些性状的遗传力分析表明(图4-B),所有性状的普通广义遗传力介于57.4%~85.9%,明显高于其互作广义遗传力;这些性状的普通狭义遗传力介于10.8%~35.7%,明显低于其普通广义遗传力;除RAT、SDW、MESW、COLW、RW 外,其余性状的互作狭义遗传力均低于其狭义遗传力。说明玉米13 个性状均显著受基因加性与非加性效应及其与环境互作效应调控。
2.6 玉米耐深播性状的配合力分析
13 个性状的GCA 和SCA 方差分析表明(图5),这13 个性状在F1组合间均达到差异极显著(P<0.01)。说明玉米这些耐深播性状在不同F1组合间的遗传差异真实存在。此外,所有性状的父/母本GCA及F1组合SCA 间均差异显著或极显著。表明玉米这些性状的GCA 和SCA 在亲本及F1间差异真实存在,可进一步预测亲本相关性状的加性效应及F1的显性效应。
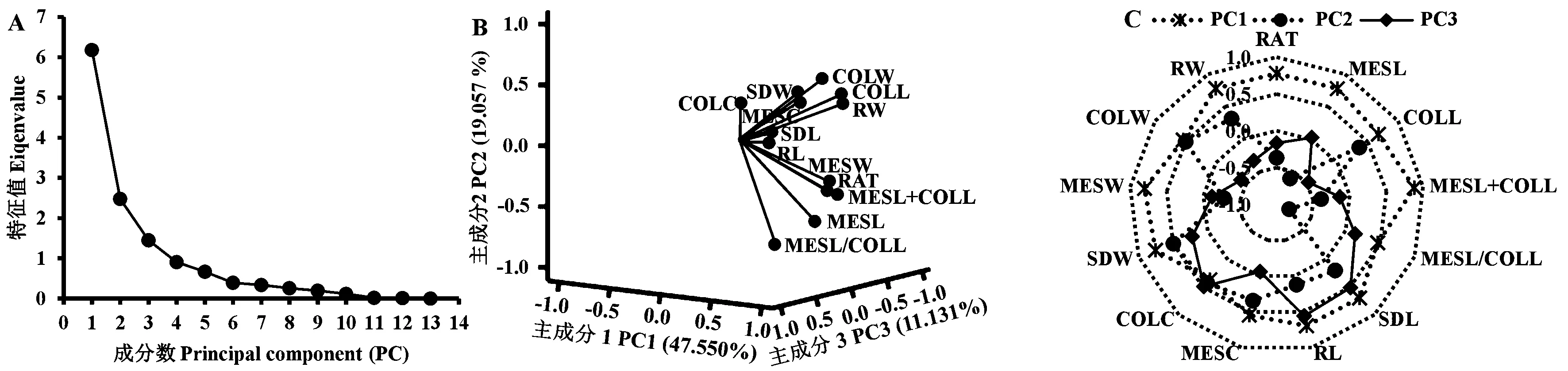
图2 玉米耐深播性状PCA 分析Fig.2 Principal component analysis (PCA) of deep-seeding traits in maize
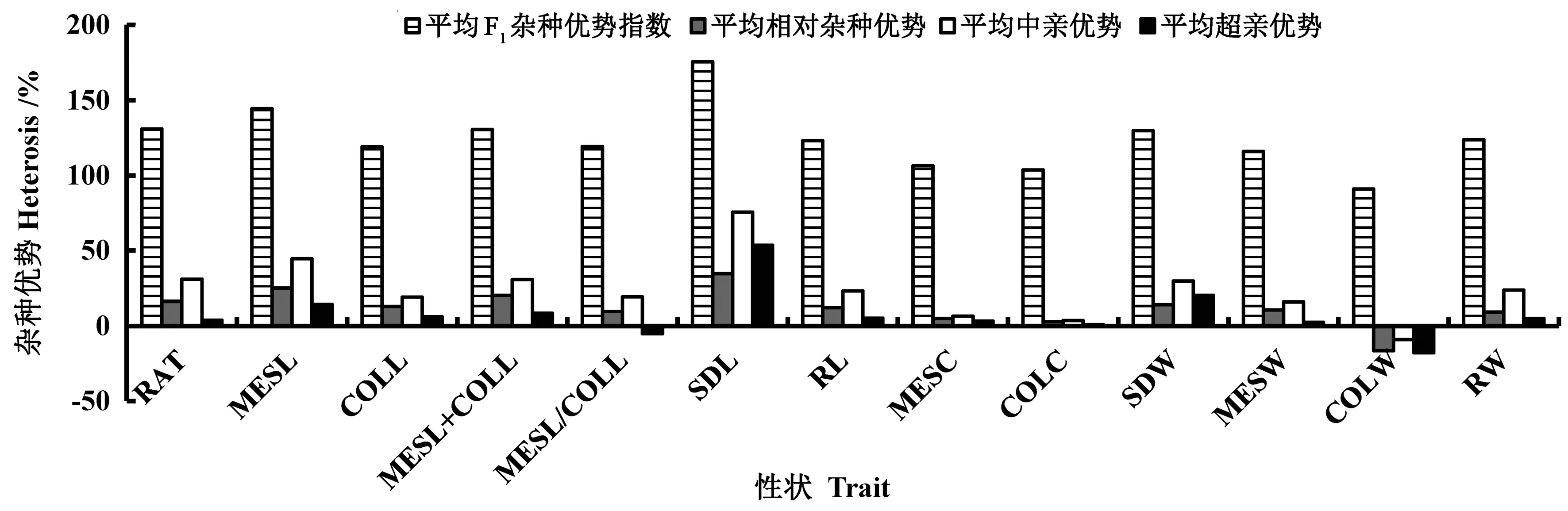
图3 玉米耐深播性状杂种优势分析Fig.3 Heterosis analysis of deep-seeding traits in maize
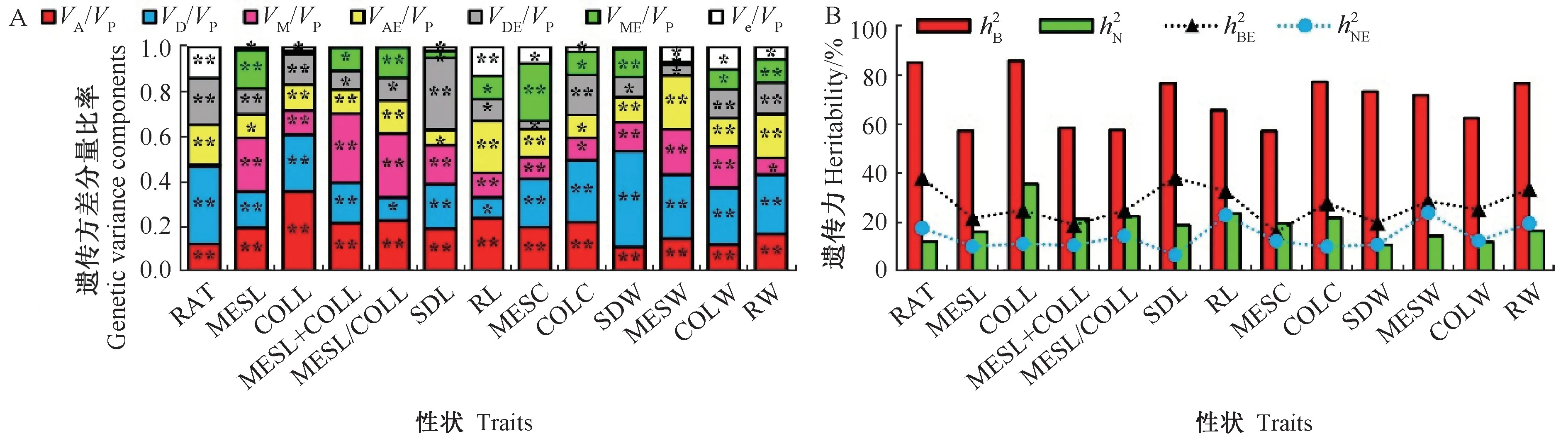
图4 玉米耐深播性状的遗传方差分量比率和遗传力估计值Fig.4 Estimates proportions of genetic variance components and heritability of deep-seeding traits in maize
