元江干热河谷低海拔地带退化生态系统植物多样性及物种分布模式
2021-02-16杨逢春刘景欣黄华平构箭勇文慧婷KRITANAPrueksakornHONGAnhThiNguyenCHAYASarathchandra
杨逢春,刘景欣,黄华平,构箭勇,文慧婷,李 叶,陈 娴,KRITANA Prueksakorn,HONG Anh Thi Nguyen,CHAYA Sarathchandra
(1宜宾职业技术学院,四川宜宾 644003;2中国科学院西双版纳热带植物园环境教育中心,云南勐腊 666302;3中国热带农业科学院环境与植物保护研究所,海口 570100;4云南省红河哈尼族彝族自治州气象局,云南蒙自 661100;5宋卡王子大学国际教育创新,社会参与和可持续发展中心,泰国普吉 83000;6中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085;7斯里兰卡拉贾拉塔大学应用科学学院生物科学系,斯里兰卡密亨塔勒 50300)
0 引言
全球变化给地球生物及其生活环境都带来了深远的影响。物种生长、分布、以及物种与他们所赖以生存的空间的相互联系亦受到全球变化的影响[1]。与此同时,这些生态系统也以各种方式,如自我重建来抵御或适应这些影响[2]。占陆地生态系统40%的干旱和半干旱生态系统在全球变化背景下发生了深刻的变化[3]。在干旱生态系统中,物种与环境之间的相互适应可以通过形态变化或生理改变等途径,而这些变化或改变受到具体的环境因子的影响。由于环境因子存在具体的空间限制,因此,植物的变化和适应亦受限于这些空间限制。为了研究干热河谷的退化生态系统的环境特征,本研究选取了河谷低海拔地段的植物种类及其组合作为媒介开展研究,拟通过分析各种植物种类及其组合在干热河谷内的分布差异,来探究该干热河谷内的生态环境差异。环境条件对河岸植物的影响主要为环境因子的不稳定性[4]。然而目前,关于植物对各类环境因子的耐受性亦即生态幅还没有一个全面可靠的数据库可供参考,无法准确定义某一物种或某一类类群的抗性区间或耐受度,只能根据自然地理分布状况给出大致的范围,属于经验值,这也是研究受限的地方。
