海水铝浓度对海链藻活性、生产力及其固碳性的影响
2021-02-03周洁玉周军明魏辉煌李梦圆宋雅然刘红昌
周洁玉, 刘 冬, 3, 田 倩, 周军明, 魏辉煌, 李梦圆, 宋雅然, 刘红昌
海水铝浓度对海链藻活性、生产力及其固碳性的影响
周洁玉1, 2, 刘 冬1, 2, 3, 田 倩1, 2, 周军明1, 2, 魏辉煌1, 2, 李梦圆1, 2, 宋雅然1, 2, 刘红昌4
(1. 中国科学院广州地球化学研究所, 矿物与成矿重点实验室, 广东 广州 510640; 2. 中国科学院大学, 北京 100049; 3. 近海海洋环境科学国家重点实验室(厦门大学), 福建 厦门 361005; 4. 中南大学, 资源加工与生物工程学院, 湖南 长沙 410083)
近年来, 随着人类工业活动的增多及强淋溶性酸雨的频繁出现, 大量的铝被输入到海洋中, 导致海水环境中铝浓度的急剧增加。由于可溶性铝对绝大多数水生生物具有生物毒害性, 其浓度的增加势必将影响到水生生物, 尤其是对环境变化较为敏感的水生植物等的生命活动及其所涉及的元素地球化学循环作用。作为全球生产力的主要贡献者, 硅藻提供了40%的海洋初级生产力, 并通过“生物泵”作用, 调控着全球碳循环。海水中铝浓度的增加, 是否将影响其生物作用及其固碳能力, 尚不清楚。基于上述问题, 本研究通过对近海典型硅藻(威氏海链藻)的实验室富铝培养, 以探讨可溶性铝浓度对该硅藻的生物活性、生产力及其所驱动的碳元素的生物地球化学循环的影响及机制。结果表明: 当海洋环境中铝浓度略有升高时(≤50 nmol/L), 海链藻未发现有铝毒害产生, 其初级生产力反而有所提高。然而, 不同铝浓度条件下, 海链藻的固碳能力表现出明显差异: 在铝浓度为20 nmol/L时, 海链藻的固碳能力较之未添加铝时有显著提高。对铝迁移的研究发现, 环境中的铝可被硅藻通过生物作用捕获, 并用于其硅质壳体的构建。上述结果表明, 海水中铝浓度对于硅藻的生命活动有显著影响, 也可以影响硅藻的固碳能力, 进而影响到硅藻在全球海洋“生物泵”中的作用。
硅藻; 铝捕获; 海洋初级生产力; 固碳; 硅藻壳体