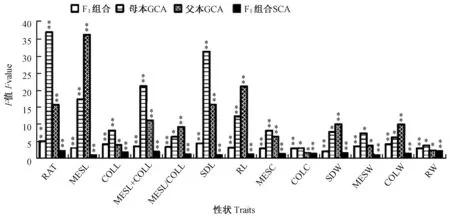
图5 玉米耐深播性状配合力方差分析(F 值)Fig.5 Variance analysis of combining ability of deep-seeding traits in maize (F value)
2.7 玉米亲本及F1 耐深播性状的加性和显性效应分析
由于61.5%的玉米耐深播性状的加性效应高于加性×环境互作效应(图4-A),因此可对22 份亲本的加性主效应进行分析。总体而言,22 份亲本13 个性状间的加性效应值差异较大(表3)。说明不同亲本耐深播性状的GCA 不同。其中8802A 的COLL、SDL、RL、MESC、SDW、MESW、COLW、RW 的加性效应表现均最高,其F1这8 个性状分别增加13.48%、1.08 cm、14.05 cm、0.26 mm、5.90 cm、0.74 g、0.13 g、0.13 g 和0.70 g。由于玉米这13 个性状均显著受显性效应及显性×环境互作效应的调控(图4-A),因此分析这些性状在特定F1中的显性效应十分必要。整体而言,22份F1这13 个性状的显性效应差异较大(表4),说明,特定环境下不同F1的SCA 不同。其中A413 ×K12RAT(26.51%)的正向显性效应最大,且MESL+CoLL、MESL/CoLL、SDL 和RL 呈显著的正向显性效应。A413 × A154RAT 的正向显性效应次之,为13.18%,且CoLL、MESL+COLL、CoLW 和RW 的正向显性效应显著。
3 讨论
