基于组成型抗性性状的油茶抗茶籽象的评价模型*
2021-01-25张守科方林鑫舒金平汪阳东王浩杰
张守科 方林鑫 王 毅 张 威 舒金平 汪阳东 王浩杰
(1. 中国林业科学研究院亚热带林业研究所 杭州 311400; 2. 中国林业科学研究院林木遗传育种国家重点实验室 北京 100091; 3. 青田县林业局 青田 323900)
长期以来,病虫害是造成作物损伤乃至减产的关键因素,培育并推广抗病虫品种是治理病虫害最根本、最经济有效的手段,抗病虫育种一直是育种研究的重要内容(吴元奇, 2001)。构建有效的抗病虫评价模型以评估不同品种的抗病虫能力,对于新品种的选育和应用有着重要意义。
育种工作中品种表型稳定性是评价品种优良程度的重要指标。植物抗性分为诱导性抗性和组成型抗性两大类(张蓬军, 2007; 李新岗等, 2008; 谢辉等, 2012),抗病虫品种表型的筛选往往聚焦于植物组成型抗性性状。组成型抗性是植物固有的、基因决定的包括植物自身形态结构和生理生化等方面的抗性(Karbanetal., 1997; 桂连友等, 2004)。植物组成型抗性主要受基因控制,不因一时环境因素改变而影响其抗性品质,在植物寄主体内始终发挥作用(孟昭军, 2008),在抗性品种选育中更具优势。植物抗虫不单单靠一个防御系统,有时是多系统联用来抵御害虫。植物在植食性昆虫危害的选择压下进化出各种抗性策略,表现为化学性状(驱避性挥发物、有毒次生代谢物质积累等)及物理性状(果皮/叶片厚度、硬度等)的改变(Myersetal., 2017),而植食性昆虫在此进程中与植物形成了“军备竞赛”式的协同进化关系(Tojuetal., 2011)。因而,植物抗性评价模型的构建需要考虑多抗性指标联合才可提升模型的精度和准度(吴元奇, 2001)。
油茶(Camelliaoleifera)是我国目前主栽的木本油料作物之一,在南方山地经济发展中占据重要地位。因其产油率高、油品质量好等因素与油橄榄(Oleaeuropaea)、油棕(Elaeisguineensis)、椰子(Cocosnucifera)并称为世界四大木本油料(姚小华等, 2012)。随着我国油茶产业的迅猛发展,油茶种植面积不断扩大,集约程度不断提高,油茶病虫害也随之发生,其中茶籽象(Curculiochinensis)(鞘翅目Coleoptera: 象甲科Curculionidae)最为严重。茶籽象是油茶最为主要的种实害虫,以成虫和幼虫危害茶果,造成大量落果,严重影响茶果产量和茶油品质。茶籽象危害隐蔽,一旦错过成虫羽化产卵的关键防治期,防治十分困难(舒金平等, 2013; 何立红等, 2014; 张守科等, 2019)。当前,茶籽象的防治仍依赖化学防治,但农药防治效果差而且有农药残留问题,食品及生态安全风险大,因此,筛选抗虫性好、产油率高的无性系极为迫切。近年来,山茶属(Camellia)植物抗病虫研究仅茶树(C.sinensis)开展了一些诱导性抗性研究(Zhuetal., 2019),对于组成型抗性的研究较少(李荣林等, 2015; 赵丰华等, 2015; 金珊等, 2016; 钟明跃, 2018),而对于油茶抗虫组成型抗性性状的研究尚未开展,以至于对油茶不同品种的抗虫性评价尚未有统一标准(何立红等, 2014; Zhangetal., 2020),因此构建基于多个组成型抗性性状参与的抗性评价模型对于油茶抗性育种有着重要的参考价值。
本研究以山茶属植物抗茶籽象研究试验验证的3种抗性指标(果形指数、果皮厚度、关键时期茶皂素积累量)作为依据(Zhangetal., 2020),以普通油茶无性系作为研究对象,长时间尺度调查茶籽象为害率,通过逐步回归分析,确定普通油茶抗茶籽象关键组成抗性性状,并初步构建抗性评价模型,为油茶新品种抗茶籽象与否提供判断依据,并为我国油茶抗虫育种提供新思路。