硅藻是水体中分布最为广泛的单细胞藻类之一, 其种类繁多——是海洋中种群最多的自养型浮游生物。海洋硅藻贡献了全球初级生产力的40%, 相当于所有陆地热带雨林的总和[1-4], 并对全球范围内的碳、氮、氧、硅等元素的地球化学循环均有重要影响。硅藻对于生长环境的变化较为敏感, 环境因子的变动对其生长、生殖会产生较大影响, 进而影响到其在元素循环中的作用。以碳循环为例, 研究表明, 大洋寡铁区域铁离子浓度的升高, 将大大促进对铁元素敏感的硅藻藻属(如拟菱形藻等)的生长, 进而提高该区域硅藻的整体生产力[5-7]。基于该发现, Martin在1990年提出了“铁假说”[8], 认为铁的供给增加能够使硅藻等浮游生物勃发, 从而提高海洋输出生产力并降低大气CO2含量, 该理论促成了全球十余次中等尺度规模的海洋施加铁肥, 用以调控全球大气CO2含量的实验[9]。可见, 水体环境的变化, 是影响硅藻“固碳”能力的重要因素。
铝是地壳中含量最高的金属元素, 约占地壳总量的8.0%, 主要以铝硅酸盐的形式存在于岩石和矿物中[10]。近年来随着人类工业的飞速发展, 水体中, 尤其是海洋中的铝浓度逐渐增高[11]。其主要原因有: 1) 含铝废水的大量排放, 使得铝被直接或间接地输入海洋; 2) 工业化导致大气中二氧化碳浓度升高, 使得酸雨愈发频繁, 岩石和土壤所遭受的淋溶风化作用更加强烈, 更多的铝被释放到水体。加之海洋酸化, pH值降低, 使得可溶性铝浓度逐渐增加[12]。铝被认为对大多数水生生物具有一定毒性, 且其浓度越高, 毒性越强[13]。其生物毒害机制主要为: 导致生物的呼吸障碍及对体内离子的调节性障碍[14]。因此, 随着海洋中铝浓度的升高, 海洋硅藻的生命活性及其生产力是否受到影响, 以及这些影响对其所参与的“碳生物泵”效率的作用机制, 尚不清楚。
针对上述问题, 本研究选取近海分布最广的硅藻之一——海链藻为研究对象。通过实验室培养实验中添加铝的方式, 模拟海洋环境中铝浓度的增加, 以分析硅藻生命活性及初级生产力对环境铝浓度升高的响应机制, 进而探明其对海链藻固碳能力的影响及机制。本研究中, 叶绿素的含量作为衡量初级生产力的指标。这是由于硅藻的初级生产力可以根据硅藻叶绿素的含量等因子进行评估[15]。
1 材料与方法
1.1 实验材料
海链藻藻种采自中国南海惠州海域(考洲洋), 经实验室分离、纯化后保存于4℃冰箱中, 随后于培养箱中进行扩大培养。经多代无铝培养后所获海链藻藻种通过能量色散X射线光谱(EDS)进行线扫描元素分析(分析方法等详见1.6), 未发现铝赋存。
1.2 硅藻加铝培养