深播下玉米种子顶土出苗过程往往涉及诸多复杂生理代谢反应,并通过不同组织的协同或拮抗作用,最终形成玉米的耐深播响应机制。彭云玲等[3,5]和Wilhelm 等[19]研究表明,中胚轴与胚芽鞘协同伸长是深播下玉米种子顶土出苗的关键因素;而赵小强等[7]及Zhao 等[20]研究表明,深播下各种植物激素能调控玉米中胚轴与胚芽鞘细胞的伸长与扩大或细胞数目的增减。本研究也得到一致的结果,即播深从3 cm 增加到15 或20 cm 时,玉米亲本和杂交种的中胚轴和胚芽鞘均显著伸长,且中胚轴(126.80%)伸长幅度远高于胚芽鞘(54.64%)(表1),致使玉米的中胚轴与胚芽鞘和(90.89%)、中胚轴与胚芽鞘比(57.22%)也协同升高;而玉米的中胚轴粗(7.56%)则略微减小,胚芽鞘粗(15.96%)略微增大。本研究还发现,不同播深对玉米幼苗、根系及其干物质积累量等也存在一定的影响,播深增加时玉米的苗长、根长及根重呈不同幅度的降低,而苗重、中胚轴重及胚芽鞘重呈不同幅度的增加。这与Sanusan 等[21]和李明月等[22]的研究结果一致。表明玉米的这些性状可能与其耐深播性存在着十分密切的关系。本研究在3 种播深下相关性状相关分析表明,13 个性状彼此间及其与播深间均呈显著正或负相关(图1)。进一步对3 种播深下相关性状的PCA 分析表明,前3 个主成分的特征值均大于1,累计贡献率达77.738%。因此,前3 个主成分代表了玉米耐深播特性的绝大部分信息,且中胚轴长、胚芽鞘长、中胚轴与胚芽鞘和、苗长、根长、胚芽鞘粗、中胚轴重、胚芽鞘重、苗重、根重、出苗率是造成不同玉米耐深播性差异的主要因素。因此这13 个性状可作物玉米耐深播性评价的主要参考指标,解析这些性状在后代中的杂种优势表现、遗传效应大小及配合力强弱,为耐深播优良种质遗传改良及耐深播品种选育奠定基础。
