10个玉米ZmbZIP基因表达模式分析
2021-02-12贾利强吴洪梅曾科美龙金云赵秋芳
贾利强,吴洪梅,曾科美,龙金云,郑 飞,赵秋芳,陈 曙
(1. 贵州师范学院, 贵州 贵阳 550018;2. 中国热带农业科学院, 广东 湛江 524091)
0 引言
【研究意义】玉米作为重要的粮食和经济作用,在保障粮食安全和种植者创收增效中发挥着重要作用。玉米生长周期内,经常遭受各种逆境胁迫因子的侵害,致使产量下降,经济收入遭受损失,培养高产稳产抗逆性好的玉米新品种至关重要。bZIP蛋白作为转录因子中种类最多、分布最广的调控因子之一,其功能在越来越多的作物中进行研究,bZIP广泛参与植物生长发育进程及逆境胁迫响应途径。
基于此,以玉米自交系郑58为试验材料,人为模拟盐、干旱、低温和氮缺乏胁迫,系统研究bZIP基因的表达模式,可以为深入挖掘这些基因的功能提供有价值的科学数据。【前人研究进展】bZIP转录因子广泛分布于真核生物中,包括酵母、动植物等,并在其生长发育进程中发挥着重要的作用。bZIP转录因子结构特征是含有bZIP结构域,包含N端保守的碱性蛋白基序和C端保守的亮氨酸拉链基序,前者功能为核定位信号和靶基因的标定,而后者的功能为二聚体化,激活下游基因的表达[1]。随着基因组测序计划的开展,越来越多的植物bZIP基因家族成员被报道出来。 拟南芥[2]编码78个,葡萄[3]有55个,大枣[4]有45个,禾本科作物水稻[5]、玉米[6]、高粱[7]和荞麦[8]分别有89个、125个、92个和96个,油料作物油菜[9]、大豆[10]和芝麻[11]各含有247个、131个和63个,茄科作物番茄[12]、马铃薯[13]和辣椒[14]分别有69个、80个和60个。根据序列相似性、保守蛋白基序和蛋白结构等差异,拟南芥bZIP蛋白可以划分为10个家族[1],而在水稻中,可以划分为11个亚家族[5]。bZIP基因广泛参与植物生长发育进程及逆境胁迫响应途径[15],诸如生物或非生物胁迫、种子萌发,开花和种子发育、激素和糖信号转导等[16-18]。FD蛋白,bZIP家族成员之一,调控植物开花[19],而另一个成员,HY5蛋白抑制茎的伸长和侧根发育[20]。