红色荧光蛋白基因DsRED在核桃植株再生过程中的表达稳定性*
2021-01-25张佳琦胡恒康黄有军娄和强张启香
任 飞 张佳琦 胡恒康 梁 璧 黄有军 娄和强 张启香
(浙江农林大学林业与生物技术学院 省部共建亚热带森林培育国家重点实验室 杭州 311300)
报告基因是编码易被检测的蛋白质或酶的基因。在含有或被转入报告基因的细胞内,报告基因的表达产物可以直接产生信号或通过特定的酶促反应催化底物间接产生信号(Jawedetal., 1990)。相比于内源性生色团、化学染料和纳米探针,报告基因的编码产物具有稳定性好、特异性强及适用范围广等优点(李登峰等, 2018)。目前,在转基因植物中,使用最为广泛的为β-葡糖苷酸酶(GUS)。据不完全统计,至今已获得了上千种融合GUS报告基因的转基因植物(Honmaetal., 2019)。其优点为检测方法简单、灵敏度高,然而,由于其不能进行活体染色,组织化学染色后试验材料失去继续培养的可能,在检测植物茎秆和叶片等时会对材料造成很大的伤害和损失(李新锋等, 2016)。
荧光蛋白家族是从水螅虫纲(Hydrozoa)和珊瑚虫纲(Anthozoa)动物中分离出的一类同源性蛋白,由荧光蛋白基因编码,包括绿色、红色、黄色和青色荧光蛋白等,在细胞内蛋白质的定位以及动态示踪等方面有着广泛的应用(杨宇等, 2011)。相较于其他分子标记,荧光蛋白更加易于与目的基因蛋白融合,对活细胞毒性小,应用更加便利(王文旭等, 2019)。目前应用最为广泛的绿色荧光蛋白(green fluorescent protein, GFP)是从维多利亚多管发光水母(Aequoreavictoria)中分离所得,由于其可以自发荧光,无需辅助因子和底物,无毒性,因此适用于监测活细胞或生物体中蛋白质的动态变化(吴瑞等, 2005)。GFP通过一系列体外分子进化,发展出了多种荧光波长不同、发光强度极高的荧光蛋白,如增强型绿色荧光蛋白(EGFP)、蓝色荧光蛋白(BFP)、青色荧光蛋白(CFP)等。但这些荧光蛋白的发射波长均在440~529 nm之间,极易激发胞内物质产生荧光,导致成像时背景较高,在一定程度上限制了这些荧光蛋白的应用。
红色荧光蛋白(red fluorescent protein, RFP)是从香菇珊瑚(Discosomasp.)中提取的一种与绿色荧光蛋白同源的生物发光蛋白(Alfordetal., 2012)。Clontech公司将其进行低毒、低寡聚化处理后形成突变体DsRED,其激发波长和发射波长均较长,具有对动植物组织损伤小、光漂白作用低等优点(Yanushevichetal., 2002),目前已被广泛用于动植物以及酵母等真核细胞内基因表达的报告基因(Yarbroughetal., 2001; Jachetal., 2001; Czymmeketal., 2002; 刘娜等, 2005; Zhangetal., 2015)。然而,尽管DsRED已作为报告基因转入多种植物,但成功获得再生植株的案例报道不多(高嵩等, 2017)。
目前,探究外源基因在植物中表达的稳定性和时空差异等研究多以当年再生植株为试验材料(陈盼飞等, 2016),多年生林木方面的研究较少,尤其果树该方面的研究更为鲜见(王倩倩等, 2013)。在果树转基因研究中,番木瓜(Caricapapaya)、苹果(Malusdomestica)、核桃(Juglansregia)等的研究比较深入。在核桃中,较早转化的外源基因如GUS基因、卡那霉素抗性基因(nptⅡ)、苏云金芽孢杆菌基因[BtcryIA(c)]、查耳酮合成反义基因(CHS)等(Mcgranahanetal., 1990; Dandekaretal., 1998; El-Euchetal., 1998; 汤浩茹等, 2000),已转入核桃并得到表达,但外源基因在核桃中表达的后续报道较少。本研究以前期通过根癌农杆菌(Agrobacteriumtumefaciens)介导法获得的携带外源报告基因DsRED的体细胞胚(简称“体胚”)和体胚再生植株(Zhangetal., 2015)为研究对象,利用荧光显微镜和PCR、qRT-PCR、Western Blot等技术检测DsRED基因在核桃体胚、组培苗以及温室3年生苗中表达的稳定性,以探讨红色荧光蛋白基因DsRED作为外源报告基因能否在核桃再生植株中稳定表达,及其对核桃再生植株生长发育的影响,为进一步拓宽DsRED作为报告基因的应用范围以及开展果树嫁接专用砧木的分子辅助育种提供参考。