1 材料与方法
1.1 试验地及油茶试验林概况
试验地分别位于江西省新余市分宜县长埠实验林场油茶种质园(27°57′28″N,115°04′11″E,海拔156 m)、浙江省青田大尖山油茶种植园(28°27′32″N,120°16′48″E,海拔432 m)。2处试验地均属于亚热带季风气候,均为油茶纯林,油茶林基本处于无人为喷施农药防治病、虫、杂草的自然状态,林下分布芒萁(Dicranopterispedata)、杠板归(Polygonumperfoliatum)及禾本科(Poaceae)狗牙根(Cynodondactylon)、白茅(Imperatacylindrica)等杂草。长埠实验林场油茶种质园油茶树龄15 ~ 20年,种植长林系列多个无性系且无性系间混种,均有结实,周围有柑橘(Citrusreticulata)林分布。大尖山油茶种植园油茶树龄10 ~ 15年,主栽长林系列多个无性系且无性系间混种,均有结实,周围无其他树种分布。自2016年,本研究选取我国主栽的长林系列普通油茶8个高产优质无性系(长林3号、长林4号、长林18号、长林22号、长林40号、长林53号、长林55号、长林166号)作为抗性评价模型构建的样本无性系,连续4年不间断观测其生长状况、结实规律及茶籽象危害情况。
1.2 茶籽象为害率调查
于2016—2019年,每年9月下旬茶果采摘前,对选定的油茶8个无性系进行随机采样。每一个无性系随机选择5株样株,每株分为上、中、下3个层次,东、南、西、北4个方位,每个层次每个方位随机摘取20个茶果,每株共计240个茶果,不足240个的植株则全部采摘。将2处搜集的茶果按照无性系编号带回杭州市中国林业科学研究院亚热带林业研究所养虫室,放置于室温环境下,任老熟幼虫自由逸出茶果,统计老熟幼虫数量。待11月底,如持续7天未记录到幼虫逸出,则认定该无性系茶籽象已经完全老熟逸出。按照为害率公式(李苗苗等, 2016): 为害率(%)=(茶籽象逸出孔数/油茶果个数)× 100%,统计不同无性系的受害率。
1.3 果形指数及果皮厚度测定
于2016—2019年,每年5月下旬茶籽象产卵高峰期时,在2块试验地内采集茶果,采样方法同1.2。将采集的幼嫩茶果使用游标卡尺测定果长(a)、果宽(b)、果高(L),按照公式: 果形指数=(a+b)/2L,分别求得8个油茶无性系果形指数(李青等, 2012; 何立红等, 2014)。果皮厚度的测定按照果实基部、中部、端部3处厚度平均值作为一个茶果的果皮厚度。每个无性系随机选取5株,每株240个果子,共测定1 200个茶果。
1.4 茶籽象发育关键期油茶种子中茶皂素含量测定
1.4.1 样品准备 基于前期对茶籽象对长江中下游地区山茶属(Camellia)植物的危害规律的调查(Zhangetal., 2020),山茶属植物种子茶皂素含量变化与茶籽象幼虫发育时间密切相关,茶籽象幼虫的发育可分为产卵高峰期(5月下旬—6月上旬)、初孵幼虫期(6月中旬—6月下旬)、幼虫发育期(7月上旬— 8月中旬)和幼虫成熟期(8月下旬— 9月下旬)4个阶段。2016—2019年,在这4个关键时期分别进行茶果样品采集,采集方法同1.2,但每个层次每个方位仅随机摘取5个茶果,每株共计60个茶果; 将茶果放置于4 ℃冰箱内,带回实验室后将种仁剥出置于 -20 ℃冰箱内冷冻保存供试。每个无性系随机选取5株,每株60个茶果的种仁作为一个样品,共5个重复。