越来越多的研究表明bZIP基因在植物抵御逆境胁迫进程中也发挥着重要作用,包括水分胁迫[21]、盐毒害[22]、温度胁迫[23]、激素信号转导[23-25]、病虫害[26-27]等。拟南芥AtbZIP1、AtABF3、AtbZIP24等基因参与调控拟南芥应答温度胁迫、盐胁迫和干旱胁迫[28-29]。水稻中的OsbZIP23、OsbZIP71和OsbZIP52也参与调控各类逆境胁迫响应途径,超表达OsbZIP23和OsbZIP71能显著提高水稻的抗旱和抗盐性能[3],而OsbZIP52则负调控水稻抵御抗旱和抗冷性能[28]。大豆GmbZIP1、GmbZIP44、GmbZIP62和GmbZIP78也参与调控各类逆境胁迫响应途径,超表达这些基因会提高大豆的抗逆性能[30-31]。拟南芥bZIP亚家族A、D、H和S等成员广泛参与调控N吸收效率,其中AtbZIP1是传递N信号途径的主要调控因子[32],AtHY5和AtHYH等基因调控硝态氮、铵态氮转运体以及硝酸盐和亚硝酸盐还原酶活性[20,33],小麦TabZIP60负调控氮素吸收和小麦产量[34]。【本研究切入点】玉米自交系郑58作为我国重要的骨干核心种质资源,在各类逆境胁迫处理下,包括高盐、干旱、低温和硝态氮或铵态氮缺乏胁迫下,ZmbZIP响应表达模式还未见报道。【拟解决的关键问题】本文利用人为模拟各类逆境胁迫,设置200 mmol·L-1的NaCl溶液、20%PEG6000、4 ℃低温和硝态氮或铵态氮缺乏胁迫试验,选取10个ZmbZIP基因,探测这些基因的应答表达模式,以期为进一步深入研究玉米bZIP基因生物学功能提供科学数据。
1 材料与方法
1.1 试验材料
本研究所用到的试验材料为玉米核心自交系郑58,保存于中国热带农业科学院玉米研究中心。玉米自交系郑58授粉扬花期,采集根、茎、叶、花和授粉15 d的嫩穗,快速液氮处理,以备后用。
1.2 试验方法