1 材料与方法
1.1 研究区概况
元江干热河谷是元江-红河中上游的一个典型干热河谷,位于中国西南部,横跨亚热带亚热带,全年高温干燥。自17世纪以来,该地区的植被与植物种类组成经历剧烈的变化[5]。元、明时期当地还是茂密的森林植被,随着人类开发强度加大而逐步退化,到17世纪中叶,它就已经完全转变为热带山地萨王纳植被,以稀疏乔木和高大草本为主要特征[6]。旱生植物覆盖地表,低矮灌木、丛生型草本、多肉多浆和一年生植物为主要优势物种。岩石风化严重、破碎化程度较高,土壤主要为紫色砂壤土以及页岩风化形成的燥红土,土层薄,保水保肥能力低[7-8]。
1.2 研究方法
1.2.1 样带及样方设计 研究地点位于元江干热河谷核心区,全长105.26 km,设计2种类型的样带(图1)。一种为垂直于河流的样带(P-样带),从河边起始点开始向外延伸,每条样带长70 m,样带总数为6条,标记为A-F。在P-样带上,从河漫滩向河岸坡地延伸,每隔20 m设置一个样方,样方尺寸为10 m×10 m。每条样带上有4个样方,共24个样方。另一种样带为平行于河道的理论样带(L-样带),它依托垂直样带A-F的样方1-4,从上游向下游延伸。因此每条L-样带理论长度为105.26 km,样带间距20 m,共4条样带,每条L-样带从上游到下游共有6个样方,共24个样方。
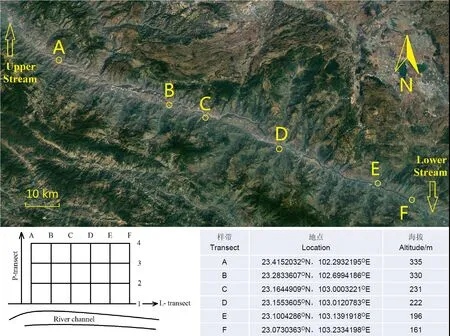
图1 研究地点及样带设计
样方内物种鉴定以中国植物志英文版(http://www.iplant.cn)为准。
1.2.2 研究区域的气候概况 本研究前期整理了1968—2018年元江干热河谷内4个低海拔气象台站(312~576 m)的气象监测数据表明,元江干热河谷的年均温、降水量和蒸发量分别为23.95℃,814 mm和205 mm(真值)。