1 材料与方法
1.1 植物材料 植物材料来源于课题组2015年获得、2017年再次经荧光和PCR检测为DsRED阳性的核桃体胚和再生植株(Zhangetal., 2015; Liuetal., 2017)。 其中DsRED基因为pKGW-RR载体所携带,该pKGW-RR载体包含拟南芥(Arabidopsisthaliana)的组成型泛素启动子和卡那霉素抗性基因,载体结构见Smit等(2005)。本研究所用组培苗为同一无性系DsRED体胚的子叶胚于2017年经脱水萌发获得的再生植株增殖并生根获得; 3年生苗则是将上述组培苗于2017年经生根、炼苗、驯化并定植于温室,培养生长3年。
1.2 试验方法 1)培养条件 核桃体胚增殖培养、植株再生以及组培苗增殖、生根培养等的培养条件见Zhang等(2015),光照时间每天16 h,光照强度1 500~2 000 lx,培养温度(25±2)℃。体胚每周继代培养1次,组培苗每2周继代培养1次。3年生苗培养于浙江农林大学智能温室中,常规管理。
2)临时切片制作 分别取组培苗和3年生苗的根、茎和小叶,用5%(W/V)的琼脂糖进行包埋,冷却凝固后,切割修理组织块。将组织块固定到振动切片机VT1200(Leica公司)底座上,设定切片厚度100 μm后对组织进行切片。并将切片置于载玻片上制成临时切片,显微镜下观察并拍照。每样品选10个植株,生物学重复3次(下同)。
3)荧光检测 取核桃DsRED体胚、组培苗的根、茎、小叶以及3年生苗的根和小叶(包括对照体胚、对照组培苗以及对照3年生苗的相应结构),置于体视显微镜SteREO Discovery V12(Zeiss 公司)下,分别在明场和荧光光照(激发波长540 nm)条件下利用ZEN lite成像软件进行连续拍照。
将制作的核桃DsRED组培苗的根、茎和小叶横切面以及3年生苗的根和小叶横切面临时切片(含对照),放置于正置荧光显微镜BX60(OLYMPUS公司)下,分别在明场和荧光光照(激发波长540 nm)条件下利用DP Controller成像软件对各组织切片进行连续拍照。
4)茎叶生长参数测定 对DsRED组培苗和3年生苗复叶的小叶数量进行统计; 用直尺分别测量组培苗和3年生苗的株高、株幅、茎粗、单叶长、单叶宽,并用扫描仪V330(EPSON公司)对每片单叶扫描成像,通过Image J软件计算叶面积。
5)DsRED基因检测 以CTAB法分别提取DsRED体胚、组培苗的根、茎、小叶(包括对照体胚、对照组培苗)的DNA,并进行PCR检测。引物由上海生工生物工程公司合成,正反向引物序列为F: 5′-ACGATGGTGTAGTCCTCGTTGT-3′; R: 5′-CTCC TCCAAGAACGTCATCAAG-3′。PCR扩增程序为: 95 ℃预变性4 min; 95 ℃ 30 s,62 ℃ 30 s,72 ℃ 30 s,共30个循环; 72 ℃延伸4 min。最后1% TAE琼脂糖凝胶电泳中若出现 681 bp大小的条带,则为阳性。
6)DsREDmRNA的检测 用RNAprep Pure多糖多酚植物总RNA提取试剂盒(DP441,天根),分别提取DsRED体胚、组培苗的根、茎、小叶(包括对照体胚、对照组培苗)总RNA。于上海生工合成正反向引物F: 5′-GCCGAACGGGAAATTGTC-3′和R: 5′-AGAGATGGCTGGAAGAGG-3′,以β-肌动蛋白基因作为内部参照,正反向引物为F: 5′-GCCGAAC GGGAAATTGTC-3′和R: 5′-AGAGATGGCTGGAA GAGG-3′,对DsRED外源基因的表达量进行校正,来实现对DsRED基因表达量的定量解析(Guoetal., 2012),用2-ΔΔCT法计算得出植物各组织部位中DsRED的表达量。