海链藻在光照培养箱(上海一恒科学仪器有限公司)中培养, 培养瓶为1 L锥形瓶, 海水培养液由人工海水(成分详见表1)和f/2培养基(NaNO3、NaH2PO4、微量元素、维生素、Na2SiO3)组成。以AlCl3作为铝源,使用去离子水配制0.001 mol/L AlCl3溶液加入上述硅藻培养液中, 分别获得铝浓度为10 nmol/L、20 nmol/L和50 nmol/L 的培养液。培养液的初始pH为8.41, 海链藻的培养温度为25℃, 明暗周期为12 h/12 h, 光照强度为100 µEm–2s–1, 培养时间为15天(海链藻的生长周期)。为避免外源铝污染, 本实验中所有使用的培养器皿均使用盐酸等进行了去微量金属清洗, 人工海水和培养液等也为无铝试剂配制[16]。
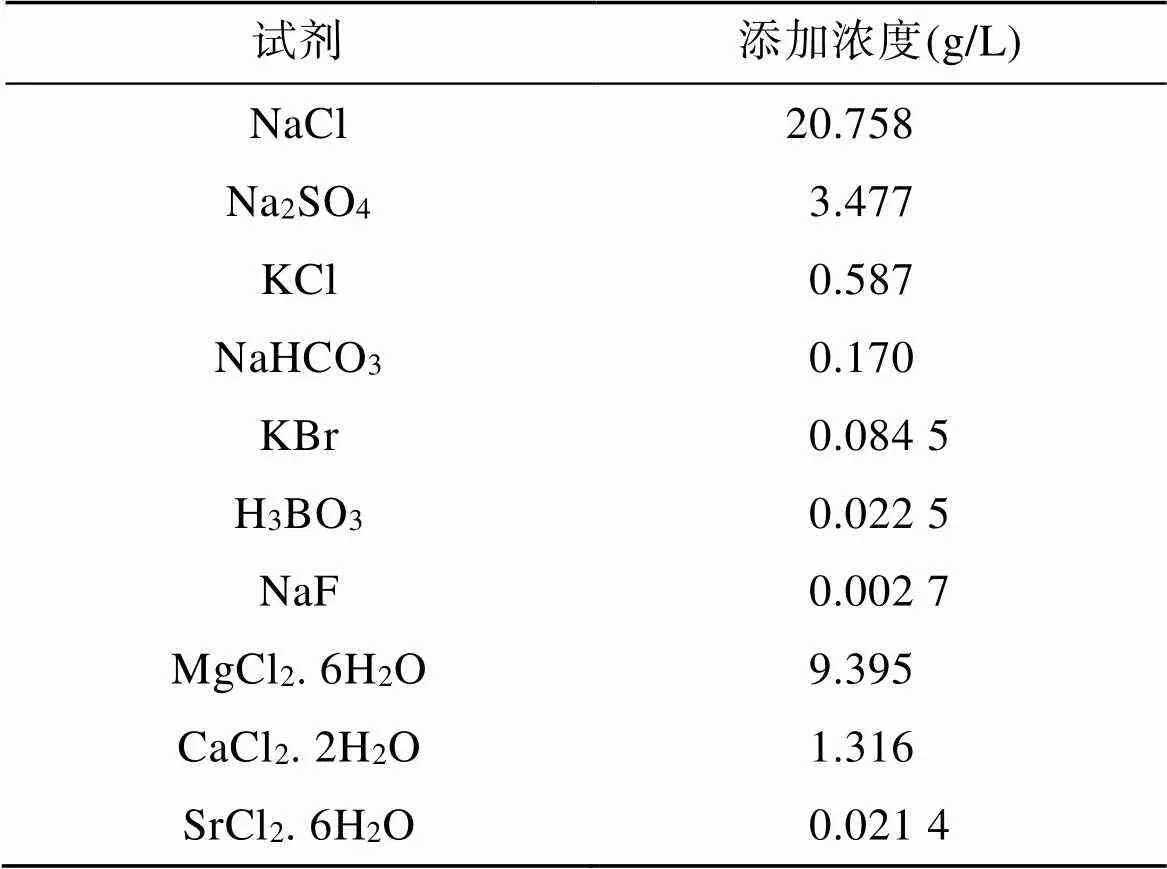
表1 人工海水配方
1.3 硅藻计数
海链藻的初始细胞密度是4.74×104个/mL, 培养期间每天取样对细胞进行取样计数。具体步骤为: 晃动锥形瓶, 使锥形瓶中的海链藻分布均匀, 从每个锥形瓶中分别取2.0 mL样品。于计数板中各加入20 µL样品, 使用Count Star细胞计数仪(上海睿钰生物科技有限公司)对其进行计数, 并计算出海链藻的细胞密度。为保证数据准确性, 每次进行3次平行样品测试, 使用平均值进行计算。
1.4 叶绿素a提取及分光光度法测定叶绿素a浓度
为保证较高精确度, 选在不同铝添加样品细胞密度的极大值日取样。分别取1.5 mL含海链藻的培养液置于离心管中, 并于高速离心机(德国Sartorius Sigma 1-14)中以15 000 rmp转速离心7 min, 将上层清液弃去, 所获固体为海链藻细胞。向上述海链藻细胞中加入1.0 mL无水乙醇(4℃)[17-18], 用旋涡混匀仪使细胞悬浮, 将装有悬浮液的离心管用铝箔纸封口并置于4℃冰箱保存24 h后, 于高速离心机中以15 000 rmp转速离心7 min。收集上清液并使用分光光度计(UV 2400型, 上海舜宇恒平科学仪器有限公司)在665 nm和649 nm处分别测定其吸收值。叶绿素的浓度计算公式为:
CChl-a= 13.7 ×665– 5.76 ×649
式中: CChl-a为叶绿素的浓度, 单位为 μg/mL;665为波长在665 nm处的吸光值;649为波长在649 nm处的吸光值; 13.7和5.76为该分光光度方程的系数[18]。
1.5 处理海链藻及海链藻壳体样品