1.2.3 生物多样性计算 生物多样性的计算包括物种多样性α和生境多样性β两个方面[9]。α多样性采用以下指标:Shannon-Wiener指数H;均匀性指数J;以及显著性指数Dominance。β多样性采用Bray-Curtis,它是一个相似性的指标,值越大,表明两个样带或样方之间的生境差异越小。
1.2.4 物种期望值、稀有指数和丰富度指数计算 物种期望值Species_estimated:在t个样方内的物种数量(累计值)。
物种稀有指数包括Singletons和Uniques[10-11]。其中,Singletons指征在t个样方内只有一个植株的物种数量(累计值),植株数量唯一,但生境不是唯一的;Uniques指征仅出现在一个样方中的物种数量(累计值),植株数量不唯一,但生境是唯一的。
丰富度指数选用ACE、ICE、和Chao2[10-11]。其中,ACE为基于物种丰度的覆盖度指数(均值);ICE为基于物种发生率的覆盖度指数(均值);而Chao2是由Singletons修正的物种丰富度指数。三者可相互验证。
所有样方水平和样带水平上的数据矩阵全部加载到EstimateS(Version 9.1)进行999次计算。然后使用R程序中lme4程序包生成简单线性模型GLMM[12-13],使用Vegan程序包进行多变量的分析[14],最后使用labdsv程序包开展基于物种个体数和出现频率的指示种分析[14]。所有模型均经由凯方检验(χ2,P<0.05),残差由Shapiro-Wilk检验进行正态性分析(P>0.05)[15]。
1.2.5 指示种分析 采用R程序里面的indicspecies程序包指示种函数IndVal进行多级模式分析后筛选特定指示种[16-17],显著性水平控制在P<0.05。
2 结果与分析
此次研究中,共发现56科107种植物共15013株,其中外来植物11科15种5496株。乔木12科17种973株,灌木有11科19种1259株,草本28科65种12650株,另有6科7种131株属于攀援型藤本。无论物种数量还是个体数量上看,草本植物都是优势类型。
2.1 物种多样性α相关指数在P-样带和L-样带的变化