7)红色荧光蛋白DsRED的检测 分别称取DsRED体胚、组培苗的根、茎、小叶(包括对照体胚、对照组培苗)各100 mg鲜组织,用植物蛋白抽提试剂盒(C500053, 生工)提取蛋白。Western Blot免疫表达条件如下: 以100 V 120 min为条件进行12%(W/V)SDS-PAGE垂直电泳,以110 V 70 min为条件转膜至孔径为0.22 μm的硝酸纤维素印迹膜,5%脱脂牛奶(W/V)封闭1 h, Anti-RFP Taq(D195308, 生工)与β-Actin(AC009, ABclonal)以1∶1 000比例稀释作为一抗孵育2 h,HRP标记的羊抗鼠IgG(D110087, 生工)以1∶1 000比例稀释作为二抗孵育1 h,ECL发光剂显影后用Amersham Imager 600成像仪(美国通用电气公司)成像拍照。利用ImageJ软件对WB条带进行灰度分析(牛东东等, 2014),计算蛋白质相对表达量。
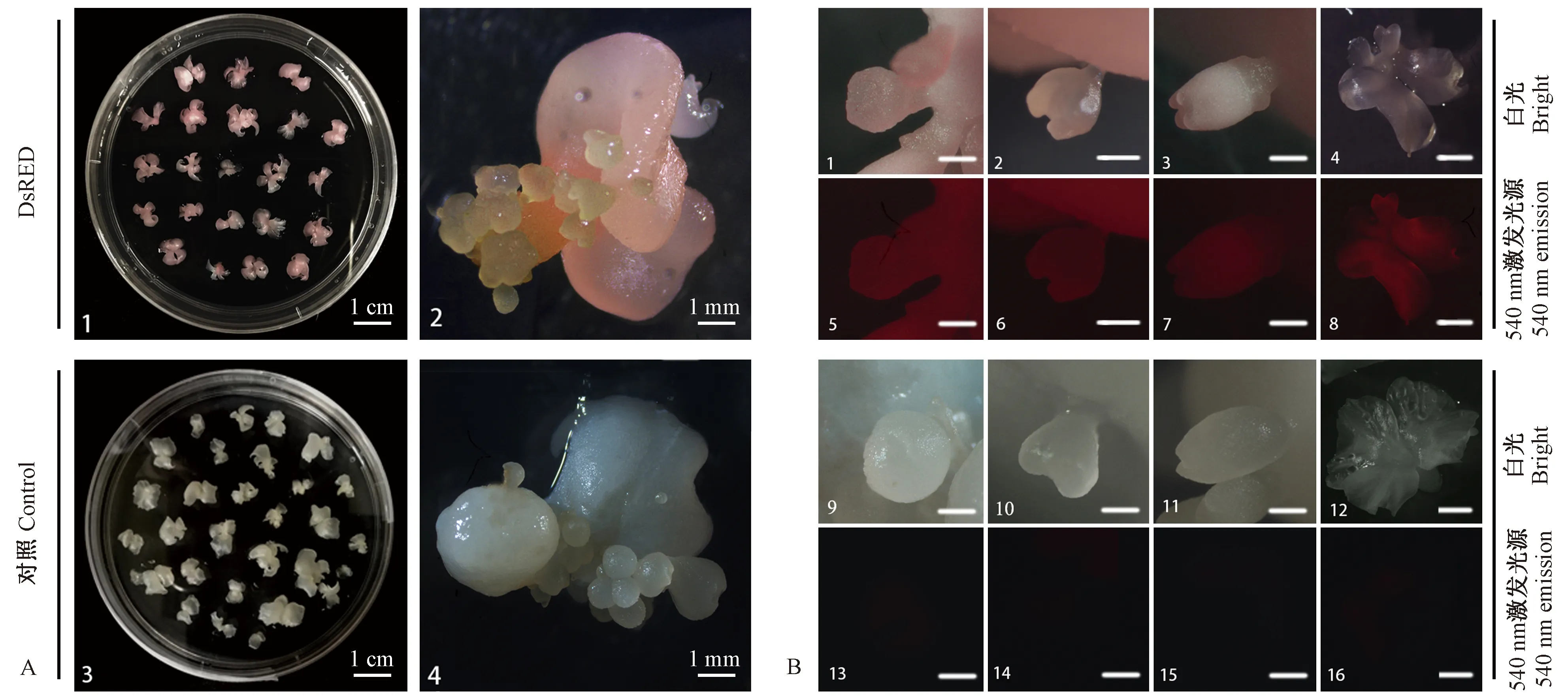
图1 红色荧光蛋白基因DsRED在核桃体胚中稳定表达Fig.1 Red fluorescent protein gene DsRED expressed stably in walnut somatic embryosA. 核桃体胚正常增殖 1,2: DsRED体胚; 3,4: 对照体胚。B. 核桃体胚发育阶段(标尺: 500 μm) 1-4: 分别为白光视野下DsRED体胚的球形胚、心形胚、鱼雷胚和子叶胚表型; 5-8: 分别为荧光视野下DsRED体胚的球形胚、心形胚、鱼雷胚和子叶胚表型; 9-12: 分别为白光视野下对照体胚的球形胚、心形胚、鱼雷胚和子叶胚表型; 13-16: 分别为荧光视野下对照体胚的球形胚、心形胚、鱼雷胚和子叶胚表型。A. Walnut somatic embryo proliferation 1,2: DsRED somatic embryos; 3,4: Control somatic embryos. B. Walnut somatic embryo developmental stages(Bar: 500 μm) 1-4: Globular embryo, heart-shaped embryo, torpedo embryo, cotyledonary embryo of DsRED somatic embryos in bright field; 5-8: Globular embryo, heart-shaped embryo, torpedo embryo, cotyledonary embryo of DsRED somatic embryos in fluorescent field; 9-12: Globular embryo, heart-shaped embryo, torpedo embryo, cotyledonary embryo of control somatic embryos in bright field; 13-16: Globular embryo, heart-shaped embryo, torpedo embryo, cotyledonary embryo of control somatic embryos in fluorescent field.
1.3 数据处理 试验数据用SPSS 22.0软件进行单因素方差分析,采用LSD法进行多重比较,利用GraphPad Prism 7作图。
2 结果与分析