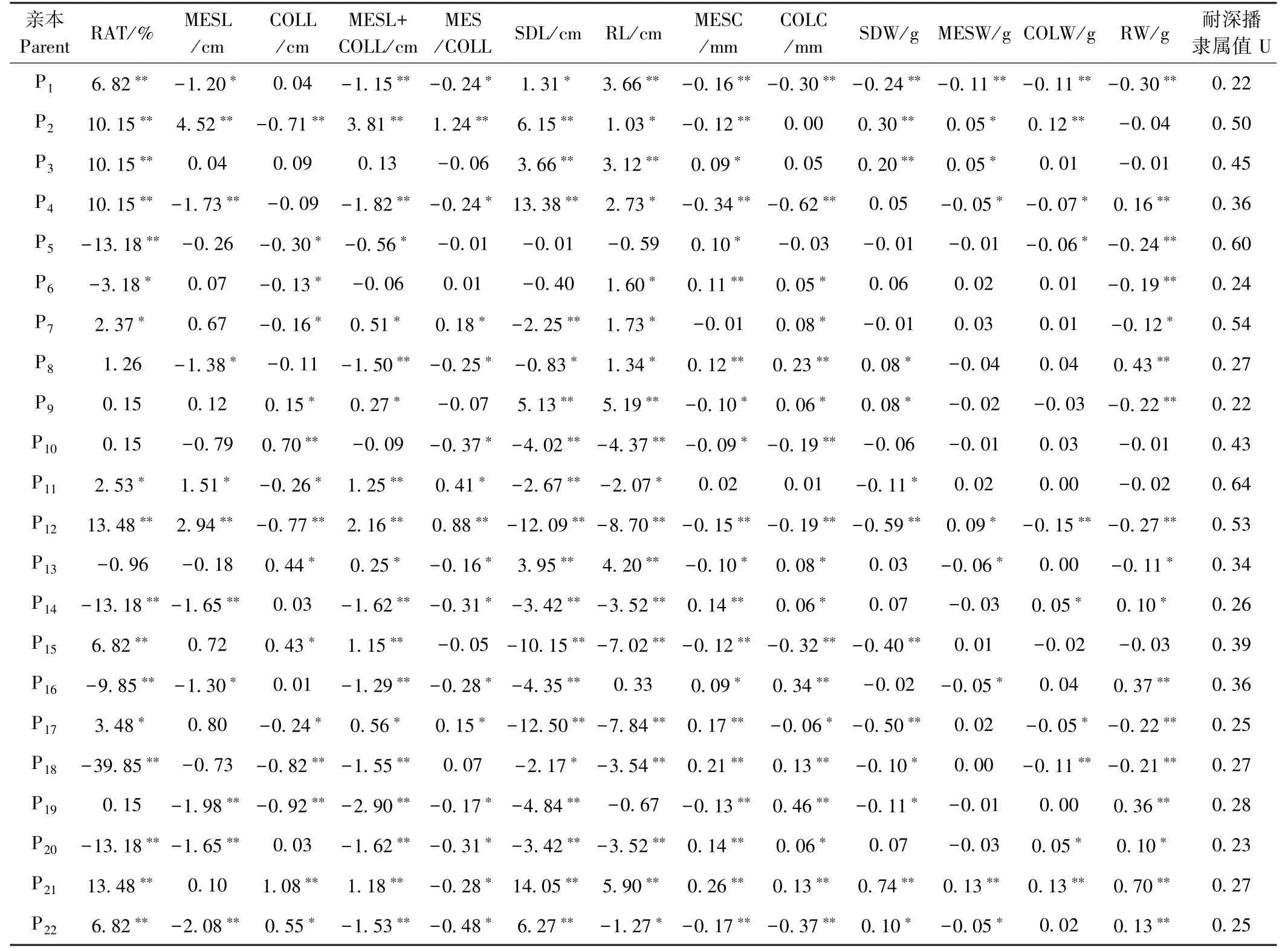
表3 玉米亲本耐深播性状加性效应及亲本耐深播性综合评价Table 3 Additive effects of deep-sowing traits in parents and comprehensive evaluation of deep-seeding tolerance in parents
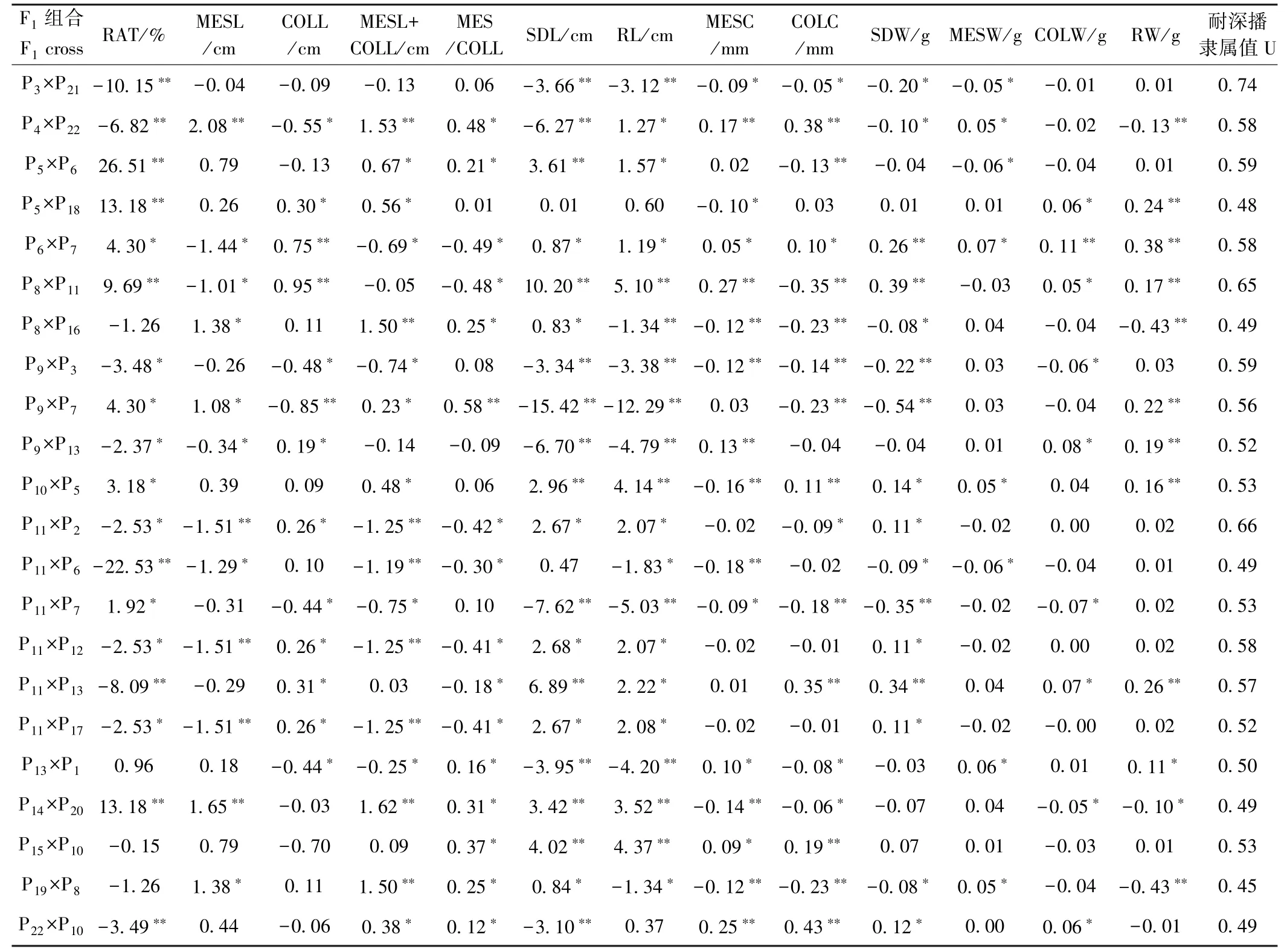
表4 玉米F1 耐深播性状显性效应及F1 耐深播性综合评价Table 4 Dominance effects of deep-sowing traits in F1 and comprehensive evaluation of deep-seeding tolerance in F1
作物性状间遗传性的差异造成其杂种优势表现的方向和程度也不尽相同[23]。因此明确玉米耐深播性状的杂种优势对优良耐深播抗旱新品种培育具有重要意义。王云生等[24]研究表明,玉米亲本整体耐深播性在其杂交种中的杂种优势指数平均为197%,杂交优势表现明显。Zhang 等[10]分析发现,玉米中胚轴长、胚芽鞘长、苗长及出苗率的平均相对杂种优势分别为115.74%、92.83%、92.05%和111.63%。而本研究表明,13 个性状的杂交优势均表现明显,平均F1杂种优势指数介于90.97%~175.64%;除胚芽鞘重的平均相对杂种优势为负值(-16.49%),其余性状的平均相对杂种优势均为正值;除F1的平均胚芽鞘重显著低于父母本、F1的平均中胚轴与胚芽鞘比低于高值亲本,F1的平均其余性状均显著高于双亲,这些性状均表现出较高的正向中亲和超亲优势。