待15天培养结束后, 离心, 用纯水反复洗涤, 并冷冻干燥, 以保留海链藻的完整组分。对部分海链藻样品进行化学处理, 在30%过氧化氢溶液中浸泡72 h[19], 至样品变白, 以去除其有机质, 随后用纯水洗涤。重复上述浸泡和洗涤过程, 直至硅藻所有有机组份完全去除(经碳含量测试评估, 碳含量低于仪器检测限)。将离心所获海链藻壳体干燥后保存待测。
1.6 透射电镜(TEM)、能量色散X射线光谱仪(EDS)及碳含量测试
TEM及EDS测试: 将硅藻及壳体样品与水混合, 然后摇晃使其分散, 将分散液滴在铜网上, 待表面干燥后, 用透射电子显微镜对单个硅藻进行定位以及用EDS进行元素分析, 以确定铝元素的赋存。TEM型号为: FIE Talos F200S (测试电压为200 kV, 测试电流为1 nA), 该仪器自带了一个EDS系统, EDS型号为: FEI Super-X。
碳含量测试: 取上述硅藻样品约2.00 mg, 放入6×12 mm的锡舟中, 用Vario ELⅢ元素分析仪对海链藻进行碳元素分析, 每个样品进行三次平行测试, 取平均值用于分析。仪器检测限40 ppm。
1.7 数据分析方法
采用了单因素方差分析(ANOVA), 以值0.05作为衡量显著性差异的标准, 用SPSS和Origin 8.0对所得数据进行处理和作图。
2 结果与讨论
2.1 富铝环境对海链藻生长的影响
为评估环境铝浓度对海链藻分裂增殖的影响, 本研究对不同初始铝浓度(对照组、10 nmol/L添加组、20 nmol/L添加组和50 nmol/L添加组)培养液中所获海链藻细胞密度进行了分析。计数仪测试结果表明, 海链藻前期增长速度极快, 到第5天时, 增长速度放缓进入稳定期, 其藻细胞密度于第8天达到极大值(图1)。随后, 在第12天左右, 其细胞密度逐渐下降, 这是由于部分硅藻达到生命周期后不再分裂, 并有部分硅藻死亡所致。对于富铝培养液中的硅藻, 在不同铝添加组中, 其表现出不同的分裂生殖特征。在海链藻的指数生长期内, 加铝组的细胞分裂速度相较于对照组均有所升高。需要注意的是, 对照组和铝添加组中的海链藻细胞密度极大值并无显著性差异(>0.05), 这表明在上述初始铝浓度下, 添加铝对硅藻生长、生殖所产生的生物总量影响较小, 这也表明上述铝添加浓度(10、20和50 nmol/L)对海链藻的生长无明显毒害作用。
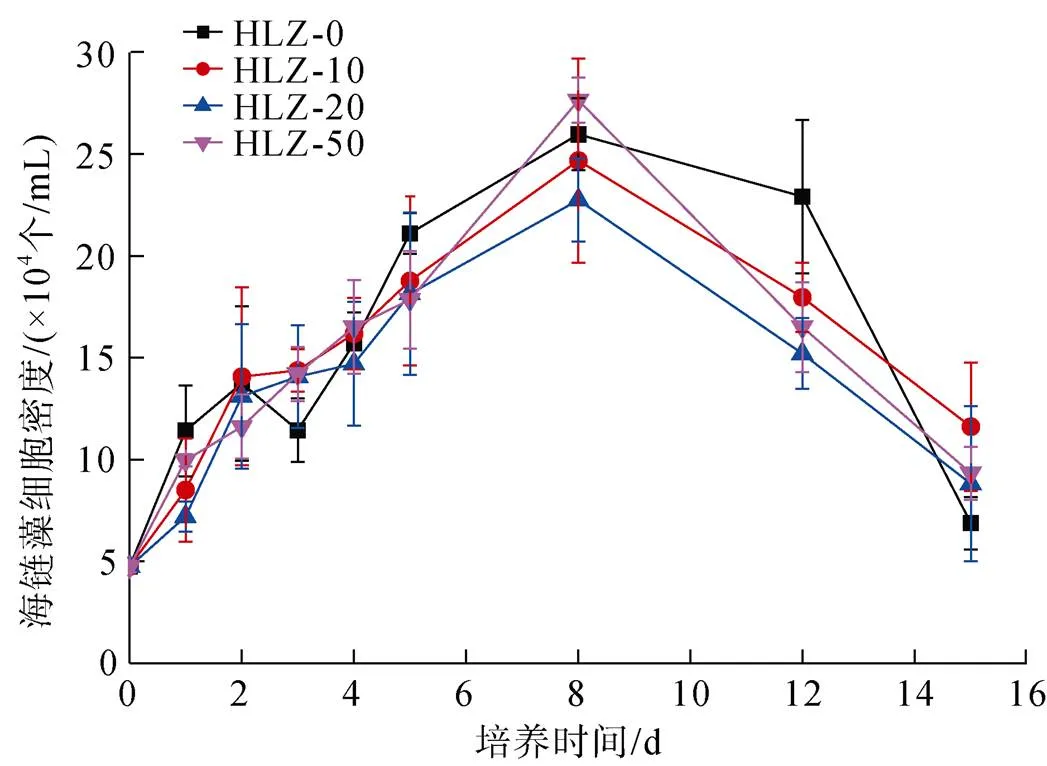
图1 添加/未添加铝培养液中不同培养时间的海链藻细胞密度
2.2 富铝环境对海链藻初级生产力的影响