2.1 红色荧光蛋白基因DsRED在核桃体胚中表达稳定 本研究所用的携带DsRED荧光蛋白的核桃体胚于2015年通过农杆菌介导转化法成功获得, 2017年再次通过荧光和PCR鉴定其阳性。目前,核桃DsRED体胚在自然光下呈粉红至红色,而对照体胚在自然光下呈白色,二者均可正常生长与增殖(图1A)。DsRED体胚经过3年增殖和继代培养后,通过镜检发现,DsRED基因在核桃阳性体胚中表达稳定,阳性体胚呈比较均一的粉红色至红色,形态正常,增殖良好; DsRED核桃体胚的增殖途径大多为直接发生途径,即次生胚不经历愈伤组织途径而主要从母胚的子叶表面产生,体胚发育经历了球形胚、心形胚、鱼雷胚和子叶胚4个阶段,各发育阶段体胚在白光下呈粉红至红色,540 nm荧光激发下呈现明亮的红色(图1B 1-8)。同样胚龄的对照体胚生长增殖良好,增殖途径为体胚直接发生途径,体胚发育同样经历了球形胚、心形胚、鱼雷胚和子叶胚4个阶段,各发育阶段体胚在白光下呈白色至乳白色,540 nm荧光下无激发(图1B 9-16)。以上结果表明DsRED基因对核桃体胚生长和增殖无不良影响且可以在体胚中稳定表达。为了排除体胚荧光假阳性,对DsRED体胚DsRED基因在DNA和mRNA层面的表达水平分别进行PCR和qRT-PCR检测。研究结果表明,DsRED体胚扩增获得目的条带,条带大小与预期一致(681 bp),对照体胚无相应条带(图2A)。进一步通过qRT-PCR检测DsRED体胚mRNA的相对表达量,发现体胚mRNA相对表达量与对照体胚具显著性差异,将DsRED体胚DsREDmRNA相对表达量定为1.0,则对照体胚的DsREDmRNA相对表达量为0(图2B)。
为了检测DsRED基因的翻译水平,进一步对DsRED体胚进行Western Blot免疫表达分析。试验结果表明,DsRED体胚具有强阳性条带,而对照体胚中无特异性条带(图3A)。进一步利用ImageJ软件进行WB条带灰度分析,计算结果表明,DsRED体胚荧光蛋白表达量与对照相比具显著性差异。将DsRED体胚荧光蛋白相对表达量定义为1.0,则对照组体胚的表达量为0(图3B)。表明DsRED在核桃体胚中可正常翻译并稳定表达。
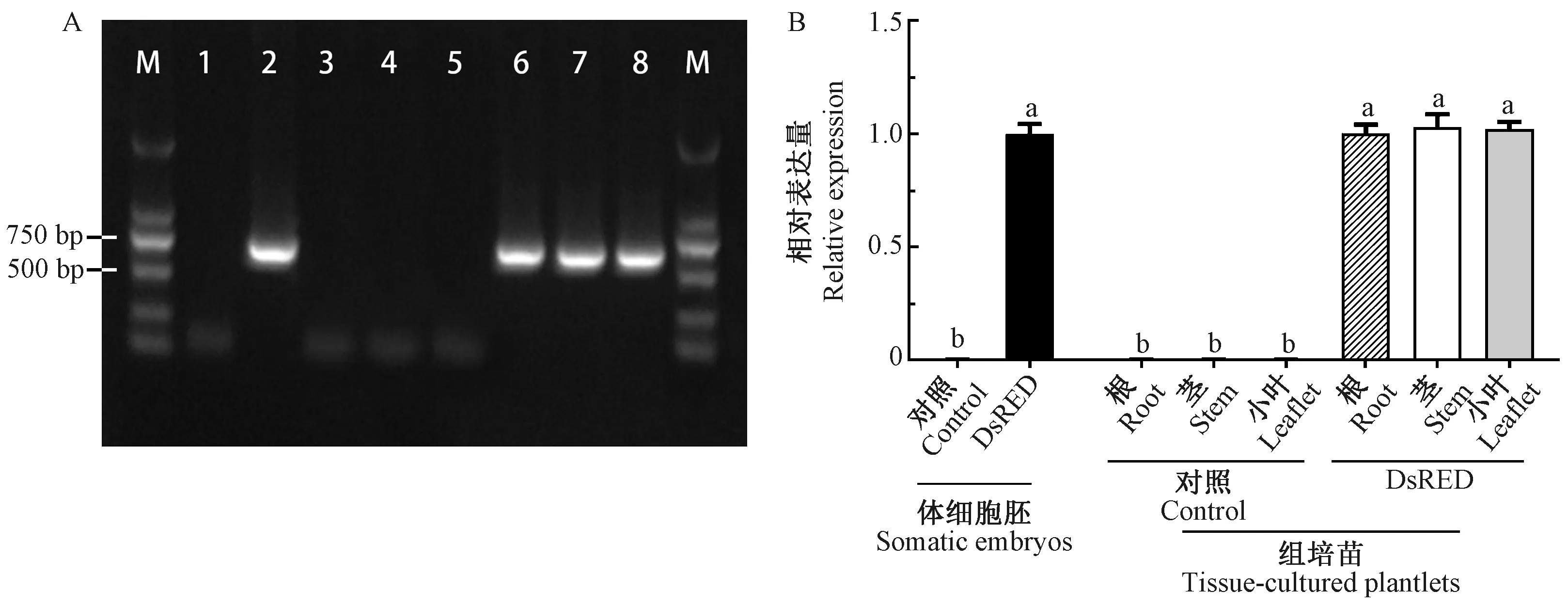
图2 PCR和qRT-PCR对核桃体胚和组培苗的DsRED基因的检测Fig.2 PCR and qRT-PCR assay of DsRED gene in walnut somatic embryos and tissue-cultured plantletsA. DsRED基因的PCR检验 M: Marker; 1: 对照体胚; 2: DsRED体胚; 3-5: 对照组培苗根、茎、小叶; 6-8: DsRED组培苗根、茎、小叶。B. DsRED mRNA的相对表达量(P<0.05)。A. PCR assay of DsRED gene M: Marker; 1: Control somatic embryo; 2: DsRED somatic embryo; 3-5: Root, stem, leaflet of control tissue-cultured plantlet; 6-8: Root, stem, leaflet of DsRED tissue-cultured plantlet. B. Relative expression of DsRED mRNA(P<0.05).
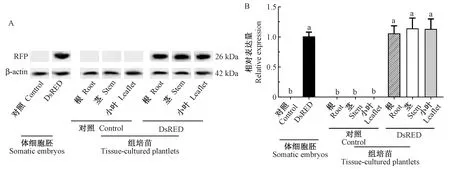
图3 Western Blot检测核桃体胚和组培苗中的DsRED红色荧光蛋白Fig.3 Western Blot assay of red fluorescent protein DsRED in walnut somatic embryos and tissue-cultured plantletsA. DsRED红色荧光蛋白的免疫印迹; B. DsRED红色荧光蛋白的相对表达量(P<0.05)。A. Western Blot of DsRED red fluorescent protein; B. Relative expression of DsRED red fluorescent protein (P<0.05).