1.4.2 茶皂素含量测定 参考Zhang(2016)的方法测定茶皂素含量。色谱预处理条件如下: 色谱柱为安捷伦Eclipse XDB-C18(安捷伦科技公司)(4.6 mm × 250 mm,5 μm),流动相为甲醇∶水(体积比9∶1); 检测波长为210 nm; 柱温25 ℃。标准处理: 准确称量茶皂素标准品0.5 g(精确至0.000 1 g),用甲醇超声溶解,体积为100 mL,分别取1.00、3.00、5.00、7.00、9.00 mL,置于5 个50 mL体积的容量瓶中。0.45 μm微孔膜过滤。标准曲线以质量浓度作为横坐标和对应的峰面积作为纵坐标计算绘制。样品中茶皂素含量测定: 称取样品0.058 g(精确至0.000 1 g),用甲醇超声溶解,定容于50 mL容量瓶中。0.45 μm微孔膜过滤。每个样品5个重复。按公式计算茶皂素含量,茶皂素含量(%)=X×V/m× 100%,式中:X为样品溶液浓度(g·mL-1),V为定容体积(mL),m为样品质量(g)。
1.5 数据统计分析及模型构建
利用SPSS 20软件对8个油茶无性系间茶籽象为害率、果形指数、果皮厚度及4个时期茶皂素含量等数据进行ANOVA单因素方差分析。并利用Graphpad绘图软件绘制小提琴散点图及进行方差分析。基于果形指数数据,利用R语言kmean()函数进行8个无性系聚类分析(汤银才, 2008); 并利用R语言中ggplot2包中geom_ribbon()函数绘制基于果形指数分布的峰峦图直观展示8个无性系果形指数分布状况及聚类情况。利用网络在线绘图工具(https:∥www.omicshare.com/tools/Home/Soft/heatmap)绘制不同时期茶皂素含量变化热图(Zhangetal., 2020)。回归模型的构建采用R语言中MASS包中的stepAIC()函数(汪海波等, 2018),利用果形指数(X1)、果皮厚度(X2)、4个时期的茶皂素含量(X3-6)、茶籽象为害率(Y),依据精准AIC准则进行逐步回归分析,筛选出与Y最为相关的X值带入模型。
2 结果与分析
2.1 茶籽象对8个油茶无性系的为害率
4年间对油茶长林系列8个主推无性系茶籽象为害率的追踪调查发现,长林3号(46.02% ± 0.02%)和长林18号(37.30% ± 0.02%)4年受害率维持较高(图1); 而长林166号受害率为35.33% ± 0.01%,在选取的8个无性系中处于中间类型; 其余长林4号、22号、40号、53号及55号彼此之间受害率差异不大(P>0.05),维持在20%左右,与长林3号、18号、166号差异显著(P<0.05)(图 1)。4年内长林22号的受害率极大值(30%)与极小值(12%)差异最大,长林55号极值变异最小(图 1)。
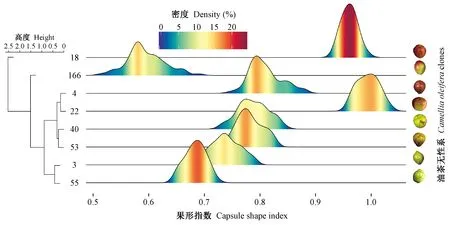
图2 油茶8个无性系的果型指数差异Fig.2 The difference of capsule-shape indexes among 8 clones of Camellia oleifera
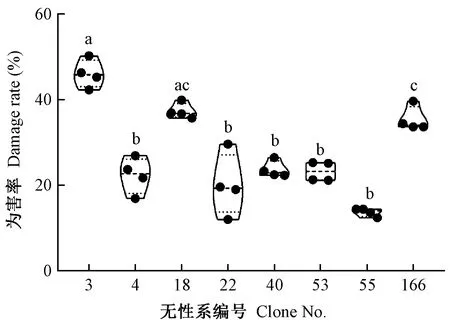
图1 茶籽象对油茶8个无性系的为害率Fig.1 Damage rate of Curculio chinensis to 8 clones of Camellia oleifera不同小写字母表示差异显著(P <0.05)。下同。The bar marked with different lowercase letters is significantly different at the level of P<0.05. The same below.