本研究通过测定不同铝添加组下的海链藻的叶绿素含量, 用以评估加铝对其初级生产力的影响。使用低温-乙醇法提取硅藻液中的叶绿素, 并经紫外分光光度计测得其吸光值, 通过分光光度方程计算得到叶绿素浓度。结果表明, 铝添加组和对照组中的叶绿素浓度并无显著性差异(>0.05)。然而, 考虑到测试所选取的海链藻溶液中, 海链藻的细胞密度不同, 我们对数据进行了标化处理, 以获得对照组和不同铝添加组中单个海链藻细胞叶绿素的含量(图2)。结果表明, 随着添加铝初始浓度的增加, 单位海链藻的叶绿素含量先升高后降低, 其中20 nmol/L添加组的单位海链藻叶绿素含量显著高于对照组的值(<0.05)。值得注意的是, 即使初始铝浓度高达50 nmol/L, 单位海链藻的叶绿素含量也高于对照组的样品值。这表明, 一定浓度(≤50 nmol/L)的铝可有效提高海链藻的初级生产力, 而在上述几组不同铝添加组中, 20 nmol/L添加组具有最好的效果。
2.3 富铝环境对海链藻“固碳”能力的影响
死亡的硅藻通过“携带”自身合成的有机质穿过水-沉积层界面进入埋深, 实现“固碳”作用, 并成为“生物泵”的重要环节[4]。为评估富铝环境对海链藻“固碳”能力的影响, 本实验对培养所获硅藻的含碳量进行了分析: 待15天培养结束后, 通过元素分析的方法, 分别对对照组、10 nmol/L添加组、20 nmol/L添加组及50 nmol/L添加组所获硅藻样品的含碳量进行测定。结果表明, 在20 nmol/L添加组中培养的海链藻, 含碳量显著高于另外三组(<0.05)(图3)。由于本测试针对的是硅藻自身所含碳的评估, 因此该结果表明, 在铝浓度为20 nmol/L时, 硅藻固碳能力显著提高。
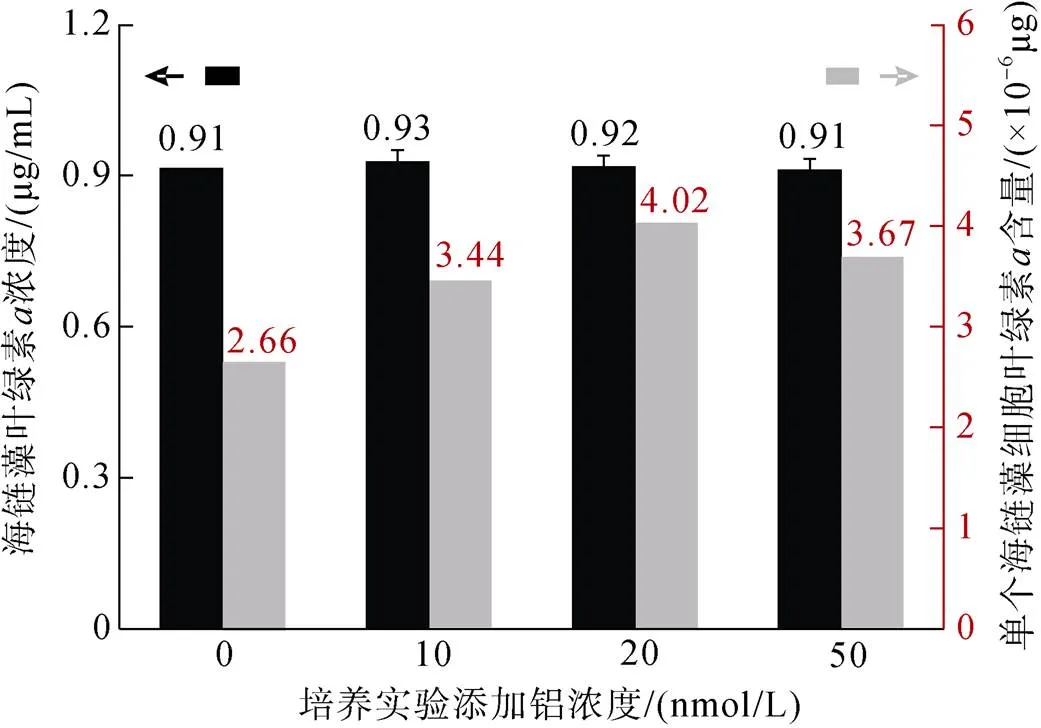
图2 不同铝浓度培养下海链藻的叶绿素a浓度以及单个海链藻细胞的叶绿素a含量
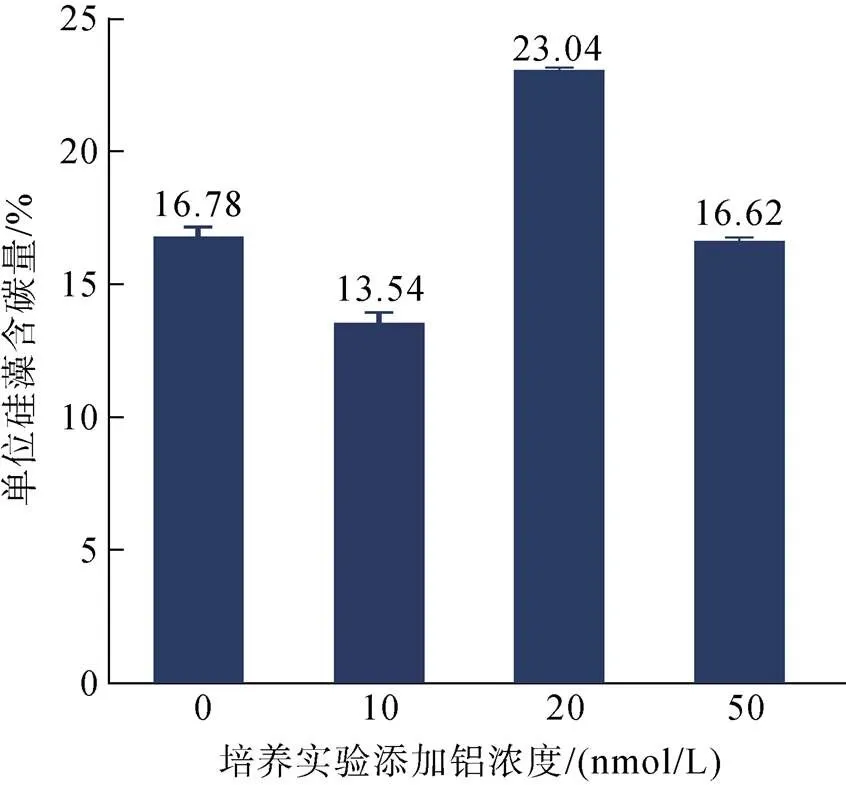
图3 不同铝浓度下培养所获海链藻的含碳量
2.4 富铝环境培养所得海链藻中铝的赋存特征
前人在开展铝在海洋中的生物地球化学循环研究时曾发现: 硅藻可以通过生物捕获作用由环境中获取铝[20]。进一步的研究发现, 海水中铝的浓度受控于硅藻, 基于此, 研究者提出了生物驱动下的海洋中硅、铝共循环观点[21-23]。通过对硅藻中铝进行探测发现, 铝很有可能赋存在硅藻壳体结构中[24-27]。基于培养实验和沉积物中硅藻壳体的元素分析等, Koning等认为壳体中铝的赋存量(按照铝硅摩尔比计)为10–4~10–3[27]。然而, 由于受到硅藻壳体提纯困难及原位微区分析技术缺乏的影响, 铝在硅藻壳体结构中的赋存量仍不明确[28-30]。
