辽东山区典型次生林空间结构对草本物种多样性的影响
2021-01-30刘红民董莉莉高英旭汪成成赵济川
刘红民,董莉莉,高英旭,汪成成,赵济川,王 琴
(辽宁省林业科学研究院,沈阳110032)
生物多样性是指在一定时间和一定地区所有生物(植物、动物、微生物等)及其遗传变异和生态系统的复杂性的总称[1],是人类赖以生存和发展的物质基础[2]。 物种多样性是生物多样性的重要组成部分,能够反映生物群落在组成、结构、功能和动态等方面的异质性[3]。森林生态系统是地球上的生物与其环境相互作用形成的复杂的系统之一,蕴含了陆地生态系统约82%的生物量和50%的生物多样性[4]。 作为森林生态系统基本组成结构的植物群落是维护和调节森林生态系统平衡和改善生态环境的基础, 种群间相互作用和种群与生态环境因子间相互关系决定着森林植被群落的发展进程[5]。 森林植被群落在垂直方向上可以明显划分为乔木层、灌木层和草本层3 层,乔木层是森林生态系统固碳的主体,草本层占据森林生态系统物种丰富度的90%以上。 乔木空间结构直接影响着林下草本物种生长的微环境,决定着草本物种的生长、繁殖和死亡,并在很大程度上决定着林分的稳定性、发展方向和经营措施[6]。 草本植物更新周期短、适应能力强,对林下微环境变化更敏感。 因此,研究乔木层空间结构对林下草本物种多样性的影响具有重要的实践指导意义。
目前,众多学者对林下草本物种的多样性进行了研究。谭一波等[7]在桂西南喀斯特地区研究了土壤、地形和光照等环境因子对蚬木林林下草本植被分布格局的影响机制;朱媛君等[8]针对张北杨树人工林研究了树高、胸径、冠幅、密度、枝下高、死亡率等林分因子对林下草本物种多样性的影响;张维伟等[9]研究了黄土高原桥山林区麻栎林树高、胸径、冠幅、枝下高等林分结构特征与草本层物种多样性相关关系;崔静等[10]采用时空替代法研究了黄土丘陵区人工柠条林年龄结构对林下草本植物物种多样性的影响;杨振奇等[11]采用CCA、PCCA 方法研究了黄土高原砒砂岩区人工沙棘林下草本物种多样性与地形、土壤等环境因子的关系特征。 综合来看,现阶段的研究主要集中在人工林环境因子(地形、土壤、光照等)与非空间结构因子(树高、胸径、冠幅、密度、年龄结构等)对林下草本物种多样性的影响方面,针对次生林林分空间结构因子对林下植被物种多样性的研究较少。辽东山区是我国东北地区重要的天然林区,次生林为其主要的林分类型,合理地经营次生林对于实现辽东山区森林资源的可持续发展具有重要的现实意义。 因此,本研究以辽东山区的栎类纯林、栎类混交林和胡桃楸混交林3 种典型次生林为研究对象,在定量分析林分空间结构参数和物种多样性指数的基础上,利用灰色关联度分析方法探讨了不同类型次生林的林分空间结构与林下草本植物物种多样性之间的关系, 以期为该区域次生林空间结构调控和生物多样性保护提供科学依据。
1 材料与方法
1.1 研究区概况
辽东山区为长白山脉向西南的延伸部分,境内有浑河、太子河、清河和柴河等主要河流,是辽宁中部城市群的绿色屏障和工农业生产的重要水源地,总面积约3.4 万km2。地形属于中低山地,海拔为200~500m,主要山脉为龙岗山脉和千山山脉。属温带季风气候,年平均气温5~8℃,降水主要集中在夏季,年均降水量700~1200mm,无霜期140~160d[12]。土壤多为暗棕色森林土和棕色森林土。本区属长白植物区系,植物种类繁多,生长茂密,地带性森林植物群落——红松(Pinus koraiensis)针阔混交林,在长期的人为干扰后绝大部分已转变为天然次生林,现有天然次生林面积179 万hm2,占辽宁省天然林面积的84%[13]。
1.2 研究方法
1.2.1 样地设置与林分调查 2017 年和2018 年的6~9 月,采用典型取样法在本溪县草河口镇、碱厂镇、草河城镇的栎类次生林、胡桃楸次生林和阔叶混交次生林中,分别设置11 块面积为0.1hm2的圆形调查样地,共计33 块调查样地。 对样地内胸径大于等于5cm 的乔木进行每木检尺,调查乔木树种名称、水平坐标、胸径、树高、冠幅、第一枝下高、健康状况等。 在调查样地内均匀设置3 个1m×1m 草本样方,调查草本物种名称、多度、平均高度、盖度、总盖度等。 并记录经纬度、海拔、坡位、坡度、坡向、郁闭度、林龄、森林类型、土壤类型、腐殖质层厚度等样地基本信息(表1)。