在2种样带上H指数均呈递减的态势,属于负相关(图2)。Hmax在L-样带上的差异达到显著性水平水平(P=0.02136),说明从河岸边往外延伸,最大多样性是显著减小的。根据P-样带中样方的实际物种数进行线性回归,其结果也呈现类似的变化趋势(P=0.04014)(图3)。
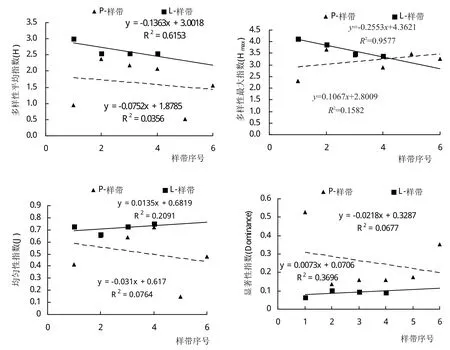
图2 多样性平均指数(H)、多样性最大指数(Hmax)、均匀性指数J和显著性指数Dominance在不同样带间的差异
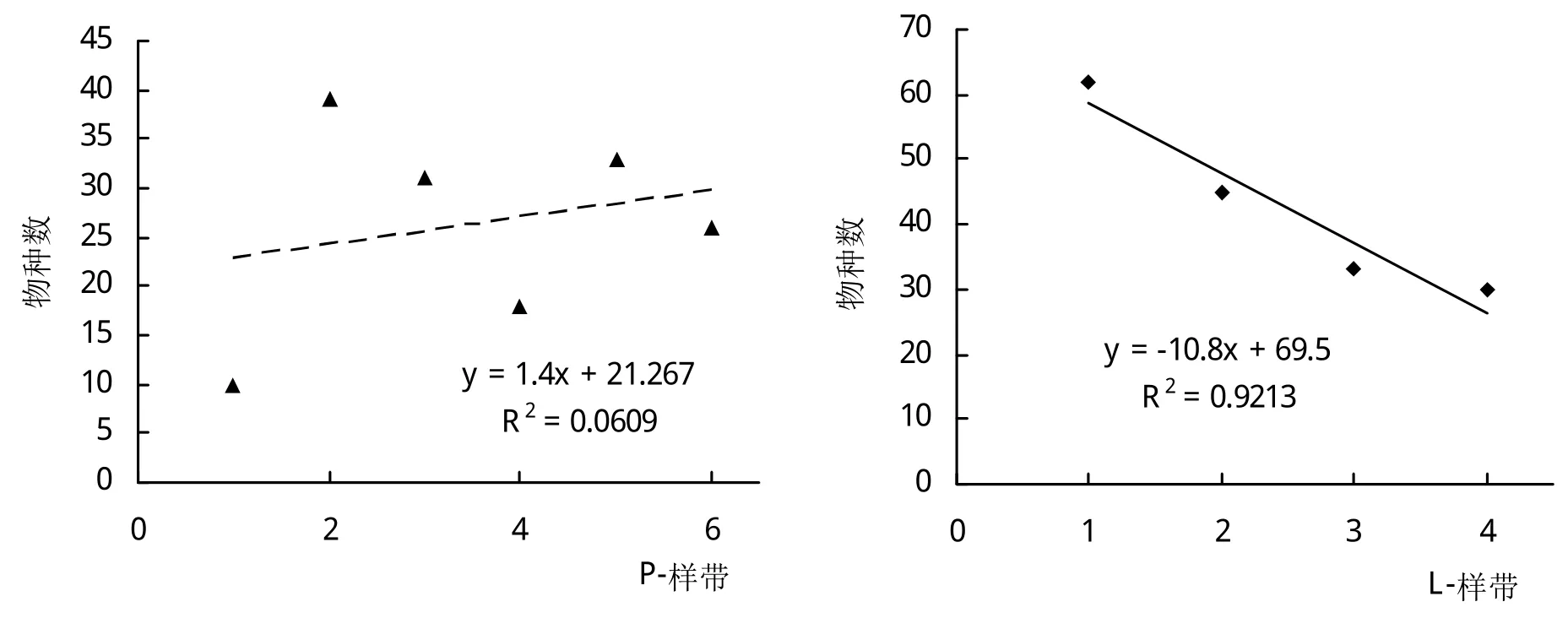
图3 实际物种数量在垂直河流P-样带和沿河平行L-样带的变化趋势
均匀性指数J在样带之间变化很大。尤其是在P-样带上,最大值与最小值之间存在较大差距,体现了上下游之间的生境差异,尤其是在下游,随着河流水量增加、河岸基质及附着植被稳定,均匀性可能会逐渐减小。
显著性指数Dominance极值分化明显,沿河流方向,上游样带的指数要高于下游样带(图2)。而在平行于河道方向上,指数呈逐渐增加的趋势(图2),说明随着样带远离河道,一些适应缺水环境的物种逐渐占据优势;而在靠近河道的样带中,显著性指数普遍不高,其原因可能是不定期的河水泛滥可将尚未长成的植株冲走(图4,A)。

图4 元江干热河谷河漫滩地带的植被特征
2.2 β多样性和生境相似性指数Bray-Curtis的变化
P-样带中Bray-Curtis指数均较高,最高达0.998,亦即2个样方的生境几乎一致(图5);而L-样带中Bray-Curtis整体较低(图5)。此外,Bray-Curtis的最大值通常出现在距离河道较远的样带和样方中,说明距离河道越远,生境趋同性越高。
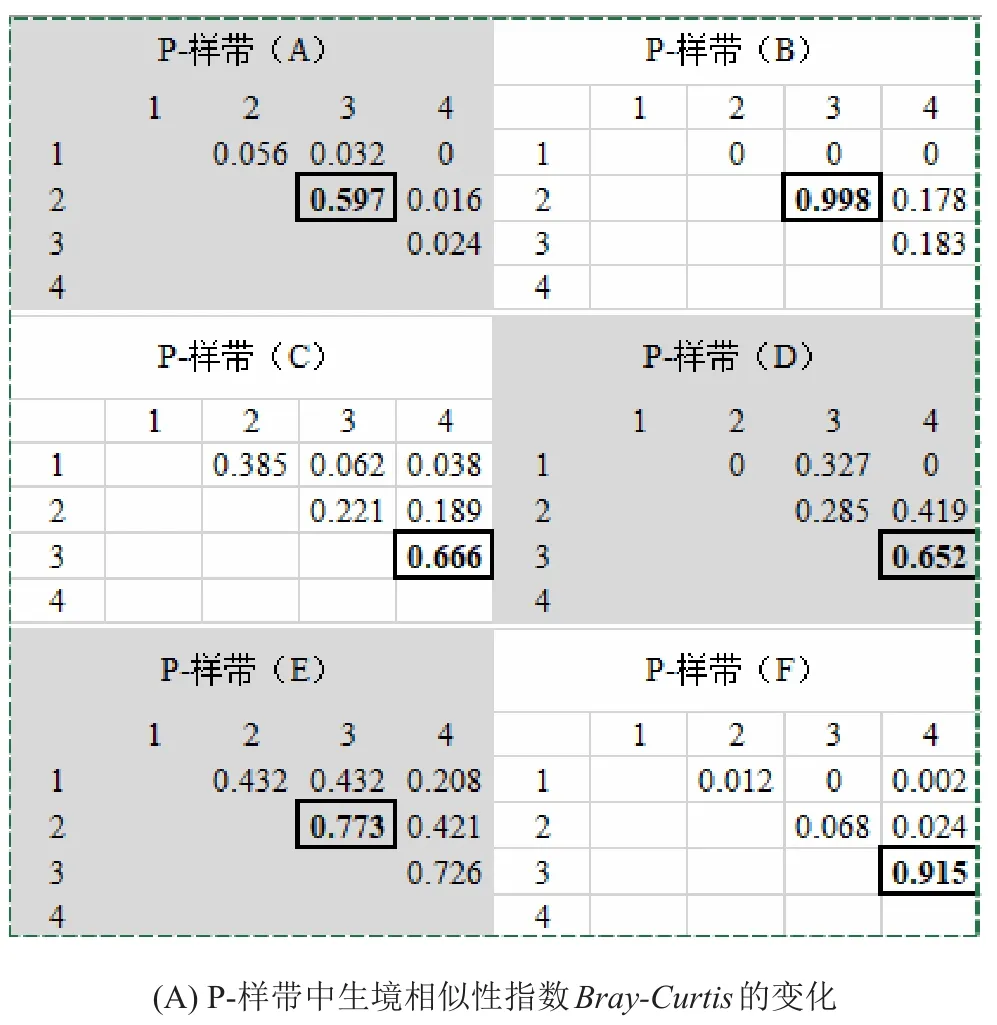
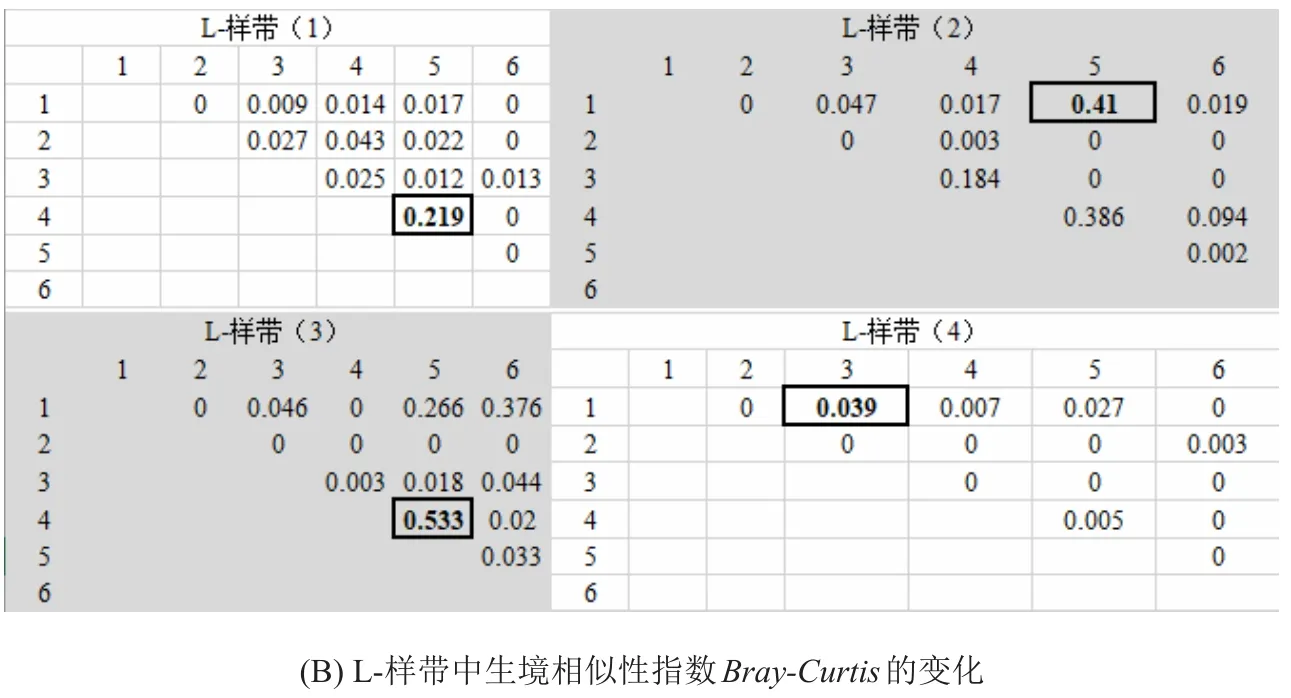
图5 不同类型样带上的生境相似性系数
2.3 物种期望值、稀有指数和丰富度指数在P-样带和L-样带的变化