为探明富铝环境中铝的生物迁移机制, 解决上述问题, 本研究利用微区探测的方法, 在微-纳米尺度对铝在海链藻中的赋存特征进行了探索。选取20 nmol/L添加组中所获海链藻和壳体为研究对象, 使用TEM进行观测, 并结合EDS对硅藻和壳体中的Si和Al分布和含量等进行了分析。结果表明, 铝赋存在海链藻中(图4a和图4c), 且在硅藻壳体中有较均匀的分布(图4d和图4f)。EDS线扫描结果表明, 20 nmol/L添加组中所获海链藻的铝含量, 以铝硅摩尔比计, 为0.055; 而其壳体的铝硅摩尔比为0.009。可见, 壳体中的确赋存有铝, 但其含量较低。TEM-EDS微观分析手段可为铝的赋存及分布特征提供直接可视化的证据。
上述结果表明, 富铝培养液中的铝可通过硅藻的生物作用进入海链藻体内, 并作为骨架元素赋存于其硅质骨架中。该结果进一步验证了前人的研究[20-30], 为硅藻驱动的铝海洋生物地球化学循环提供了直接证据。
3 结论
本研究通过实验室富铝培养, 模拟海洋环境中铝浓度(≤50 nmol/L)对海链藻的活性、生产力及其“固碳”性的影响及机制。结果表明, 在上述浓度下, 1) 铝未对海链藻产生毒害; 2) 铝可加快海链藻指数生长期的分裂; 3) 铝提升了海链藻的初级生产力, 且在铝浓度为20 nmol/L时, 其效果最佳; 4) 在铝浓度≤20 nmol/L时, 海链藻的“固碳”能力有所提高, 但当铝浓度达到50 nmol/L时,“固碳”能力降低。另外, 本研究发现, 铝可以被海链藻主动捕获, 并用于构建其骨架。上述结果表明, 在海链藻生长活动和其所参与的元素生物地球化学循环过程中, 水体中铝的浓度应被作为一个重要的环境变量加以考虑。
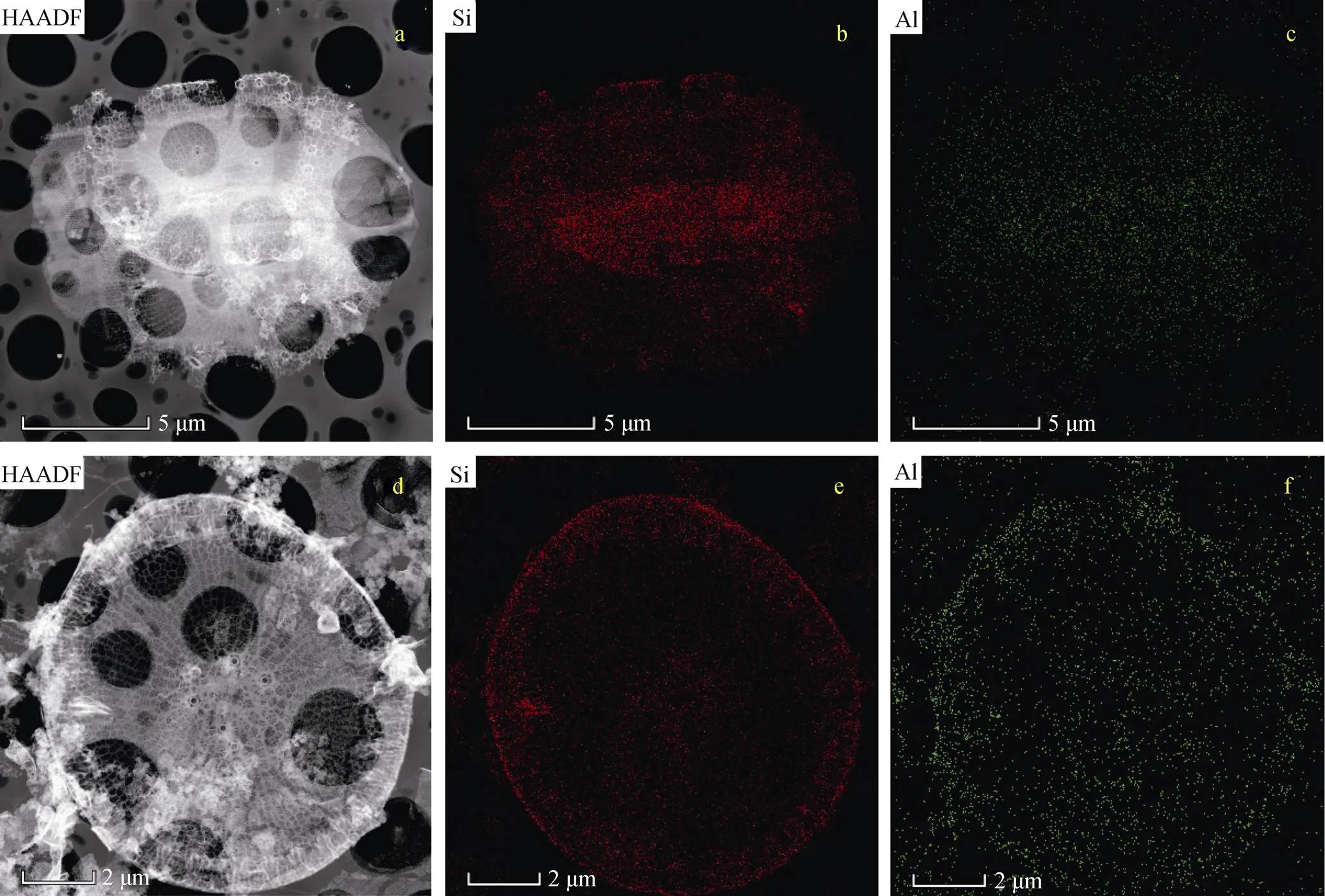
图4 海链藻TEM图
注: a: 海链藻高角环形暗场(HAADF)像; b: 海链藻硅分布图; c: 海链藻铝分布图; d: 海链藻壳体TEM-HAADF像; e: 海链藻壳体硅分布图; f: 海链藻壳体铝分布图
[1] Armbrust E V. The life of diatoms in the world’s oceans[J]. Nature, 2009, 459(7244): 185-192.