2.2 红色荧光蛋白基因DsRED在核桃组培苗中表达稳定 1)核桃DsRED组培苗表型和荧光表达分析 将荧光检测呈明亮红色且PCR、qRT-PCR验证为阳性的子叶胚以及对照子叶胚进行植株再生获得DsRED和对照组培苗,而后常规增殖培养(图4A1,2)。待新芽培养3~4周,分别检测其表型,发现DsRED组培苗为奇数羽状复叶,每复叶平均小叶数为7,叶长椭圆形,长平均0.97 cm,宽平均0.48 cm,叶面积平均0.69 cm2,株高平均5.72 cm,株幅平均3.54 cm,茎粗平均0.24 cm; 对照植株每复叶平均小叶数为7,长平均0.98 cm,宽平均0.46 cm,叶面积平均0.74 cm2,株高平均5.69 cm,株幅平均3.43 cm,茎粗平均0.26 cm。DsRED组培苗各指标与对照相比无显著差异(P<0.05)(表1)。

图4 红色荧光蛋白基因DsRED在核桃组培苗中稳定表达Fig.4 Red fluorescent protein gene DsRED expressed stably in walnut tissue-cultured plantletsA. 组培苗 1: DsRED组培苗; 2: 对照组培苗。 B. 组培苗外部荧光表达(标尺: 3 mm) 1-3: 分别为DsRED组培苗白光下根、茎、小叶外形; 4-6:分别为DsRED组培苗荧光下根、茎、小叶外形; 7-9: 分别为对照组培苗白光下根、茎、小叶外形; 10-12: 分别为对照组培苗荧光下根、茎、小叶外形。 C. 组培苗显微结构荧光表达(标尺: 600 μm) 1-3: 分别为DsRED组培苗白光下根、茎、小叶横切面; 4-6: 分别为DsRED组培苗荧光下根、茎、小叶横切面; 7-9: 对照组培苗白光下根、茎、小叶横切面; 10-12: 对照组培苗荧光下根、茎、小叶横切面。A. In vitro plantlets 1: DsRED tissue-cultured plantlet; 2: Control tissue-cultured plantlet. B. Appearance of tissue-cultured plantlets(Bar: 3 mm) 1-3: Root, stem, leaflets of DsRED tissue-cultured plantlet in bright field; 4-6: Root, stem, leaflets of DsRED tissue-cultured plantlet in fluorescent field; 7-9: Root, stem, leaflets of control tissue-cultured plantlet in bright field; 10-12: Root, stem, leaflets of control tissue-cultured plantlet in fluorescent field. C. Section(Bar: 600 μm) 1-3: Cross section of root, stem, leaflet of DsRED tissue-cultured plantlet in bright field; 4-6: Cross section of root, stem, leaflets of DsRED tissue-cultured plantlet in fluorescent field; 7-9: Cross section of root, stem, leaflets of control tissue-cultured plantlet in bright field; 10-12: Cross section of root, stem, leaflets of control tissue-cultured plantlet in fluorescent field.
为了明确增殖培养对组培苗中DsRED基因表达的影响,将DsRED组培苗的茎尖、茎段、小叶和根置于荧光体视显微镜下镜检,结果表明,DsRED组培苗的茎尖、茎段、小叶以及根表面均呈现明亮且较为均一的红色,其中叶柄切除处颜色鲜艳,可能是由于该处维管组织丰富(图4B 1-6); 对照植株的相应结构均在荧光下无激发,视野中呈现黑色(图4B 7-12)。为了进一步验证DsRED红色荧光蛋白基因在组培苗中表达的稳定性,对组培苗营养器官的显微结构也进行了荧光检测,结果表明,在组培苗的根、茎、叶中,DsRED红色荧光蛋白基因可稳定表达。其中,在根的表皮、皮层、维管柱以及髓薄壁细胞中均有表达,且维管柱中颜色最鲜艳,提示该部位表达量最高; 在茎中,在表皮、皮层、维管形成层以及髓薄壁细胞中均有表达,其中维管形成层部位的细胞荧光表达量最高,表皮细胞表达量次之,皮层薄壁细胞和髓薄壁细胞表达量较弱; 在叶的横切片中可以发现,维管束和叶肉细胞表达量较高(图4C 1-6)。对照植株的根、茎、叶横切面在540 nm荧光下无激发,显微镜视野中呈现黑色(图4C 7-12)。