表1 样地基本概况Table 1 Basic information of the sample plots

续表
1.2.2 林分空间结构单元 林木空间结构单元常用4株邻近木法和Voronoi 图法确定,而后者克服了可能将参照树的邻近木排除在外或将非邻近木计算在内的缺陷,近年来被广泛应用。本研究采用Voronoi 图中的相邻多边形确定林分空间结构单元中邻近木的个数n, 经距离缓冲区法(5m 缓冲区)进行边缘矫正后,基于林木位置坐标信息应用ArcGIS 软件生成样地的Voronoi 图(图1)。
1.2.3 林分空间结构参数 本研究采用混交度(Mi)、角尺度(Wi)、开敞度(Bi)、林层指数(Si)、竞争指数(CIi)作为表达林分空间结构的参数。 大小比数仅能从种群的角度来了解某个种在林分中的竞争情况, 而其平均值却不能反映整个林分的竞争程度, 因此本研究并未选择大小比数来表达林分空间结构。 各参数的具体计算公式见表2。
1.2.4 物种多样性指数 本研究采用α多样性测度方法中的Patrick 丰富度指数(S)、Shannon-Wiener 多样性指数(H)、Simpson 优势度指数(D)、Pielou 均匀度指数(J)[14]来分析林下草本物种多样性水平。各参数的具体计算公式见表3。

图1 由样地内林木坐标点构建的Voronoi 图Figure 1 Voronoi diagram constructed from forest tree coordinate points

表2 空间结构参数说明Table 2 Instruction of spatial structure indices

表3 物种多样性指数说明Table 3 Instruction of species diversity index
1.2.5 灰色关联度分析 灰色关联度分析是一种定量化比较分析方法, 根据系统中数据序列的几何相似程度来分析各因素间的关联程度,能够有效地反映系统中各因素的相互影响关系[15]。 本研究将30 个次生林样地林下草本物种多样性指数和空间结构参数作为一个灰色系统,以林下草本的Patrick 丰富度指数、Simpson 优势度指数、Shannon-Wiener 多样性指数、Simpson 优势度指数和Pielou 均匀度指数4 个多样性指数作为参考数列(X0),以角尺度、混交度、开敞度、林层指数和竞争指数5 个空间结构参数作为比较数列(Xi),来分析林分空间结构因子与林下草本物种多样性的关联程度。
关联系数反映参考数列与比较数列在某一时刻的紧密(靠近)程度,其计算公式为:

式中:εi(k)为参考数列与比较数列在k 点时的关联系数;Δi(k)为两个数列在k 点时的绝对差;Δmax 和Δmin 分别表示各个时刻两个数列绝对差中的最大值和最小值,一般取Δmin=0;ρ 为分辨系数,用于增加关联系数之间的差异显著性,减小由最大绝对差引起的失真,其值在区间(0,1),通常取ρ=0.5[16]。
关联度是参考数列与比较数列关联性大小的度量,用各个时刻两个数列关联系数的平均值表示,其计算公式为:

1.3 数据分析
采用Python 3.7 编程语言计算林分空间结构参数和林下草本物种多样性指数。采用SPSS 25.0 软件进行方差分析(α=0.05)和多重比较(LSD 法)。 采用DPS 18.1 软件进行林分空间结构与林下草本物种多样性的灰色关联度分析,采用标准化的方法消除各指标数据的量纲,分辨系数为0.5。 采用Excel 2016 软件进行数据处理和制图。
2 结果与分析
2.1 不同林分空间结构分析
方差分析结果表明,3 种次生林的混交度、林层指数和竞争指数存在显著差异(p<0.05),而角尺度和开敞度差异不显著(p>0.05)。 由图2 可知,3 种林分的混交度排序为:胡桃楸混交林(0.6670)>栎类混交林(0.6014)>栎类纯林(0.2396),胡桃楸混交林和栎类混交林均达到极强度混交,栎类纯林为弱度混交,且混交林和纯林的混交度存在显著性差异(p<0.05);3 种林分的角尺度达到0.3894~0.4189,林木水平分布格局均接近随机分布;3种林分的开敞度达到0.3536~0.3779,林木生长空间均处于基本充足状态;栎类纯林、栎类混交林和胡桃楸混交林的林层指数分别为0.1014,0.1884,0.2601,两种混交林对垂直空间利用程度较好,并且胡桃楸混交林与栎类纯林存在显著性差异(p<0.05);栎类纯林、栎类混交林和胡桃楸混交林的竞争指数分别为2.3594,3.3067,4.3686,混交林林木间的竞争压力明显大于纯林。综合来看,3 种天然次生林的空间结构整体表现为:栎类混交林>胡桃楸混交林>栎类纯林。
2.2 不同林分林下草本物种多样性分析
3 种次生林样地内共发现林下草本植物128 种,分属45 科93 属。 其中,栎类纯林样地内共发现林下草本植物54 种,分属28 科45 属;栎类混交林样地内共发现林下草本植物71 种,分属33 科55 属;胡桃楸混交林样地内共发现林下草本植物69 种,分属33 科60 属。
由图3 可知, 林下草本的Patrick 丰富度指数、Shannon-Wiener 多样性指数、Simpson 优势度指数和Pielou均匀度指数均表现出栎类混交林>胡桃楸混交林>栎类纯林的规律,其中,Patrick 丰富度指数和Shannon-Wiener多样性指数方差分析结果均达到差异显著水平(p<0.05),而Simpson 优势度指数和Pielou 均匀度指数在3 种林分间的差异不显著(p>0.05)。 栎类纯林、栎类混交林和胡桃楸混交林林下草本的Patrick 丰富度指数分别为11.57,18.71,17.57,栎类混交林和胡桃楸混交林林下草本的物种丰富度显著高于栎类纯林,分别高于栎类纯林的1.6 倍和1.5 倍。 栎类纯林、 栎类混交林和胡桃楸混交林林下草本的Shannon-Wiener 多样性指数分别为1.9301,2.4382,2.1396,栎类混交林与栎类纯林存在显著性差异(p<0.05)。
2.3 林分空间结构与草本物种多样性的灰色关联度分析
为进一步探讨林分空间结构对林下草本物种多样性的影响,运用DPS 18.1 数据处理系统进行林分空间结构参数与林下草本物种多样性指数的灰色关联度分析。 由表4 可知,对于栎类纯林,角尺度与Shannon-Wiener多样性指数、Simpson 优势度指数、Pielou 均匀度指数的关联程度最大,混交度与Patrick 丰富度指数的关联程度最大,说明对栎类纯林林下草本物种多样性影响最大的空间结构参数是角尺度,其次是混交度;对于栎类混交林,混交度与Simpson 优势度指数、Pielou 均匀度指数的关联程度最大,角尺度与Patrick 丰富度指数的关联程度最大,开敞度与Shannon-Wiener 多样性指数的关联程度最大,说明对栎类混交林林下草本物种多样性影响最大的空间结构参数是混交度, 其次是角尺度和开敞度; 对于胡桃楸混交林, 混交度与Patrick 丰富度指数、Shannon-Wiener 多样性指数的关联程度最大,开敞度与Simpson 优势度指数、Pielou 均匀度指数的关联程度最大,说明对胡桃楸混交林林下草本物种多样性影响较大的空间结构参数是混交度和开敞度。 可以看出,3 种林分的空间结构参数与林下草本物种多样性指数的灰色关联度均较高,同一空间结构参数与不同多样性指数的关联序存在差异,说明草本层物种多样性受多个空间结构参数指数共同影响。 总体而言,林分混交度是影响3种林分林下草本物种多样性的共同关键因子, 混交度和开敞度是影响两种混交林林下草本物种多样性的关键因子。