P-样带中,除了样带A和D以外,物种期望值Species_estimated是一种连续上升的态势(图6),物种数随样带延伸不断增加,说明在70 m的P-样带上,生境是不断变化的,这种变化与河道亦即水源的距离有关。靠近河道的样方常以耐水淹的植物为主,如积雪草Centellaasiatica(L.)Urban,野 芋Colocasia antiquorumSchott,过江藤Phyla nodiflora(L.)E.L.Greene等,以及芦苇Phragmites australis(Cav.)Trin.ex Steud.的幼苗。但在远离河道的样方,则代之以中生型或者耐旱型的植物,如大叶山蚂蝗Desmodiumgangeticum(L.)DC.,大叶千斤拔Flemingia prostrataC.Y.Wu,清香木Pistacia weinmanniifoliaJ.Poisson ex Franchetgigantean等。
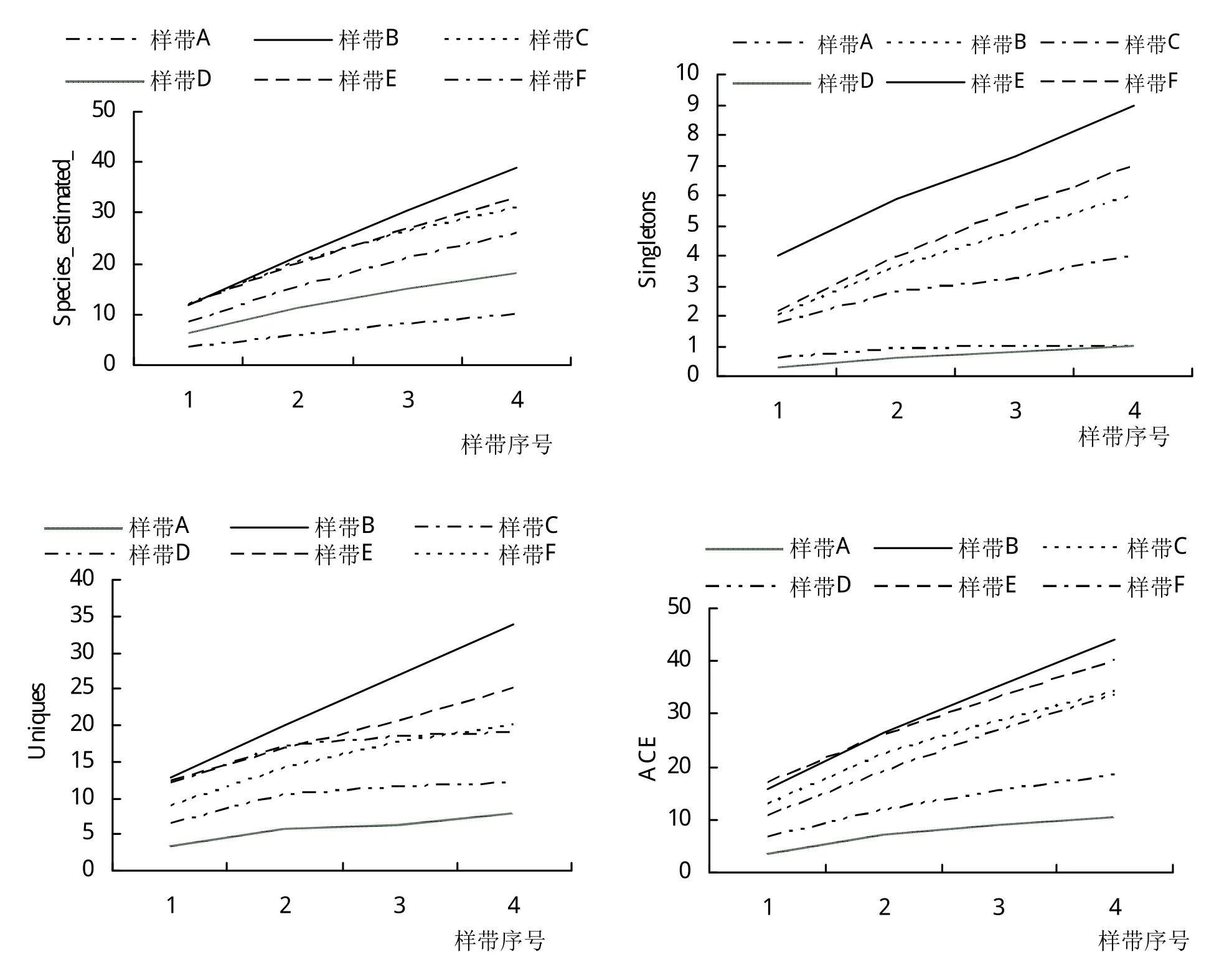
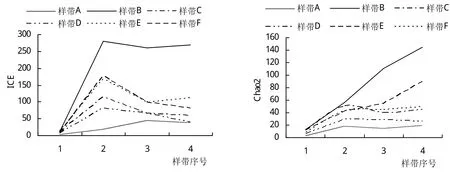
图6 垂直河流P-样带中物种期望值、稀有性和丰富度指数的变化
L-样带中,从上游到下游,物种期望值Species_estimated呈连续上升的态势(图7)。上游样方中基本被草本和灌木所占据,常见物种有荩草Arthraxon hispidus(Trin.)Makino,甜根子草Saccharum spontaneumL.,类 芦Neyraudia reynaudiana(kunth.)Keng等。而在下游样方中,优势物种则以乔木和灌木为主,如构树Broussonetia papyrifera(Linnaeus)L'Heritier ex Ventenat,大叶山蚂蝗,瘤腺叶下珠Phyllanthus myrtifolius(Wight)Muell.Arg.(外来种)等。
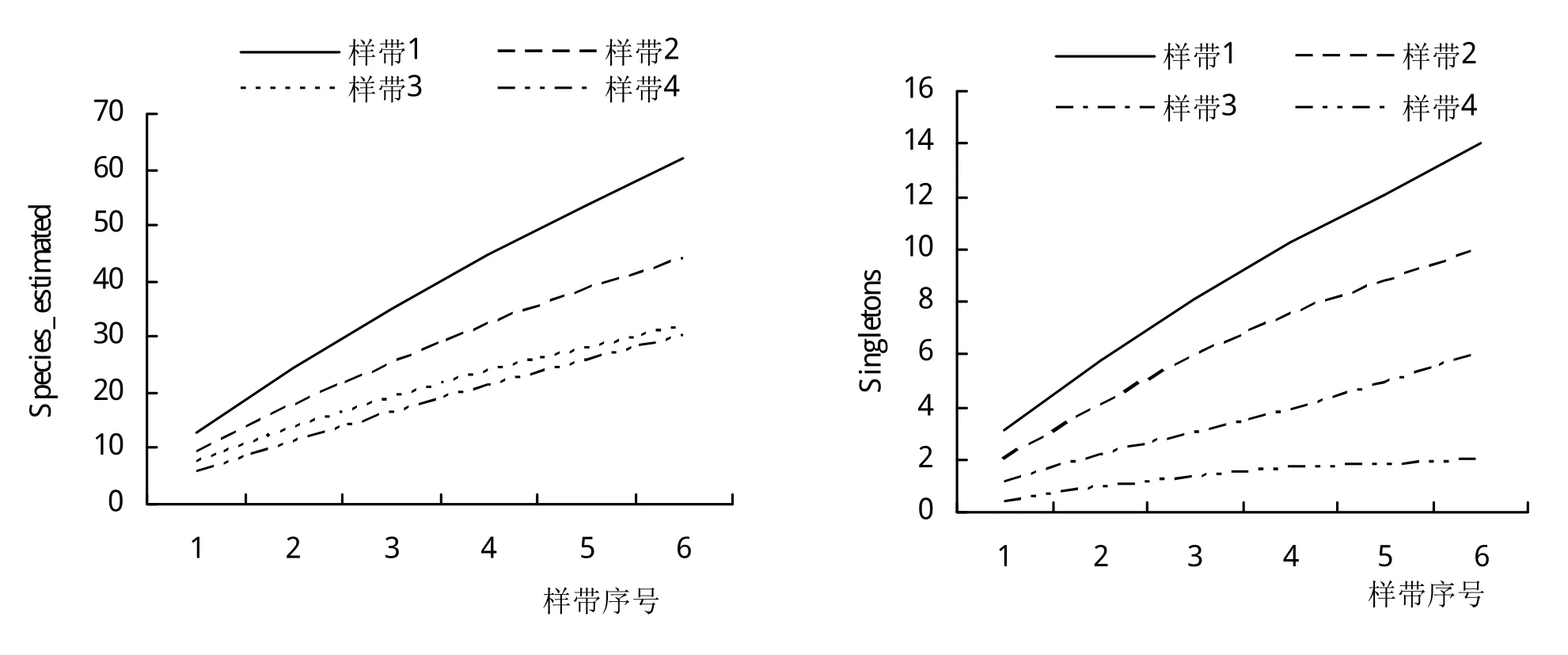
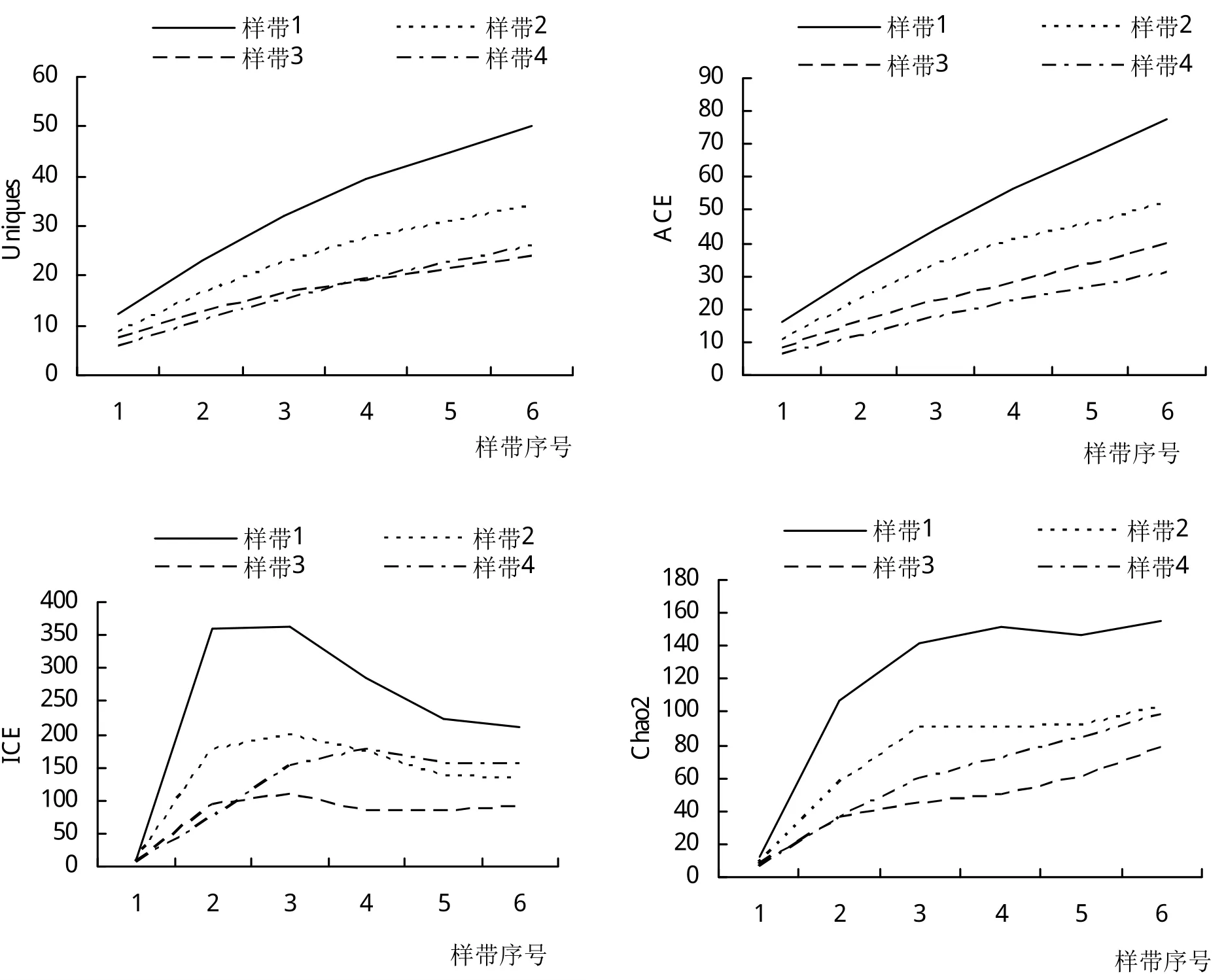