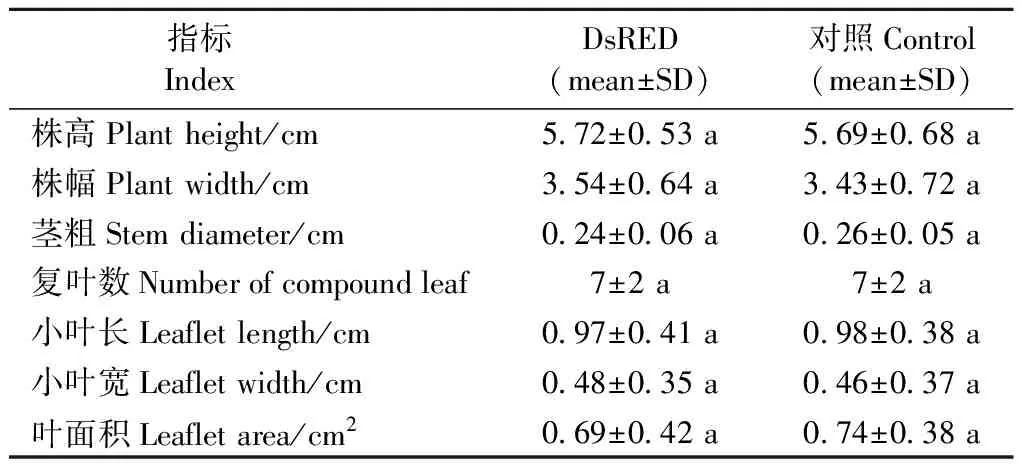
表1 DsRED基因表达对核桃组培苗茎、叶生长的影响①Tab.1 Effect of DsRED gene expression on stem and leaf growth of walnut tissue-cultured plantlets
2)核桃组培苗DsRED基因表达分析 为了排除荧光假阳性,对DsRED组培苗的DsRED基因在DNA和RNA水平的表达分别进行PCR和qRT-PCR分析。结果表明,DsRED组培苗的根、茎、叶均扩增获得目的条带,条带大小与预期一致(681 bp),对照组培苗无相应条带(图2A)。通过qRT-PCR检测DsRED组培苗的根、茎、叶中DsREDmRNA的相对表达量,发现组培苗根的DsREDmRNA相对表达量为0.97±0.05,茎为1.03±0.08, 小叶为1.02±0.04(将荧光阳性体胚DsREDmRNA相对表达量定为1.0),表明DsRED组培苗的根、茎、叶和DsRED体胚的表达量无显著性差异,而对照组培苗中的DsREDmRNA相对表达量均为0(图2B)。
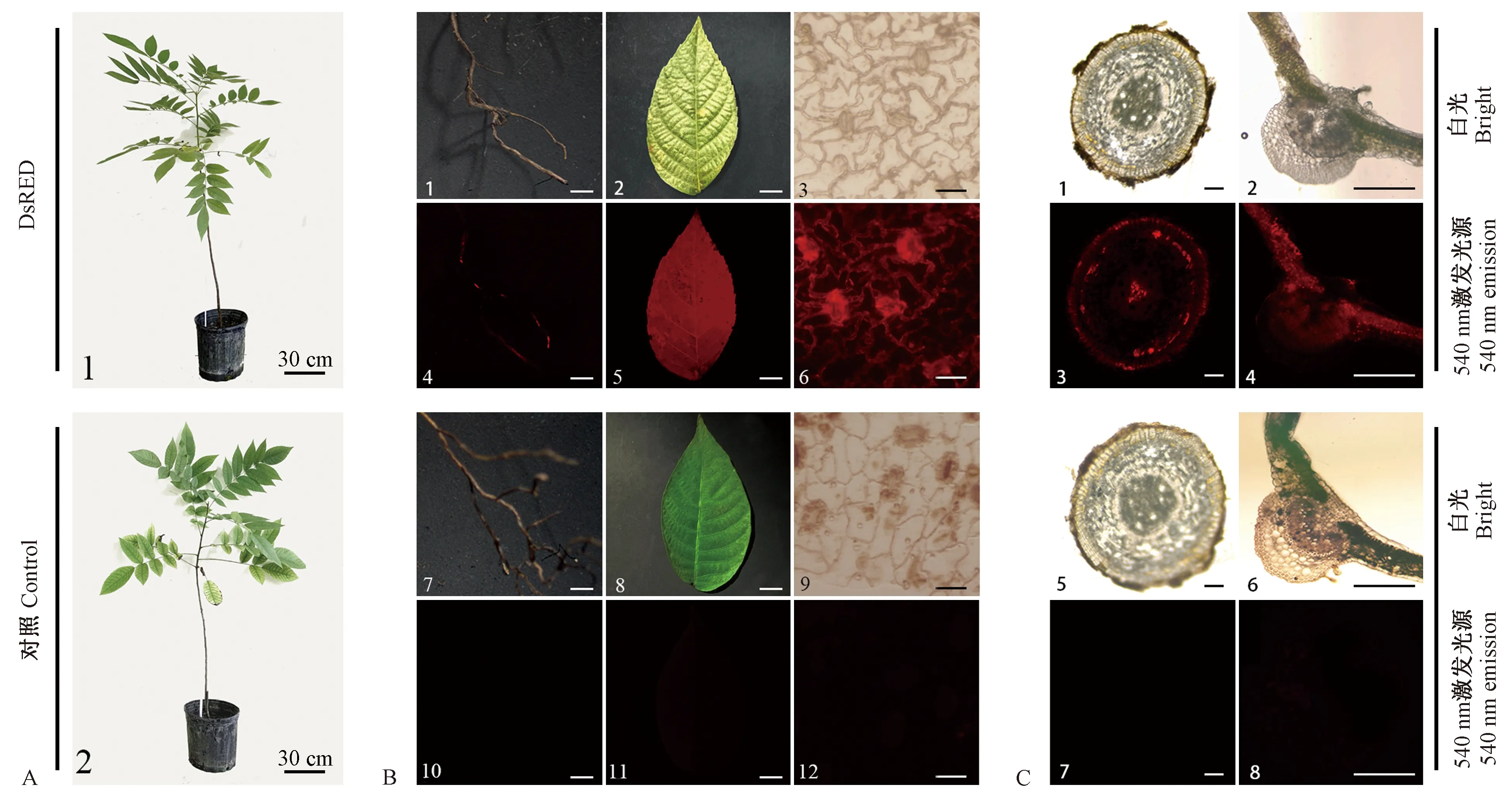
图5 DsRED红色荧光蛋白在3年生苗中稳定表达Fig.5 DsRED red fluorescent protein expressed stably in 3-year-old walnut regenerated plantsA. 3年生苗 1: DsRED 3年生苗; 2: 对照3年生苗。B. 3年生苗外部荧光表达(标尺 3, 6, 9, 12: 10 μm; 其余: 1 cm) 1-3: 分别为DsRED 3年生苗根、小叶和叶表皮在白光下; 4-6: 分别为DsRED 3年生苗根、小叶和叶表皮在荧光下; 7-9: 分别为对照3年生苗根、小叶和叶表皮在白光下; 10-12: 分别为对照3年生苗根、小叶和叶表皮在荧光下。C. 3年生苗显微结构荧光表达(标尺: 300 μm)1, 2: 分别为DsRED 3年生苗根和小叶在白光下的横切面; 3, 4: 分别为DsRED 3年生苗根和小叶在荧光下的横切面; 5, 6: 分别为对照3年生苗根和小叶在白光下的横切面; 7, 8: 分别为对照3年生苗根和小叶在荧光下的横切面。A. 3-year-old regenerated plants 1: DsRED 3-year-old regenerated plant; 2: Control 3-year-old regenerated plant. B. Appearance of 3-year-old regenerated plants(Bar 3, 6, 9, 12: 10 μm; the rest: 1 cm)1-3: Root, leaflet, leaf epidermis of DsRED 3-year-old regenerated plant in bright field; 4-6: Root, leaflet, leaf epidermis of DsRED 3-year-old regenerated plant in fluorescent field; 7-9: Root, leaflet, leaf epidermis of control 3-year-old regenerated plant in bright field; 10-12: Root, leaflet, leaf epidermis of control 3-year-old regenerated plant in fluorescent field. C. Section(Bar: 300 μm) 1, 2: Cross section of root and leaflet of DsRED 3-year-old regenerated plant in bright field; 3, 4: Cross section of root and leaflet of DsRED 3-year-old regenerated plant in fluorescent field; 5, 6: Cross section of root and leaflet of control 3-year-old regenerated plant in bright field; 7, 8: Cross section of root and leaflet of control 3-year-old regenerated plant in fluorescent field.