1.2.1 玉米ZmbZIP的生物信息学分析 本文研究中所涉及的基因、蛋白和CDS序列来自于Phytozome v9.1数据库(http://www.phytozome.net/)、MaizeGDB(Maize Genetics and Genomics Database)数 据 库(http://www.maizedb.org/)和TAIR (https://www.arabi dopsis.org/)。利用在线软件ExPasy来分析玉米ZmbZIP蛋白的分子量大小和理论等电点(http://web.exp asy.org/cgi-bin/prot-param/protparam)。依据ZmbZIP基因的物理位置,利用Mapinspect绘制其在染色体上的分布(http://www.plantbreeding.wur.nl/uk/software-ma pinspect.html)。根据ZmbZIP基因和CDS序列,利用在线软件GSDS 2.0(http://gsds.cbi.pku.edu.cn)来分析ZmbZIP基因结构。本文中有10个ZmbZIPs和78个AtbZIPs蛋白序列构成的数据矩阵,利用在线软件MAFFT进行多重比对分析,剔除非保守蛋白序列,构成保守的bZIP蛋白序列数据矩阵,用于进化树分析。利用MEGA X软件中的NJ算法来分析bZIP蛋白之间的进化关系,缺失数据参数选Pairwise Deletion,模型为P-distance,校验参数Bootstrap重复次数为1000次。
1.2.2 非生物胁迫试验 郑58自交系种子经5%次氯酸钠溶液表面消毒,28 ℃黑暗催芽,生长于Holland营养液中,生长条件为28 ℃光照16 h,25 ℃黑暗8 h,生长至3叶1心期,进行非生物胁迫试验。低温胁迫试验是将幼苗置于4 ℃培养箱中进行处理,模拟干旱或盐胁迫处理是利用20% PEG6000或200 mmol·L-1的NaCl溶液对幼苗进行胁迫处理[35],缺氮胁迫试验,将正常生长的玉米幼苗(NH4NO3为氮素来源,总氮浓度为2 mmol·L-1)转移至铵态氮或硝态氮缺乏(用KNO3或NH4Cl替换NH4NO3)的营养液中进行胁迫处理,总氮浓度保持在2 mmol·L-1水平。在胁迫处理的0、1、6、24 h后,收集对照植株(0 h)和处理植株的第2片叶用于实时定量PCR分析。每个处理3次生物学重复,每次生物学重复取5株幼苗。