2.2 茶籽象产卵高峰期8个油茶无性系的果形指数
茶籽象产卵高峰期,8个油茶无性系的幼果果形指数存在显著差异,差异主要来自无性系分组组间(F=1 766.98,P<0.001)。从聚类树来看,可以分为4个大的类群: 长林3号和55号; 长林4号、22号、40号和53号; 长林166号; 长林18号(图 2)。其中,长林18号和22号果形指数接近于1,幼果近圆形; 且长林18号幼果发育更为统一,大多数幼果果形指数差异不大,密集聚集于0.9 ~ 1之间,极值之差为0.06(图 2); 而长林22号果形指数极大值1.04,极小值0.96,且较为均匀地分布于0.96~1.04之间(图 2)。8个无性系中,长林166号的果形指数变异最大,极值之差为0.18,所有果形指数处于0.51 ~ 0.69之间,果型为椭圆形。其他无性系介于长林166号与长林18号和22号之间,果形指数差异不大(P>0.05)(图 2)。
2.3 茶籽象产卵高峰期8个油茶无性系的果皮厚度
茶籽象产卵高峰期,8个油茶无性系果皮厚度差异显著,差异主要来自无性系分组组间(F=198.92,P<0.001)。其中长林53号果皮最厚,为(5.03 ± 0.08)mm,长林22号次之,为(4.095 ± 0.07)mm(图 3); 长林3号、4号、18号和22号果皮厚度均值均处于4 ~ 5 mm之间,4者存在差异,但仅长林4号与长林22号存在显著差异(P<0.05),而从无性系内部变异分析来看,长林18号和22号无性系内极值差(分别为0.18和0.2)均大于长林3号和4号(分别为0.03和0.02)(图 3); 长林55号和166号,在茶籽象产卵高峰期平均果皮厚度最薄,处于3 ~ 4 mm,且无性系内极值差也不大,但二者果皮厚度差异显著(P<0.05)(图3)。

图3 油茶8个无性系的果皮厚度差异Fig.3 The difference of pericarp thickness among 8 clones of Camellia oleifera
2.4 茶籽象发育过程中8个油茶无性系种子中茶皂素积累规律
对茶籽象产卵高峰期、初孵幼虫期、幼虫发育期、幼虫成熟期油茶种子中茶皂素积累量进行聚类分析,结果表明,长林22号、40号和55号聚为一支; 长林3号和4号聚为一支; 长林18号、53号和166号聚为一支(图 4)。4个时期,8个无性系之间茶皂素含量差异都极显著(P<0.001),且变异主要来自组间,说明茶皂素含量无性系之间的差异大于无性系内(表 1)。从发育期来看,产卵高峰期长林3号、4号、18号、53号及166号茶皂素含量差异不大(P<0.05),22号和40号则显著高于其他无性系(P<0.05),分别为(3.5 ± 0.35)mg·g-1和(3.4 ± 0.096)mg·g-1; 初孵幼虫期,长林3号、4号、18号和166号茶皂素含量差异不大(P>0.05),与长林22号、53号及55号差异显著(P<0.05); 幼虫发育期,长林3号和4号茶皂素含量最低且与其他样本差异显著(P<0.05),长林40号含量则最高; 幼虫成熟期,长林22号、40号、55号茶皂素含量最高,长林3号、4号最低,长林18号、53号、166号居于中间,且这3组无性系之间的含量差异显著(P<0.05)(图 4)。

图4 油茶8个无性系种子中茶皂素积累规律Fig.4 The accumulation of tea saponin content in seeds among 8 clones of Camellia oleifera