[2] Field C B, Behrenfeld M J, Randerson J T, et al. Primary production of the biosphere: Integrating terrestrial and oceanic components[J]. Science, 1998, 281(5374): 237-240.
[3] Nelson D M, Treguer P, Brzezinski M A, et al. Production and dissolution of biogenic silica in the ocean- revised global estimates, comparison with regional data and relationship to biogenic sedimentation[J]. Global Biogeochemical Cycles, 1995, 9(3): 359-372.
[4] Treguer P, Bowler C, Moriceau B, et al. Influence of diatom diversity on the ocean biological carbon pump[J]. Nature Geoscience, 2018, 11(1): 27-37.
[5] Anderson M A, Morel F M M. The influence of aqueous iron chemistry on the uptake of iron by the coastal diatom[J]. Limnology and Oceanography, 1982, 27(5): 789-813.
[6] Brand L E, Sunda W G, Guillard R R L. Limitation of marine phytoplankton reproductive rates by zinc, manganese, and iron[J]. Limnology and Oceanography, 1983, 28(6): 1182-1198.
[7] Shoenfelt E M, Sun J, Winckler G, et al. High particulate iron(II) content in glacially sourced dusts enhances productivity of a model diatom[J]. Science Advances, 2017, 3(6): e1700314.
[8] Martin J H. Glacial-interglacial CO2change: the iron hypothesis[J].Paleoceanography, 1990, 5(1): 1-13.
[9] Stoll H M. 30 years of the iron hypothesis of ice ages[J]. Nature, 2020, 578(7795): 370-371.
[10] Taylor S R. Abundance of chemical elements in the continental crust: a new table[J]. Geochimica Et Cosmochimica Acta, 1964, 28(8): 1273-1285.
[11] Gillmore M L, Golding L A, Angel B M, et al. Toxicity of dissolved and precipitated aluminium to marine diatoms[J]. Aquatic Toxicology, 2016, 174: 82-91.
[12] Orr J C, Fabry V J, Aumont O, et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms[J]. Nature, 2005, 437(7059): 681- 686.
[13] Gensemer R W, Playle R C. The bioavailability and toxicity of aluminum in aquatic environments[J]. Critical Reviews in Environmental Science and Technology, 1999, 29(4): 315-450.
[14] Xie J, Bai X, Lavoie M, et al. Analysis of the proteome of the marine diatomexposed to aluminum providing insights into aluminum toxicity mechanisms[J]. Environmental Science & Technology, 2015, 49(18): 11182-11190.
[15] 潘友联. 叶绿素与海洋初级生产力[J]. 海洋科学, 1987, 11(1): 63-65. Pan Youlian. Chlorophyll and marine primary productivity[J]. Marine Sciences, 1987, 11(1): 63-65.
[16] Liu Q, Zhou L, Liu F, et al. Uptake and subcellular distribution of aluminum in a marine diatom[J]. Ecotoxicology and Environmental Safety 2019, 169: 85-92.
[17] Porra R J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophyllsand[J]. Photosynthesis Research, 2002, 73(1): 149-156.
[18] Ritchie R J. Consistent sets of spectrophotometric chlorophyll equations for acetone, methanol and ethanol solvents[J]. Photosynthesis Research, 2006, 89(1): 27-41.
[19] Ingall E D, Diaz J M, Longo A F, et al. Role of biogenic silica in the removal of iron from the Antarctic seas[J]. Nature Communications, 2013, 4(3): 1981.
[20] Martin J H, Knauer G A. The elemental composition of plankton[J]. Geochimica et Cosmochimica Acta, 1973, 37(7): 1639-1653.
[21] Mackenzie F T, Stoffyn M, Wollast R. Aluminum in seawater: control by biological activity[J]. Science, 1978,199(4329): 680-682.
[22] Stoffyn M. Biological control of dissolved aluminum in seawater-Experimental evidence[J]. Science, 1979, 203(4381):651-653.
[23] Moran S B, Moore R M. Evidence from mesocosm studies for biological removal of dissolved aluminium from sea water[J]. Nature, 1988, 335(6192): 706-708.