李月明等[22]研究也揭示,在3 种播深下玉米品种的平均出苗率均高于父母本,表现出明显的超亲优势,且这些玉米品种的出苗率还与其父母本呈显著或极显著正相关。说明玉米这13 个性状的F1杂种优势指数、相对杂种优势、中亲优势及超亲优势间的差异可能是由父母本的遗传不同引起的。因此培育综合耐深播性强的玉米新品种时,不仅需要注重对高亲、中亲及母本等的选择,还需兼顾杂种优势的影响,以提高育种选择效率。
作物性状的杂种优势是由不同遗传效应共同作用构成的基因网络系统形成的[25],因此系统解析玉米耐深播性状的遗传效应可为玉米耐深播杂种优势遗传机理的阐明奠定基础。Zhang 等[10]检测到了50.0%、33.3%、40.0%和50.0%调控中胚轴长、胚芽鞘长、苗长及出苗率的加性QTLs,检测到了33.4%、66.7%、60.0%和50.0%调控这些性状的部分显性QTLs,检测到了8.3%和8.3%调控中胚轴长的显性和超显性QTLs。而本研究采用ADM 模型对13 个耐深播性状的遗传效应分析表明,这13 个性状同时受加性、显性、母体、加性×环境互作、显性×环境互作及母体×环境互作效应的影响,这些效应在不同性状间所占比重不同。其中,中胚轴长、胚芽鞘长、中胚轴与胚芽鞘和、中胚轴与胚芽鞘比及根长的加性方差比率均高于显性方差比率。因此这5 个性状的加性遗传效应占主导地位,育种过程中可通过简单回交法或单交重组法在早代对其遗传改良。加性×环境互作方差比率、显性×环境互作方差比率在13 个性状间表现不同。说明环境互作效应对玉米这些性状的影响不同,今后应重点检测调控这些性状的QEIs(QTL-by-environment interactions)位点,遗传改良这些性状也最好在特定环境下进行,并充分发挥这些性状在特定环境下的杂种优势。