图7 沿河平行L-样带物种期待指数、稀有指数和丰富度指数变化
Singletons指数在P-样带中呈稳定增长态势,但样带A和D例外,仅有1个物种以单个个体的形式存在,分别为苦荬菜Ixeris polycephalaCass.和藠头Allium chinenseG.Don。下游样带E、F的Singletons指数要高于其他样带。而在L-样带中,Singletons最大值出现在距河道最近的样带。
Uniques指数在P-样带中的B、E和F中快速上升,但是其在样带A、C和D中Uniques指数偏低,且过早地出现转折点,说明这3条样带中的样方过早地出现了生境同质化的现象。L-样带中Uniques指数在所有4条样带里面均呈现快速增长态势,说明越往下游,生境异质性越高。
在P-样带和L-样带中,ACE指数与ICE和Chao2均差异太大,不能相互验证,因此不予采纳。P-样带中,所有丰富度指数的曲线在样方2出现转折。此处距离河道25 m,是河漫滩与河岸坡地之间的转折区域,植株的根部供水状况收到影响。L-样带中,河道附近样带的物种丰富度指数远远大于远离河道的样带。此结果可与P-样带的结果相互验证。
2.4 指示种分析
共6个种被筛选出来作为候选的生境指示植物(P<0.05),分别为银合欢(P=0.002**,外来种,世界广布),芦苇(P=0.015*,本地种,世界广布),毛果茄(P=0.019*,本地种,区域性分布),地毯草(P=0.019*,外来种,世界广布),长柄山蚂蝗Hylodesmum podocarpum(P=0.018*,本地种,世界广布),和藿香蓟(P=0.011*,外来种,世界广布)。世界广布种对于本地生境没有指示意义,因此毛果茄(图4,B)是唯一区域性分布的物种。环境选择性上,它是一种典型的河滩沙地植物,生长海拔100~1300 m,严格限定在干热河谷高温干旱的气候区间。因此它可以作为元江干热河谷的生境指示种。
3 讨论
3.1 河谷内水分状况与植物的分布