1.2.3 RNA提取及定量PCR分析 利用华越洋植物RNA快速提取试剂盒提取各样品的总RNA,检测其浓度和纯度后(NanoDrop ND-2000),按照反转录试剂盒说明书完成cDNA的合成和纯化(Prime-ScriptTMRT reagent Kit with gDNA Eraser,TaKaRa)。反转录cDNA稀释至质量浓度为100 μg·μL-1作为扩增模板。根据目的基因序列的保守性,利用Oligo 7设计特异性引物(表1),Actin作为内参基因。实时定量PCR分析利用Roche Light Cycler 480 System(Roche,Basel,Switzerland)定量检测系统,反应系统10 μL,包括 SYBR Premix EXTaqTMkit 5 μL、上下游引物各1 μL、cDNA 1 μL、ddH2O 3 μL。PCR扩增条件:95 ℃预变性 2 min;95 ℃变性 5 s,55 ℃退火 5 s, 72 ℃延伸25 s,45个循环。根据2-△△CT算法计算基因的相对表达量。对于胁迫处理的试验数据,将各处理0 h各基因表达量设定为1,并以此来计算其他时间点的相对表达量,并使用 (平均值±标准差)来表示。

表1 ZmbZIP基因表达分析的实时荧光定量引物Table 1 qRT-PCR primers for expression analysis on ZmbZIPs
2 结果与分析
2.1 ZmbZIP生物信息学分析
为了研究玉米bZIP基因的表达信息,我们选取了10个ZmbZIP基因作为本文的研究对象,其基本信息见表2。10个ZmbZIP蛋白长度从121 aa(ZmbZIP101)~246 aa(ZmbZIP3),相应的分子量从13.41 kDa~26.83 kDa。大部分ZmbZIP蛋白的理论等电点呈中性或碱性,除了ZmbZIP3(PI=5.53)和ZmbZIP101(PI=4.77)(表2)。10个ZmbZIP基因散布于8个染色体上,其中Chr1和Chr3染色体上分别有2个基因,Chr2、Chr4、Chr5、Chr6、Chr7和Chr9上分别各有一个ZmbZIP(图1A)。基因结构分析显示10个ZmbZIP基因的内含子数目变化丰富,三个基因(ZmbZIP3、ZmbZIP41和ZmbZIP52)没有内含子,其他基因各含有3~12个内含子不等(图1 B)。