[24] Van Bennekom A J, Fred Jansen J H, Vander Gaast S J, et al. Aluminium-rich opal: an intermediate in the preservation of biogenic silica in the Zaire (Congo) deep- sea fan[J]. Deep Sea Research Part A-Oceanographic Research Papers, 1989, 36(2): 173-190.
[25] Van Bennekom A, Buma A, Nolting R. Dissolved aluminium in the Weddell-Scotia Confluence and effect of Al on the dissolution kinetics of biogenic silica[J]. Marine Chemistry, 1991, 35: 423-434.
[26] Gehlen M, Heinze C, Maier-Reimer E, et al. Coupled Al-Si geochemistry in an ocean general circulation model: A tool for the validation of oceanic dust deposition fields?[J]. Global Biogeochemical Cycles, 2003, 17(1): 1028.
[27] Koning E, Gehlen M, Flank A M, et al. Rapid post- mortem incorporation of aluminum in diatom frustules: Evidence from chemical and structural analyses[J]. Marine Chemistry, 2007, 106(1-2): 208-222.
[28] Gehlen M, Beck L, Calas G, et al. Unraveling the atomic structure of biogenic silica: evidence of the structural association of Al and Si in diatom frustules[J]. Geochimica et Cosmochimica Acta, 2002, 66(9): 1601-1609.
[29] Zhou L, Tan Y, Huang L, et al. Aluminum effects on marine phytoplankton: implications for a revised Iron Hypothesis (Iron-Aluminum Hypothesis)[J]. Biogeochemistry, 2018, 139(2): 123-137.
[30] 任景玲. 长江流域及黄、东海铝的生物地球化学循环及其影响因素研究[D]. 山东青岛: 中国海洋大学, 2010. Ren Jingling. Biogeochemical cycle study of aluminum in the Changjiang Drianage basin, the Yellow Sea and the East China Sea[D]. Qingdao, Shandong Province: China Ocean University, 2010.
Effects of aluminum concentration in seawater on the activity, productivity, and carbon sequestration of
ZHOU Jie-yu1, 2, LIU Dong1, 2, 3, TIAN Qian1, 2, ZHOU Jun-ming1, 2, WEI Hui-huang1, 2, LI Meng-yuan1, 2, SONG Ya-ran1, 2, LIU Hong-chang4
(1. Key Laboratory of Minerals and Mineralization, Guangzhou Institude of Geochemistry, Chinese Academy of Sciences, Guangzhou 510640, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Key Laboratory of Offshore Marine Environmental Science (Xiamen University), Xiamen 361005, China; 4. School of Resource processing and Biological Engineering, Central South University, Changsha 410083, China)
Recently, huge amounts of aluminum (Al) have been imported into oceans due to increasing industrial activities and intensive eluviation of soils and rocks by acid rain, thus consequently resulting in a rapid increase of Al concentration in seawater. The biological activity and related biological driven geochemical cycling of most aquatic organisms that are sensitive to environmental changes will consequently be influenced by such Al concentrations increment, given the biological toxicity of Al. As an important contributor to global productivity, diatoms provide 40% of marine primary productivity and regulate the global carbon cycle through a “biological pump.” However, whether the increase in aluminum concentration in seawater will affect its biological activities and its carbon sequestration capacity remains unclear. In this study, we aimed to investigate the effect and mechanism of Al concentrations on biological activity and productivity of one marine diatom and its driven biogeochemical cycle of carbon., a typical offshore diatom, was selected as a model of marine diatoms and cultured in Al-rich medium. Results showed no toxic effect onwhen Al concentrations were ≤50 nmol/L. Nonetheless, these Al concentrations resulted in an increase in primary productivity of the model diatom (). However, the carbon sequestration capacity of the model diatom is varied with different Al concentrations, such that, when it was 20 nmol/L, carbon sequestration capacity of the diatom was higher than when Al was not added. Moreover, Al was taken up by biological behaviors of the diatom and used as a framework element to build siliceous shells. These fundamental results show that Al concentration in seawater has a significant effect on biological activity of diatoms. When the concentration reaches a certain threshold, it significantly decreases the carbon sequestration capacity of diatoms, thus affecting the global role of diatom in the “biological pump.”
Diatoms; Ocean primary productivity; Aluminum; Carbon fixation
Apr. 30, 2020
P735
A
1000-3096(2021)01-0085-07
10.11759/hykx20200430002
2020-04-30;
2020-09-02
国家自然科学基金面上项目(41772041); 国家自然科学基金青年基金项目(41202024); 近海海洋环境科学国家重点实验室(厦门大学)“访问学者基金”(MELRS2006)
[the National Natural Science Foundation of China(General Program), No. 41772041; Youth Foundation of the National Natural Science Foundation of China, No. 41202024; State key Laboratory of Offshore Marine Environmental Science(Xiamen University) “Visiting Scholars Fund”, No. MELRS2006]
周洁玉(1996-), 女, 湖南衡阳人, 硕士生, 主要从事硅藻及环境地球化学的研究, E-mail: zhoujieyu@gig.ac.cn; 刘冬,通信作者, 研究员, E-mail: liudong@gig.ac.cn
(本文编辑: 赵卫红)