图2 空间结构参数Figure 2 Stand spatial structure parameter
3 讨论与结论
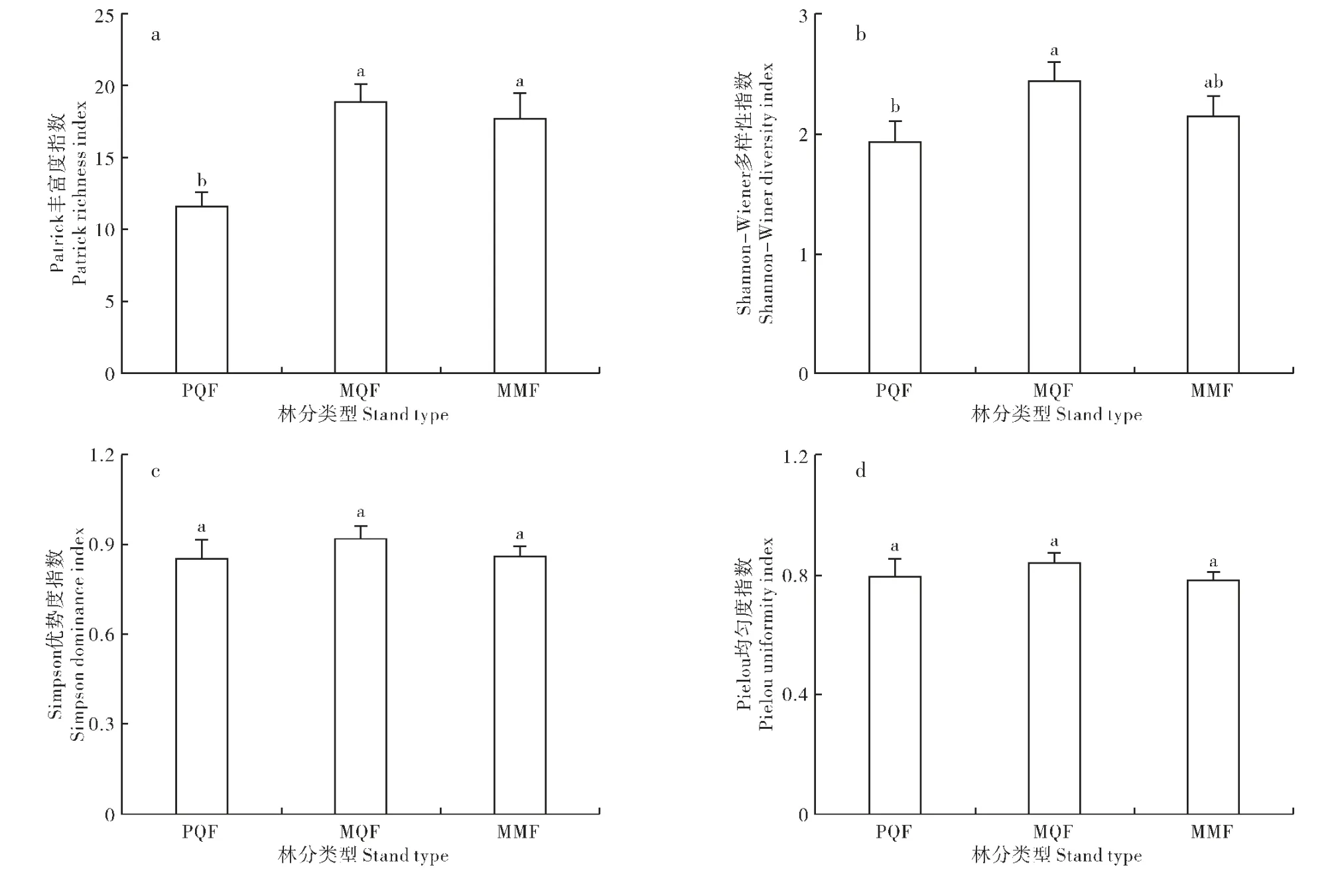
图3 林下草本物种多样性指数Figure 3 Understory herbaceous species diversity index
原始林是森林生物与环境相互依存和相互作用、长期协同进化的产物,次生林可以理解为是原始森林生态系统的一种退化,由于生态系统的基本结构和固有功能的破坏或丧失,导致生物多样性下降,稳定性和抗逆能力减弱,系统生产力降低等[17]。 次生林经营就是通过合理的人为措施使次生林的发展既符合自然演替规律,又能满足人类的生态和经济效益需求。 林分空间结构决定了林木之间的竞争势以及空间生态位,合理的林分空间结构是确保森林生态系统健康发展的关键因素[18],因此,要实现培育多功能森林的目标,就必须了解林分空间结构的现状,通过调整建立或者维护最佳的林分空间结构。 研究区的栎类纯林、栎类混交林和胡桃楸混交林3 种天然次生林在树种隔离程度(混交度)、林层多样性(林层指数)、林木竞争强度(竞争指数)3 个方面存在显著差异,总体来看,混交林的空间结构优于纯林,这与王伟平等[19-21]的研究结果一致。 坡位是在小尺度上影响森林植被分布及其结构的主要地形因子之一,下坡位林分空间结构在很大程度上要优于上坡位[22]。 本研究中的两种混交林类型相比,胡桃楸混交林的树种隔离程度、林层多样性和林木竞争强度表现较好,这可能是因为胡桃楸混交林主要分布在立地条件较好的下坡位,而栎类混交林主要分布在立地条件较差的中、上坡位[23]。 加之栎类作为辽东山区主要用材树种之一,历史上遭到严重的破坏,并且栎类次生林长期以来一直被作为改造的对象,不合理的人为干扰较为严重,导致种群结构处于退化状态[24]。
不同森林类型由于在树种组成、空间结构等方面的不同,造成了林内的微环境(光照、温度、水分等)、土壤性质和凋落物性质等林分立地条件的差异,并对林下植被的物种组成、分布格局和生长发育产生间接影响[25]。研究区3 种天然次生林林下草本的Patrick 丰富度指数和Shannon-Wiener 多样性指数存在显著差异, 胡桃楸混交林和栎类混交林较优,栎类纯林较差。