研究还指出上位性互作效应是复杂数量性状遗传和杂种优势形成的重要遗传基础[26-27]。之前曹立勇等[11]已在水稻上定位到2 对调控中胚轴长的加性×加性上位性QTLs位点。推测调控耐深播性状的上位性QTLs 位点在玉米上可能普遍存在,今后应在更为复杂的遗传背景下有效地检测和利用玉米耐深播性状的上位性QTLs 位点,为玉米耐深播特性杂种优势利用提供有益参考。
亲本性状的加性效应值决定GCA 的大小,与性状的遗传力呈正比[28]。因此,亲本加性效应值可反映亲本的育种利用价值。而亲本的非加性效应值决定亲本组配F1的SCA,虽不能在杂交世代间稳定遗传,但可以指导杂种优势利用及杂交种培育[28]。本研究配合力分析发现,13 个性状的父/母本GCA 及杂交组合SCA 间均差异显著(图5)。表明玉米这些性状的GCA 和SCA 在亲本及F1间差异显著,因此预测亲本这些性状的加性效应及F1的显性效应十分必要。就亲本的加性主效应分析可知(表3),作为LRC 种质的8802A,除中胚轴与胚芽鞘比外,其余性状均表现较高的正向加性效应,但由于其耐深播性较弱,因此可利用Reid(Ca317、8723)和TSPT(A413、N192)中的强耐深播种质改良8802A。其次,作为TSPT 种质的昌7-2和H21 其分别有9 和11 个性状表现较高的正向加性效应,这2 个亲本还表现出较强的配合力、抗旱性和耐深播性[29],因此可利用这2 份材料作为耐深播种质改良及育种的主体亲本。进一步对22 份F1的13 个性状的显性效应进行分析(表4),结果表明A413×K12 出苗率的正向显性效应最大,为26.51%,其中胚轴与胚芽鞘和、中胚轴与胚芽鞘比、苗长及根长还表现出显著的正向显性效应和较强的耐深播性,预测其贡献主要来源于母本A413。