10个ZmbZIP蛋白N端含有典型的bZIP结构域,包括保守的碱性蛋白序列和亮氨酸拉链区 (图1 C)。这10个ZmbZIP基因在该家族进化树上的位置特殊,可归为一大类(图1 D)。进化树分析结果显示10个ZmbZIP蛋白分属于不同的5个蛋白家族,显示10个ZmbZIP蛋白家族起源于不同的祖先蛋白,并沿着各自的进化途径进化而来(图1-D)。
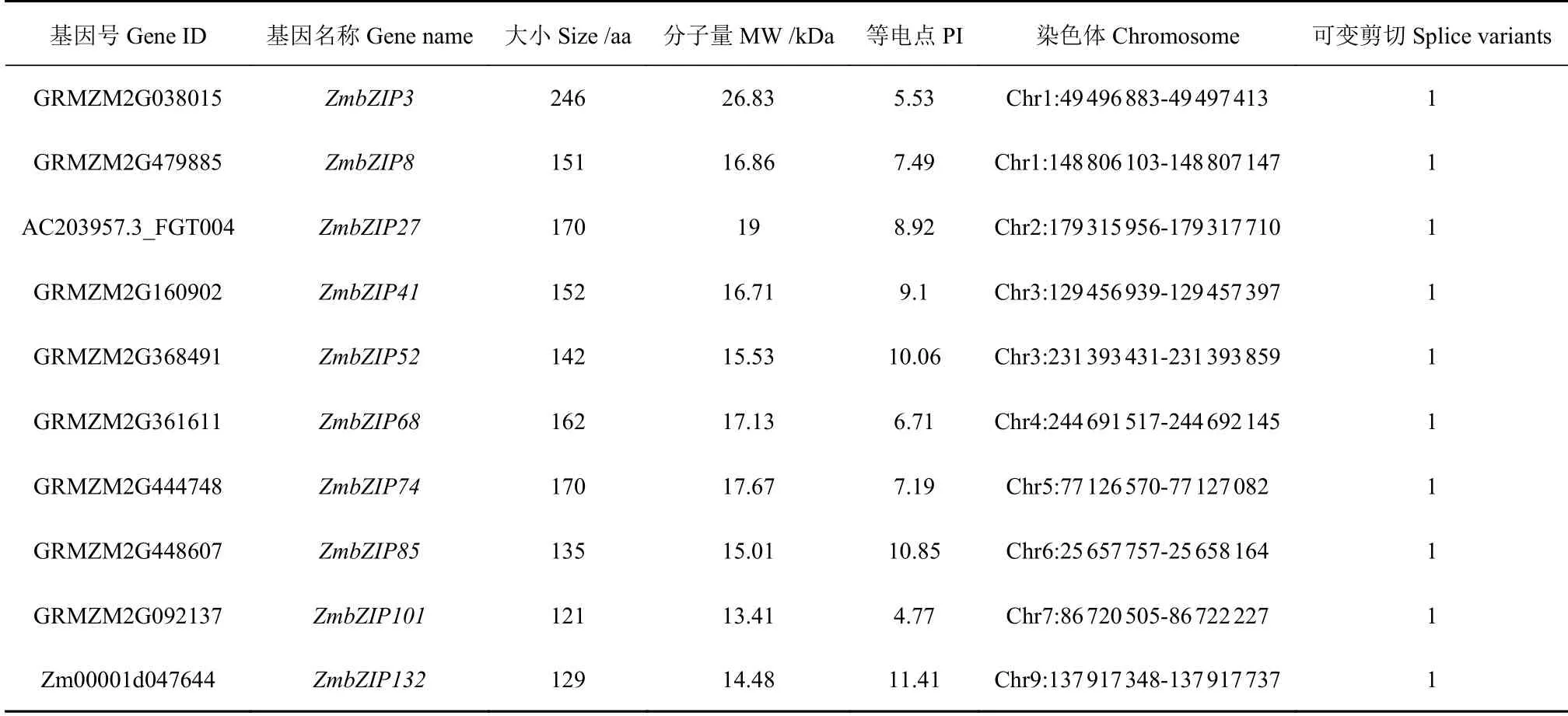
表2 10个玉米bZIP基因基本信息Table 2 Basic information on 10 bZIP genes of Z. mays
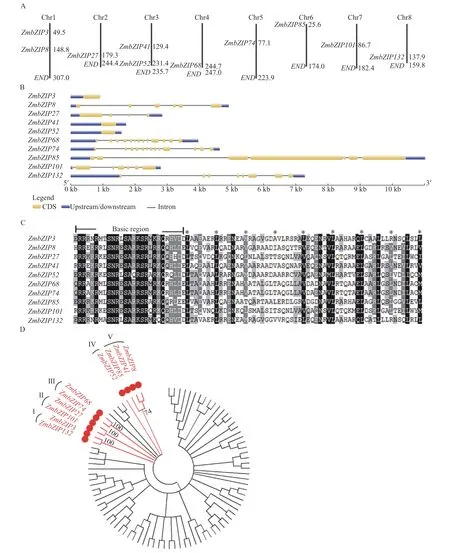
图1 10个ZmbZIPs的生物信息学分析Fig. 1 Bioinformatics of 10 ZmbZIPs
2.2 ZmbZIP组织特异性表达模式分析
为了明确ZmbZIP在玉米不同组织中的表达特异性,利用qPCR技术,在玉米授粉扬花期,检测了10个ZmbZIP基因在玉米根、茎、叶、雄花和嫩穗中的表达模式。结果显示10个ZmbZIP基因在玉米组织中的表达模式不同,7个ZmbZIP基因(ZmbZIP3、ZmbZIP8、27、41、52、101和ZmbZIP132)主要在玉米根系中表达,是其他组织器官的2倍以上,表明这些基因在玉米的根系生长发育进程中发挥着重要的生物学功能。ZmbZIP68呈组成型表达模式,ZmbZIP85主要在玉米根系和嫩穗中表达,而ZmbZIP74主要在雄花中表达,高于其他组织器官5倍以上(图2)。ZmbZIP基因表达模式分析显示该家族基因在玉米生长发育进程中发挥着重要的作用。
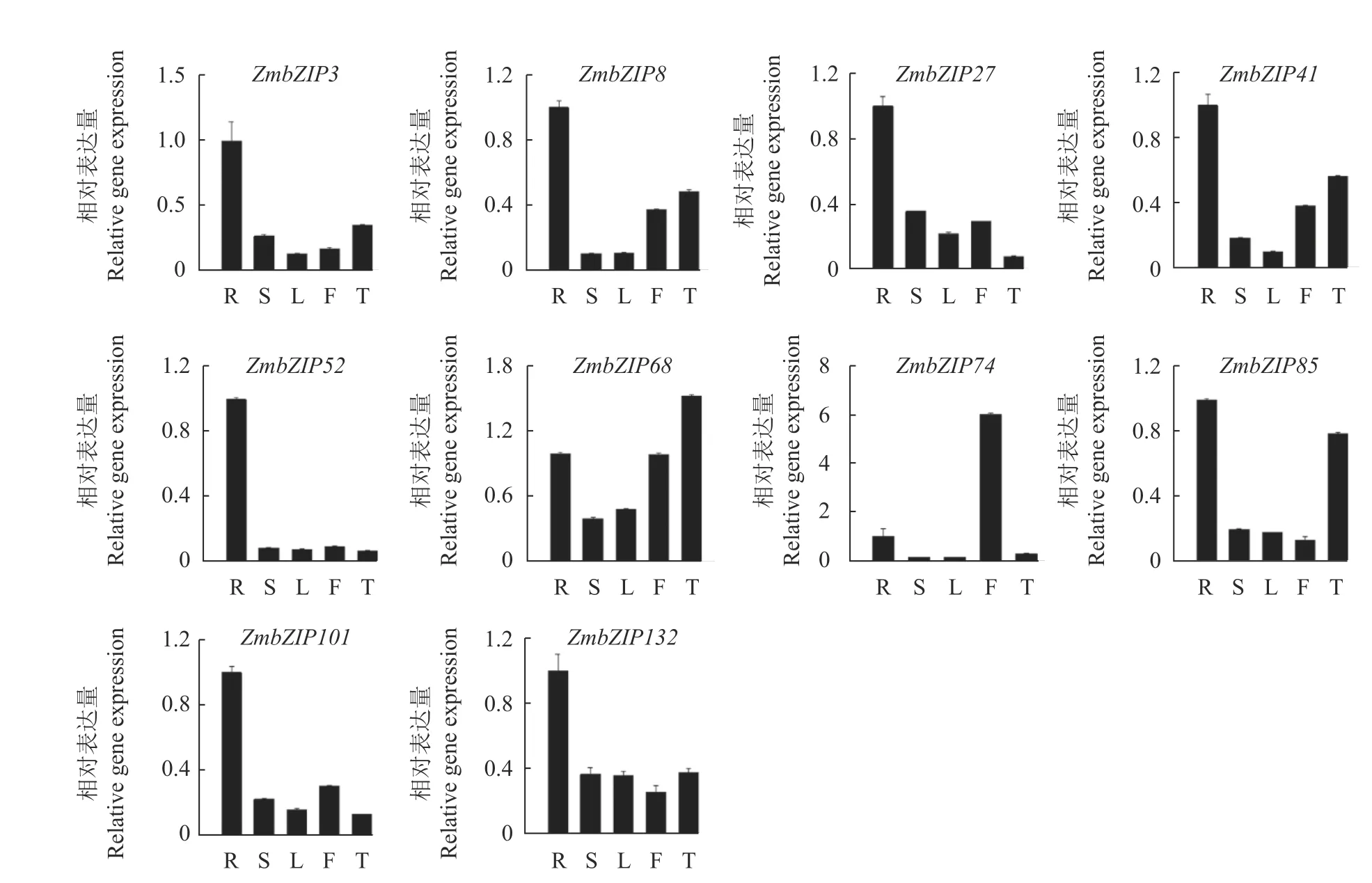
图2 ZmbZIP基因在玉米不同器官中的表达模式分析Fig. 2 Expressions of ZmbZIPs in different maize tissues
2.3 叶片中ZmbZIP对盐、干旱或低温胁迫的响应