混交林较纯林相比,其林下枯枝落叶物丰富且质量高,微生物活动频繁,养分分解能力及转化速率加快,土壤结构改善,土壤肥力和持水性能提升,有利于林下植物的生长和发育[26]。同时,混交林水平结构和垂直结构复杂,林冠呈多层镶嵌郁闭,林内光斑组成和光照强度多样化,存在光合作用的空间匹配效应,显著提升林下植被的光能利用效率,进而提高了林下草本植物的物种多样性[27]。 而栎类纯林的上林层比重较大,群落结构相对简单,林下光照条件在时空上分布差异不明显,并且栎类纯林的立地条件比较严酷,土壤贫瘠干旱[28],因而造成林下草本生物多样性偏低。
林分空间结构决定了种群内树木间的空间排列方式以及对周边环境资源的占用能力,会对林分的生长、稳定性及生物多样性等产生影响[29]。 体现林分内树种隔离程度的混交度和体现树木个体水平分布格局的角尺度,会对林下枯落物分布以及林内光斑分布造成影响,导致林下植被多样性特征出现差异[30]。 开敞度能够直观反映林木生长空间的大小,是体现林下个体所处的光环境情况的一种测量指标,林下植被多样性表现为开敞度基本充足的林分高于开敞度严重不足的林分[31]。 在本研究中,林分混交度与林下草本物种多样性表现出较为明显的关联性,角尺度和开敞度也在不同林分类型与林下草本物种多样性存在不同程度的关联性,较好地说明了林分的空间结构与林下草本的物种多样性具有明显的关系。 因此,在辽东山区次生林经营管理过程中,可以选择以调整树种结构为主,调整林木的水平和垂直空间分布格局为辅的林分空间结构优化方案,有利于提高林下草本植物的物种多样性。

表4 林下草本植被物种多样性与林分空间结构关联度及其排序Table 4 Correlation and sort of the understory herbaceous species and stand spatial structure
灰色关联度分析具有样本要求少、计算量小、量化结果与定性分析一致等优点,但其评价结果只能给出评价对象的优劣次序,无法评估影响其优劣的内部因素。同时,参数序列的选择不同也会对评价结果造成偏差。因此,本研究还需加大数据调查量,在此基础上结合回归分析、相关分析等统计方法,进一步揭示辽东山区天然次生林的林分空间结构与林下植被物种多样性之间是否存在线性或非线性关系。