在P-样带中,物种期望值、稀有指数和丰富度指数的变化揭示出水分供给和植物之间的相互关系。首先,物种期望值指数已明显指示远离河道的位置可发现更多不同的植物种类。其次,稀有指数Singletons和Uniques的增长变化与距离河道的远近有关。物种丰富度指数ICE和Chao2的变化显示距离河道越近,物种数越多。因此,在元江干热河谷中,物种的分布与河道水源补给强烈相关,受到元江水流大小的影响。这与其他研究的结果是相似的[18]。
所有受调查的样带都位于河谷内的低海拔位置,空气干燥。因此,在空气补水无法实现的情况下,植物的生长只能依赖于根部吸水[19]。干热河谷内附生植物严重缺乏。水分补给决定着植物的生长和分布,是所有环境因子中的决定性因子[20-21]。与此同时,物种丰富度被认为是环境空气湿度的反映[21-22]或被看作是环境湿度与凉爽气温的最佳指标[23]。然而,在干热河谷环境下,空气湿度视为可忽略指标,植物的生长与根部地下水供给紧密相关。河漫滩植物被认为是减弱河水冲刷作用并保护河岸[24],但是我们的调查结果显示元江-红河水流较急,且季节性洪水明显,河漫滩植物的生长明显受限,漫滩植被无法向更高阶段发展。因此,物种期望值Species_estimated和物种丰富度指数ICE和Chao2在靠近河岸的样带和样方中偏高,但是植株数量很少,仅有少数物种种群能够发展壮大,如牛筋草Eleusine indica(L.)Gaertn.,银 胶 菊Parthenium hysterophorusL.(外来种)和香附子Cyperus rotundusL.。这些种的成功完全归功于它们发达的根系,在洪水来临时可以将植株牢牢固定住。
3.2 河谷内的生境异质性
植物的生长和分布与其生活环境的环境因子紧密关联。生境异质性理论认为不同植物对环境的需求差异主要体现在生长基质、水分、温度、养分等各个方面[25]。在长期的演化与适应过程中,植物种类与生境因子之间的关系相对固定下来,特定的植物或植物组合可以反映特定的环境[26-28]。复杂和多样化的生境可以支持更多的植物种类,而单一的环境支持的物种数要少得多[29]。