非生物胁迫诸如干旱、高盐和低温胁迫会对作物叶片造成明显影响[36],因此,本文重点分析了玉米叶片中10个ZmbZIPs应答3种胁迫时的表达模式。由图3可知,10个ZmbZIP基因在应对不同逆境胁迫时的响应表达模式不同。在三种逆境胁迫1 h和6 h时,ZmbZIP74和ZmbZIP101的表达量分别上升了2倍以上,表明这2个基因可能同时参与3种逆境胁迫响应机制。ZmbZIP3和ZmbZIP68的表达至少受2种逆境胁迫的诱导(>2倍),在NaCl胁迫24 h和低温胁迫1 h时,ZmbZIP3表达量分别上调了4倍和21倍,ZmbZIP68表达量分别上升了至少2倍,表明这2个基因在这2种逆境胁迫中发挥着作用。ZmbZIP52、ZmbZIP85和ZmbZIP132受到NaCl胁迫的明显诱导,在胁迫处理24 h时,其表达量分别上升了4倍、2.3倍和6倍,而在PEG6000和低温胁迫时,这3个基因多数受到明显的抑制,比如在胁迫处理3 h时,ZmbZIP85的表达量分别下降了78%和60%,这些数据表明10个ZmbZIP基因在应对不同逆境胁迫时呈现不同的表达模式,预示着这些基因可能在玉米抵御不同逆境胁迫时发挥多样的功能。
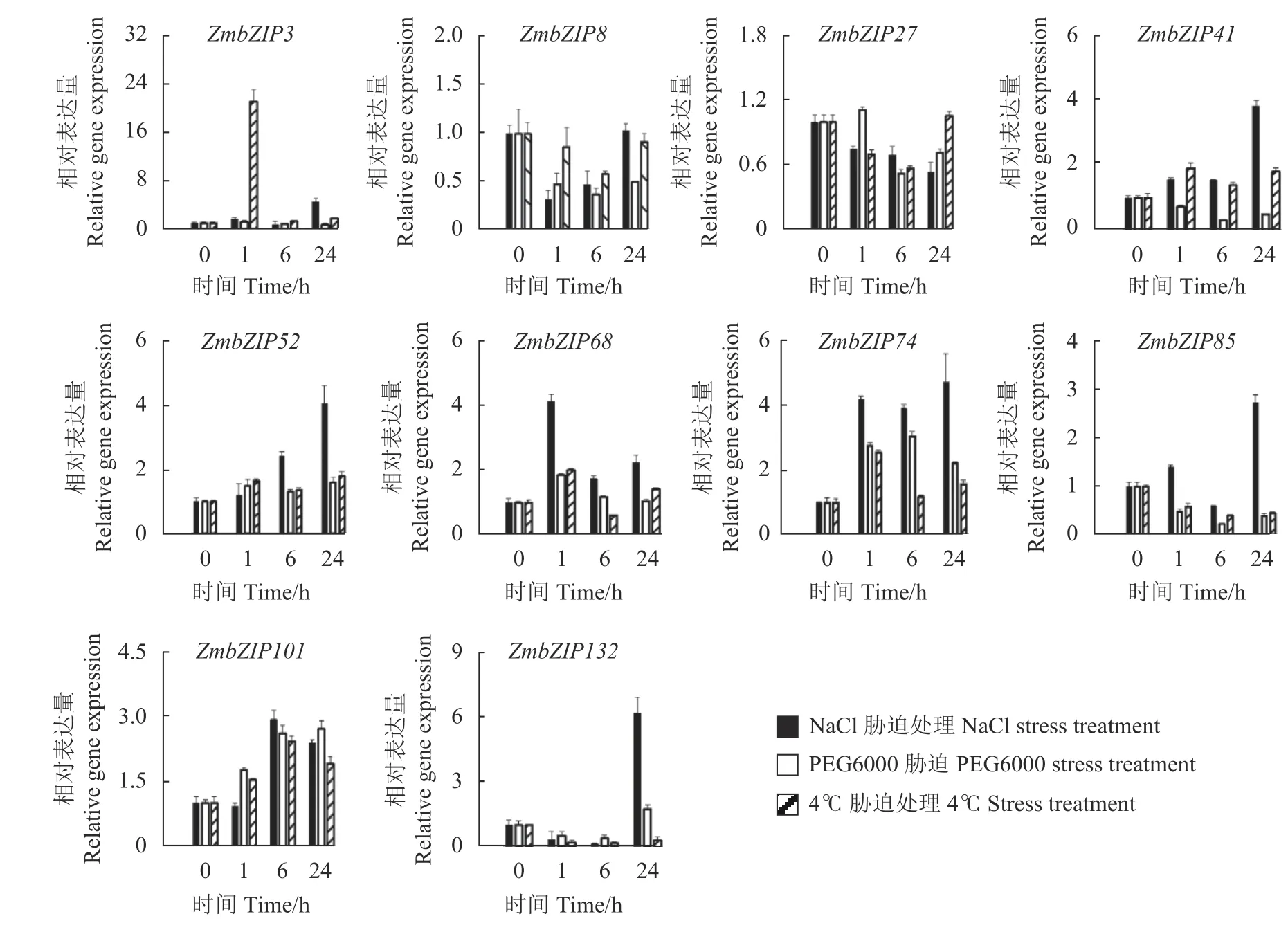
图3 玉米叶片中ZmbZIPs应答不同胁迫时的表达模式分析Fig. 3 Expression patterns of ZmbZIPs under stresses
2.4 ZmbZIP基因对氮缺乏胁迫的响应
由图4可知,叶片中ZmbZIP基因的表达模式在不同氮元素缺乏胁迫中变化比较大。一类是硝态氮缺乏胁迫对ZmbZIP基因的影响和铵态氮缺乏胁迫不同,这类包括基因有ZmbZIP3、ZmbZIP8、ZmbZIP27、ZmbZIP74、ZmbZIP85、ZmbZIP101和ZmbZIP132,这些基因中,铵态氮缺乏胁迫诱导ZmbZIP基因的表达,而硝态氮缺乏胁迫则抑制ZmbZIP基因的表达,比如ZmbZIP132在胁迫处理1 h时,硝态氮缺乏胁迫使该基因的表达下降了64%,而铵态氮缺乏胁迫则使该基因的表达上升了8倍,表明这些基因在响应硝态氮或铵态氮缺乏胁迫时的作用机制有差异。另一类是这2种胁迫处理对ZmbZIP基因的表达影响相似,如ZmbZIP68,该基因的表达同时受2类胁迫处理的抑制,表明该基因可能在应对这2类胁迫时起着类似的作用。