A413×A154 出苗率的正向显性效应次之,为13.18%,此F1其余5 个性状的正向显性效应显著,而耐深播性较弱,可能受弱耐深播父本A154 的影响。JW183×综31 中胚轴长(2.08 cm)、中胚轴与胚芽鞘和(1.53 cm)、中胚轴与胚芽鞘比(0.48)、根长(1.27 cm)、中胚轴粗(0.17 mm)、胚芽鞘粗(0.38 mm)、中胚轴重(0.05 g)的正向显性效应显著,此F1的耐深播性显著强于双亲(表3 和4)。说明JW183×综31 的耐深播性状间GCA 较强。此外,H21×8802A 耐深播性最强,但其出苗率(-10.15%)、苗长(-3.66 cm)、根长(-3.12 cm)、中胚轴粗(-0.09 mm)、胚芽鞘粗(-0.05 mm)、苗重(-0.20 g)、中胚轴重(-0.05 g)均呈显著负向显性效应。因此今后在玉米耐深播新品种培育中可优先考虑综合加性效应优良的8802A 作父本,并可在H21×8802A特定组合后代中改良创制一些优良耐深播资源。
4 结论
本研究表明,深播下玉米出苗状况、中胚轴、胚芽鞘、幼苗及根的生长形态与有机物积累通过彼此间协同或拮抗作用将玉米幼芽送出地表;13 个耐深播性状的杂交优势均表现明显,F1杂种优势指数均大于90%,除胚芽鞘重、中胚轴与胚芽鞘比外,其余性状均呈不同程度的正向中亲和超亲优势;中胚轴长、胚芽鞘长、中胚轴与胚芽鞘和、中胚轴与胚芽鞘比及根长的加性遗传效应占主导地位;其余性状同时受加性与显性遗传主效应及其与环境互作效应的影响,13 个耐深播性状间的双亲GCA 及F1的SCA 均差异显著,本研究为更系统地揭示这些性状杂种优势形成的遗传基础及改良创制耐深播资源提供了参考。