而此研究的结果证明,在元江干热河谷低海拔的地区,整体环境属于高温干燥的环境,但仍有小生境存在。对于植物而言,决定生存的主要因子就是水分的课获得性,因此水因子是元江干热河谷小生境的主要决定因子。根据多样性指数α、稀有指数Singletons和Uniques分别在L-样带和P-样带中的表现可知,随着河流向下游延伸,以及随着样方靠近河岸水源地,样方及样带的生境异质性都呈逐步增强的趋势。结合Bray-Curtis指数,其指征距离河道越远,生境趋同性越高,还是说明水分在生境中的决定左右。而在该区域的主要水源补给来自河道,随着样带样方远离河道,河水补给急剧减少,导致了生境趋同化。但是需要注意的是,随着干扰的加剧,这些小生境随时可能会消失。
3.3 指示种分析
指示种与环境因子的关系紧密,指示种种群的发生、扩大、缩小、更新等过程反映了环境因子的动态变化。由于不同的植物指示种适应于不同的生境以及环境因子,有的种还具有严格的生境限制,对环境具有严格的选择性,对环境因子的变化敏感,因此它们是指示当地环境演变过程的首选[30]。毛果茄作为一种区域性分布的本土植物,仅分布于阿富汗、印度次大陆和华南地区,具有典型的植物表型特征和环境选择特征。在元江干热河谷中,黄果茄的生境严格限制在中上游河段低海拔地区形态特征上,具有多刺、表皮蜡质、茎木质化或半木质化以及根系发达等耐旱植物的典型特征,而这些特征反映了该物种对长期燥热干旱气候的生态适应过程。因此,选择毛果茄作为元江干热河谷的环境只是植物是比较合适的。
4 结论与建议
元江干热河谷低海拔地区水热分布特殊,尽管靠近河流,但是空气干燥,气温高,生境单一,植被发育不良、生多样性低、物种稀少。水源供给是影响元江干热河谷里面生物多样性和物种分布的决定性因子。沿河道周边的特异性小生境是植物的避难所,这些小生境为干热河谷里面的生物多样性做出了贡献。但是随着全球变化的加剧和人类影响的扩大化,这些小生境的存在岌岌可危。我们可以利用毛果茄作为元江干热河谷生境变化的指示种,当然这需要更多深入细致的研究。关于干热河谷内主要环境因子如水分、温度的长期变化规律及其驱动机制也需要更多深入细致的分析。