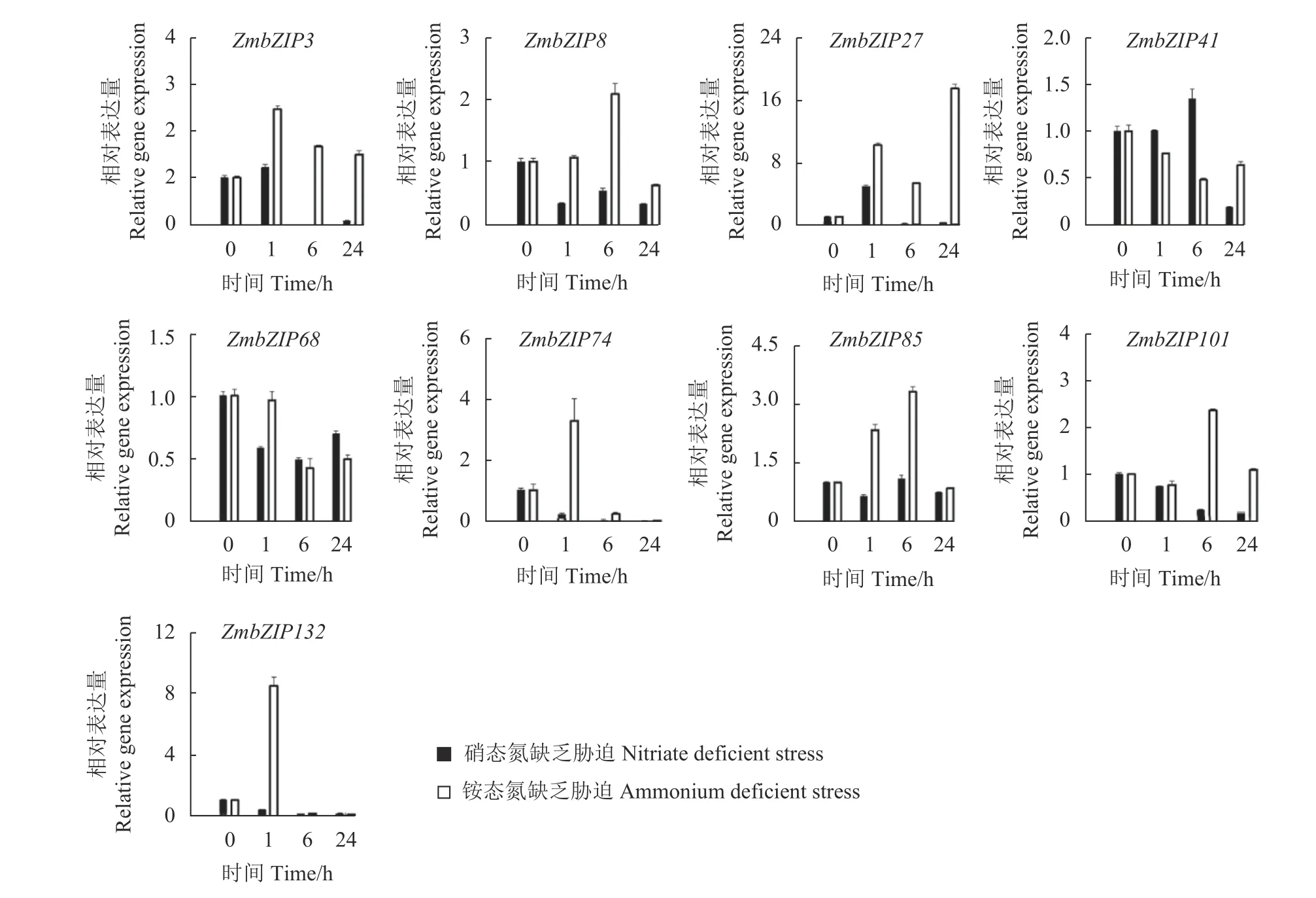
图4 玉米叶片中的ZmbZIPs应答硝态氮或铵态氮缺乏胁迫的响应表达模式Fig. 4 ZmbZIP expression patterns in maize leaves in response to nitrate or ammonium deficiency
由图5可知,玉米根系中ZmbZIP基因的表达模式受不同氮形态缺乏胁迫调控。ZmbZIP27、ZmbZIP41和ZmbZIP68同时受硝态氮和铵态氮缺乏胁迫的抑制,尤其ZmbZIP41受硝态氮缺乏胁迫的严重抑制,胁迫处理1 h时,其表达量比处理前下降超过99%,铵态氮缺乏胁迫使其表达量下降超过80%,表明这2个基因在硝态氮和铵态氮缺乏胁迫中发挥着类似作用。ZmbZIP3表达的变化趋势在硝态氮和铵态氮缺乏胁迫中类似,都是先升后降,在胁迫处理1 h时,达到最大值,之后慢慢下降。ZmbZIP8、ZmbZIP74、ZmbZIP85和ZmbZIP132的表达模式在不同氮形态胁迫中趋势相反,其中ZmbZIP8和ZmbZIP85受硝态氮缺乏胁迫的诱导而受铵态氮缺乏胁迫的抑制,而ZmbZIP74和ZmbZIP132则受铵态氮缺乏胁迫的诱导而受硝态氮缺乏胁迫的抑制。ZmbZIP基因在硝态氮和铵态氮缺乏胁迫中的不同表达模式预示着该基因在玉米应对氮缺乏胁迫中的不同作用。
图5 玉米根系中的ZmbZIPs在硝态氮或铵态氮缺乏胁迫时的表达模式分析Fig. 5 ZmbZIP expression patterns in maize roots in response to nitrate or ammonium deficiency
3 讨论与结论