为了检测DsRED基因在组培苗中的翻译水平,进一步开展了Western Blot免疫表达分析。结果表明,DsRED组培苗根、茎和小叶中均有强阳性条带,而所有对照组培苗中无特异性条带(图3A)。进一步利用ImageJ软件进行WB条带灰度分析,计算结果表明,DsRED组培苗根、茎、小叶中的相对表达量分别为1.06±0.14,1.14±0.19,1.13±0.19(DsRED体胚相对表达量定为1.0),各组织间无显著性差异,而对照组培苗根、茎、叶中的表达量均为0(图3B)。表明DsRED在核桃组培苗中可正常翻译并稳定表达。
2.3 红色荧光蛋白基因DsRED在核桃3年生苗中表达稳定 将培养获得的健壮DsRED组培生根苗(含对照)移栽至温室。DsRED组培苗生根驯化后成活率高,成活后移栽至大棚正常生长3年(图5A)。对DsRED 3年生苗表型进行测定,发现DsRED 3年生苗叶片同样为奇数羽状复叶,顶端小叶最大,其下对生小叶依次变小,每片复叶平均小叶数11片。小叶长椭圆形,平均长15.32 cm,平均宽5.84 cm,平均叶面积57.41 cm2。每片小叶顶端急尖,基部歪斜,边缘上部具稀疏细锯齿、近基部全缘。植株株高92.88 cm,平均株幅71.78 cm,平均茎粗0.89 cm。对照苗每复叶平均小叶数11,长平均14.52 cm,宽平均5.63 cm,叶面积平均50.33 cm2,株高平均94.14 cm,株幅平均69.24 cm,茎粗平均0.88 cm。DsRED植株各指标与对照相比无显著性差异(P<0.05)(表2)。表明DsRED基因对核桃植株叶形等表型没有产生影响。
体视显微镜荧光镜检结果表明,DsRED 3年生苗小叶及根表面均呈现明亮的红色,同时,叶表皮细胞荧光检测发现,细胞壁中荧光表达量明显高于原生质,气孔保卫细胞的DsRED荧光表达量高于其他表皮细胞,对照苗的相应结构均在荧光下无激发,视野中呈现黑色(图5B)。为了进一步跟踪检测DsRED红色荧光蛋白在核桃DsRED 3年生苗显微结构中的表达,对3年生苗也进行了根和叶的横切面荧光检测。结果表明,DsRED红色荧光蛋白在根的横切面上有明显表达,其中,在初生木质部、维管形成层以及周皮中表达量较高,在皮层薄壁细胞以及次生木质部表达量稍低,而对照3年生苗在荧光视野下无激发,视野中呈现黑色(图5C)。
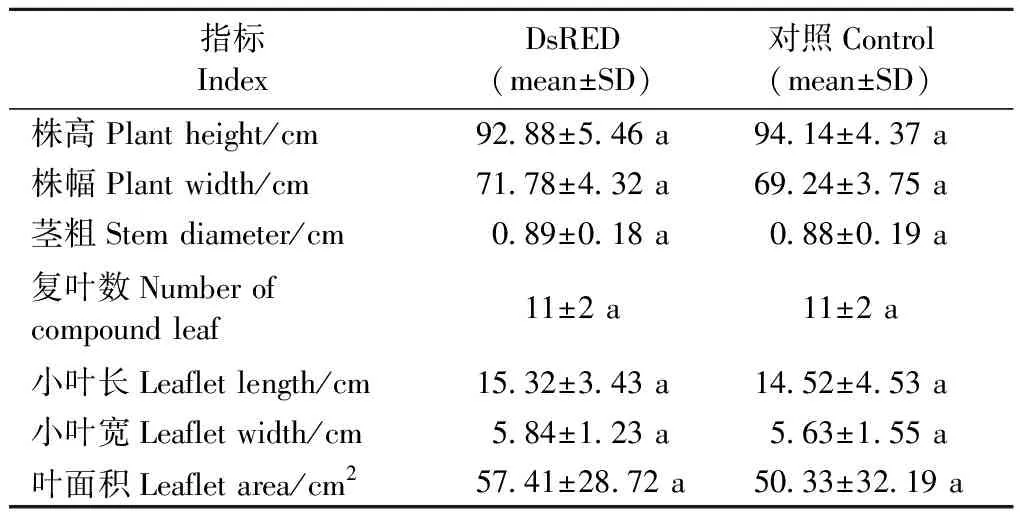
表2 DsRED基因表达对核桃3年生苗茎、叶生长的影响Tab.2 Effect of DsRED gene expression on stem and leaf growth of 3-year-old walnut regenerated plants
3 讨论
报告基因因其优异性状被广泛应用,DsRED作为一个新型报告基因,已在多个物种中成功应用(高嵩等, 2017)。在真菌中,通过农杆菌介导法将DsRED荧光蛋白基因转入从北京玉米(Zeamays)品种‘西玉3号’病株分离出的新月弯孢(Curvularialunata)菌株中,成功对玉米新月弯孢菌株进行遗传标记并获得稳定遗传(陈茂功等, 2012); 将DsRED通过农杆菌介导法转入拟轮生镰刀菌(Fusariumverticillioides)中,深入探究拟轮生镰刀菌与水稻间的互作关系(Wuetal., 2016)。在农作物中,采用农杆菌侵染萌动胚的方法,将DsRED基因转入玉米品种‘郑单958’中,用于快速筛选含有目标基因的种子(高嵩等,2017);DsRED基因被用于研究水稻(Oryzasativa)胚乳中淀粉体分裂和淀粉合成涉及的分子机制(Wuetal., 2016); 将DsRED基因转入陆地棉(Gossypiumhirsutum)愈伤组织中,之后在愈伤组织、体细胞胚和大多数再生植株的组织、器官中都可以检测到红色荧光,并获得多个品系,进一步将其中转基因品系Yz-2-DsRed2作为雄性亲本与其他棉花品种杂交后,红色的遗传力依旧非常稳定(Sunetal., 2018); 转入DsRED基因的大豆(Glycinemax)依旧可育,自花授粉得到的T0代种子在荧光显微镜下显示亮红色荧光,继续培养得到T2代种子,其与非转基因大豆生长方式相似,依旧表达DsRED基因且未出现性状分离(Nishizawaetal., 2006)。与DsRED在棉花体胚中的表现相似,在本研究中,试验材料自Zhang等(2015)通过农杆菌介导将DsRED基因转入核桃体胚,至今阳性体胚已继代培养5年,DsRED基因依然能够在核桃体胚中稳定表达,阳性体胚外表依旧呈粉红色,并且能在540 nm激光下发出明亮红色荧光。可见DsRED基因可在核桃体胚中稳定表达且对核桃体胚的增殖和生长无不良影响。