2.5 基于8个油茶无性系的组成型抗性指标构建抗性评价模型
以茶籽象为害率(Y)为因变量,运算开始时模型包含6个自变量(X1: 果形指数;X2: 果皮厚度;X3: 产卵高峰期茶皂素含量;X4: 初孵幼虫期茶皂素含量;X5: 幼虫发育期茶皂素含量;X6: 幼虫成熟期茶皂素含量),每一次拟合是依据贡献值排除1个变量,初始AIC=-136.53,随着变量的依次排除,AIC值发生由-160.21、-163.67、-173.5、-175.86的变化; 且果形指数(X1,P=0.216 9)、果皮厚度(X2,P<0.001)、4个时期的茶皂素积累量(X3,P=0.129 9;X4,P<0.001;X5,P<0.05;X6,P<0.01)6个自变量,模型检验结果拟合度较好。但经过偏回归检验分析: 果皮厚度(X2,P=0.072 99)、茶籽象发育后3个时期的茶皂素积累量(X4,P<0.001;X5,P<0.01;X6,P<0.01)4个变量与0有显著差异,其余变量与0没有显著差异。最终基于4个变量进行逐步回归,构建模型为:Y=0.531 6-0.301X2- 0.436 57X4+ 0.059 526X5- 0.019 4X6,拟合程度好(R2=0.773 7,P<0.01)。
3 讨论
林木抗虫育种研究起步晚,且受限于遗传转化体系难构建、育种周期长的劣势,研究进展一直相对缓慢(万志兵等, 2012)。在国家倡导生态文明建设推动经济林产业发展的形势下,林木抗虫品种的选育仍然是现阶段林木遗传育种研究的重要内容。选育稳定遗传的组成型抗虫表型是抗虫育种的关键(吴元奇, 2001)。植物为了适应植食性昆虫危害的选择压力进化出各种组成型抗性性状,表现为化学性状(拒食性挥发物、各种有毒次生代谢物质稳定积累)及物理性状(厚度、硬度)的改变(Myersetal., 2017)。
山茶属植物组成型抗性包括果皮厚度、果形指数、果实绒毛、颜色等物理指标(李荣林等, 2015; 赵丰华等, 2015; 金珊等, 2016)和茶皂素、单宁酸含量等化学指标,可以作为山茶属植物重点的抗性性状加以筛选(Zhangetal., 2020)。山茶属植物果实多呈圆球状,针对不同无性系油茶果实测定的果形指数介于0.5 ~ 1.4之间(左继林等, 2012; 董万领等, 2013; 李小梅等, 2013),呈现椭圆形到圆形。何立红等(2014)在对小果油茶(Camelliameiocarpa)果形指数与茶籽象为害率的研究中发现,果形指数越接近于1即果子越圆为害率越高。本研究对油茶8个长林无性系的果形指数测定基本与何立红等(2014)结果一致,无性系之间果型变异较大(果形指数在0.5 ~ 1.2之间),但在多元线性回归分析中果形指数拟合度不佳,且偏回归系数与0差异不显著(P=0.769 4),构建模型时被舍去。茶籽象幼虫无转寄主特性,不同品种油茶受害率与茶籽象成虫产卵选择密切相关,因此成虫产卵期的果形指数可能与茶籽象产卵偏好有关。本研究测定了茶籽象产卵期不同品种的果形指数,但其在回归拟合时被舍去,说明在茶籽象产卵高峰期果形指数在油茶抗性评价中贡献不显著(X1,P=0.216 9)。何立红等(2014)认为果形指数与茶籽象危害密切相关,其测定的是幼虫老熟期茶果的尺寸,而本研究是成虫产卵期的果形指数,结果不一致的原因可能与取样时间不一致有关。