作为粮经饲三元粮食作物,玉米在保障全球及我国粮食安全方面发挥着重要作用。然而在其生长周期内,经常遭受各类逆境胁迫,造成产量减产,使种植者遭受经济损失。所以挖掘控制逆境胁迫的遗传因子,解析其调控的分子生理机理,对于培育抗逆性好、高产稳产的玉米新品种显得意义重大。bZIP蛋白成员广泛分布于植物基因组中,在调控作物抗逆性方面发挥着重要作用。本文中,逆境胁迫广泛的调控bZIP基因的表达,这与前人的研究类似。不同逆境胁迫对ZmbZIP85基因影响因玉米种质的不同而各异,比如,玉米自交系Han21胁迫处理时,该基因的表达没有发生明显改变,而在自交系Ye478中,在严重胁迫时该基因的表达明显上调[6]。拟南芥中ZmbZIP85的同源基因AtbZIP1是逆境胁迫的正调控因子,提高该基因的表达能显著的增强拟南芥的抗逆性[6]。而在本研究中,该基因应答盐、干旱和低温胁迫时,表达模式各异,200 mmol·L-1的NaCl溶液显著提高该基因的表达,而20% PEG6000或4 ℃低温胁迫明显抑制该基因的表达,反映了该基因在不同玉米种质中应答不同逆境胁迫时的不同调控机制。类似的结果在其他研究中也有报道,以玉米Yu882为试验材料,ZmbZIP8、ZmbZIP41、ZmbZIP52、ZmbZIP68和ZmbZIP85明显受到盐胁迫的诱导[37],而在我们的试验中,ZmbZIP8受到盐胁迫抑制,反映了该基因在不同种质中的生物学功能差异。
植物根系依靠硝态氮转运体或铵态氮转运体来吸收土壤中的氮素,在这一过程中,大量转录因子成员参与调控氮素吸收和代谢平衡。超表达玉米DOF1基因能提高拟南芥吸收氮素能力[38],该基因也能提高生长于低产土壤中水稻的产量,增强氮素吸收利用效率[39]。小麦TaNFYA1-6B和TaNAC2-5A能促进根系发育和激发NTR1和NRT2家族基因的表达水平,提高氮素的吸收效率和小麦的产量[40-41]。干旱胁迫下,小麦氮饥饿诱导基因TabZIP2也参与协调碳氮代谢平衡,提高小麦抗旱能力[42]。在本文中,叶片或根系中ZmbZIP基因广泛受硝态氮或铵态氮缺乏胁迫的调控,在叶片或根系有些基因受两种胁迫处理诱导的表达模式类似,比如,ZmbZIP3、ZmbZIP68、ZmbZIP74和ZmbZIP101等4个基因,而其他基因在叶片或根系受2种胁迫处理的影响不同,比如,叶片ZmbZIP27同时受2种胁迫处理的抑制,而根系中的ZmbZIP27则同时受到诱导而上调,表现了该基因在叶片或根系等不同组织中的不同生物学功能。