转基因植株离体保存需利用植物组织培养技术。周瑞金等(2011)研究证明外源基因在继代培养6~8年的组培苗中稳定存在,并在大部分株系中表达。不仅在组培苗中,外源Bt基因能够在8年生741杨[Populusalba× (P.davidiana+P.simonii)×P.tomentosa]中稳定表达(陈盼飞等, 2016)。王倩倩等(2013)对继代保存了13年的转基因苹果(Malusdomestica)组培苗和在田间定植 6~8 年的转基因植株中外源基因的遗传和表达进行研究,结果表明所有苹果转基因株系组培苗中均可检测出CpTI特异基因片断,转入的外源CpTI基因在所测田间株系的花粉、叶片及大部分的根系中均能检测到,但在极少数株系的根系中未能检测到特异条带,表明外源基因可在转基因植株中稳定存在,但在不定根发生过程中可能会出现丢失现象,这与曾凡锁等(2008)对外源基因(Bgt基因、GUS基因、NptⅡ基因)在转基因白桦(Betulaplatyphylla)中的结果一致。本研究对核桃DsRED继代培养3年的组培苗和温室3年生苗进行检测的结果表明, 核桃DsRED组培苗和3年生苗各组织部位中均稳定高度表达DsRED基因,可见DsRED基因的表达对核桃植株再生和生长无不良影响,且其表达具有高度的稳定性。
然而,经过多代培养的转基因组培苗还涉及材料变异和外源基因是否丢失的问题。如香蕉(Musanana)(朱靖杰, 1995)、草莓(Fragaria×ananassa)(Swartz, 1981)、番木瓜(李卫东等, 2006)的离体培养后代有明显的变异,且随继代次数的增加,变异机率会大大增加,朱靖杰(1995)和李卫东等(2006)分别提出,香蕉与番木瓜离体培养代数应分别控制在10代和32代内。此外,外源基因的插入可能会导致细胞分裂过程的异常,从而造成外源基因的丢失(李小方等, 2001)。外源基因进入受体后因拷贝数大小不一、插入位点不定、整合的DNA结构变异大及受外界环境因素诱导等可能会导致受体植物中的外源基因出现失活、沉默、丢失等现象,从而影响外源基因在受体植物及其后代中的稳定性(赵彦平等, 2011)。报告基因作为外源基因能否在试验对象生长过程及后代繁殖中稳定表达需要进一步研究。Taylor(1997)认为外源基因的表达水平在后代延续过程中会发生变化。有研究发现,植物的生长发育及复杂的外界环境都会对导入植物材料的外源基因造成失活和沉默,从而影响外源基因的表达稳定性,报告基因也不例外(Stemetal., 1997; 夏兰芹等, 2000)。转玉米二氢酮醇还原酶基因(A1)的矮牵牛(Petuniahybrida)移栽至大田后,由于环境温度升高、光照增强,DNA的甲基化引起外源基因的失活(Meyer, 2000)。研究发现,盐胁迫在一定程度上可以诱导转基因107杨(Populus×euramericana‘4/76’)的外源Bt基因、BADH基因表达加强,推测正常条件下外源基因表达受到植物的某种抑制,胁迫改变了植物的代谢,缓解对外源基因的表达抑制(陈盼飞等, 2017)。可见,不同外源基因在不同试验材料中的表达稳定性有所不同。在报告基因中,有研究发现红、绿、黄荧光蛋白标记根瘤菌接合子在传代 5、10、20 代的过程中,荧光质粒丢失率不断增加(杨晓玫等, 2018)。同样地早期GUS染色为阳性的橡胶树(Heveabrasiliensis),在连续继代再生植株的过程中出现了基因丢失的现象(李季等, 2013)。在转GUS基因白桦中,11个转基因植株中有2株出现了GUS基因沉默,其余植株均有不同水平的GUS表达,且这种差异与GUS基因的拷贝数没有必然联系(曾凡锁等, 2009)。
本研究中,尽管DsRED在核桃体胚、组培苗及3年生苗中表达稳定,但后续表达是否会降低或丢失,表达部位是否具有组织特异性,如在花、果实和种子中是否会表达等,还需要进一步跟踪研究。此外,有关荧光蛋白的研究目前主要集中于荧光蛋白的实际应用(Dongetal., 2018),缺乏对荧光蛋白本身的功能性和遗传动态的研究(Webbetal., 1995; 杨晓玫等, 2019)。若能深入了解荧光蛋白的发光机理、标记条件、适用结构等,可更有针对性地选择相应的荧光蛋白用于科学研究,对于指导荧光蛋白的精准定位具有十分重要的意义。
4 结论
红色荧光蛋白基因DsRED转入核桃体胚后,不仅能够在体胚中稳定表达,并且在继代培养3年的组培苗和3年生温室苗中同样能够稳定表达。进一步在DNA、mRNA和蛋白水平分析表明,DsRED基因在核桃各组织部位中均有表达,是核桃理想的遗传转化报告基因。本研究结果可为DsRED作为报告基因在果树中的应用提供参考。