植物表皮组织加厚,是抵御昆虫危害的一种适应机制。不同茶树(Camelliasinensis)种质对黑刺粉虱(Aleurocanthusspiniferus)的抗性随着叶片海绵组织增厚而表现抗性增强(王庆森等, 2009); 沙棘(Hippophaerhamnoides)树皮表皮厚度、表皮细胞紧密程度等与抗沙棘木蠹蛾(Holcocerushippophaecolus)有显著相关性(赵延霞等, 2012); 在日本山茶(Camelliajaponica)与日本山茶象(Curculiocamelliae)协同进化关系的研究中也发现类似规律,象甲喙的长度与茶果果皮厚度关系密切(Tojuetal., 2005; 2006; 2011; Toju, 2011)。本课题组的前期研究发现,茶籽象喙长约5 ~ 7 mm,而除浙江红山茶(Camelliachekiangoleosa)(果皮厚度 >8 mm)外大部分山茶属植物的果皮厚度都在6 mm以下,且我国各地区茶籽象并未形成基于寄主适应的局域种群(Zhangetal., 2018; 2019),因此基于最优觅食理论(Charnov, 1976)及山茶属植物果皮厚度与茶籽象为害率的关系分析推测,果皮厚度可能是阻碍茶籽象产卵的重要屏障,果皮厚度增加了雌性茶籽象的取食、产卵难度。从本研究中的8个油茶无性系茶籽象产卵高峰期果皮厚度测定结果来看,不同无性系之间果皮厚度存在明显差异,且多元线性回归模型中果皮厚度拟合度高且显著(P<0.001),但偏回归系数与0差异显著性较低(P=0.769 4),果皮厚度在模型中贡献权重仅次于初孵幼虫期茶皂素含量。
植物能稳定表达产生对害虫有毒、抗营养和难消化的次生代谢化合物直接杀伤昆虫或降低昆虫对食物的消化能力,使昆虫不能获得足够的营养,从而可减少或阻止昆虫的进一步取食,达到直接抗虫的目的。植物皂苷是一类重要的防御性化合物,包含三萜类和甾体类皂苷。研究表明茶皂苷会刺激小鼠(Musmusculus)肠道黏膜引起毒性病变(Kawaguchietal.,1994),甾体类皂苷则会引发羊的肠胃疾病,从而降低肠道蠕动,降低食物利用效率导致植食性动物体质量下降(Wisløffetal., 2008)。山茶属植物可以通过茶皂素抑制茶籽象与营养代谢相关的肠道菌群丰度,进而造成茶籽象发育不良,来发挥抗虫作用(Zhangetal., 2020)。油茶8个长林无性系种子中的茶皂素在茶籽象4个发育关键时期的积累规律存在明显差异,其中初孵幼虫期、幼虫发育期和幼虫成熟期茶皂素含量在多元线性回归模型中占据较高权重且拟合显著(P<0.001)。产卵高峰期茶皂素浓度在模型拟合过程中由于拟合度不高被舍弃,说明产卵高峰期油茶无性系种子中茶皂素含量低且对雌性茶籽象产卵影响不大,与后期为害率相关性不大。
茶籽象危害是导致茶果采前脱落的重要原因(舒金平等, 2013),但茶籽象危害导致落果的多少与品种、种植年限、种植条件及气候等因素均有关系。本研究发现9月下旬在选定的2个试验样地内还未出现明显的虫害落果现象和幼虫逃逸现象,这可能与本研究中调查的对象均是栽种时间不长的新品种林且种植及管理情况良好有关。但在落果较早且较为严重的油茶种植区,选择7月中、下旬进行茶籽象为害率的调查可能更为准确。
4 结论
茶籽象是我国油茶产业健康发展的重要威胁性虫害。本研究基于油茶长林无性系组成型抗性性状测定,并采用多元线性回归模型综合分析果形指数和果皮厚度以及茶籽象4个为害关键期油茶种子中茶皂素含量与茶籽象为害率的关系,初步构建了以茶籽象为害率为因变量、以果皮厚度及茶籽象初孵幼虫期、幼虫发育期和幼虫成熟期油茶种子中茶皂素含量为自变量的油茶抗茶籽象评价模型。模型的构建可为油茶新品种对茶籽象的抗性评价提供依据,为我国油茶品种的抗病虫选育提供新思路。
