喀斯特区改良与未改良草地植被和土壤特征研究*
2021-01-22李梦瑶赵一军宣文婷毛文娅
李梦瑶 ,赵一军,王 文,宣文婷,毛文娅,徐 震
(1.兰州大学,农业农村部草牧业创新重点实验室,甘肃 兰州 730020;2.兰州大学 草地农业科技学院,甘肃 兰州 730020;3.云南省种羊繁育推广中心,云南 昆明 655204;4.兴义市众犇农牧发展有限公司,贵州 兴义 562407)
东非狼尾草(Pennisetum clandestinum)又称隐花狼尾草、铺地狼尾草,是禾本科狼尾草属的多年生草本植物,具发达根状茎,茎节着地生根且须根粗硬,入土深,能够在贫瘠土壤中生长,叶片柔嫩多汁,适口性好,为各类家畜所喜食[1]。由于东非狼尾草既有较好的适口性,又有较强的抗逆性和侵占性,故其在水土保持、草坪和草地建植及改良等方面具有十分广阔的应用前景[2]。东非狼尾草原产于东非高原,目前作为重要牧草资源被广泛栽培于澳大利亚、中国、新西兰、南非和美国等国家及地区[3]。自20 世纪90 年代引入中国云南以来,东非狼尾草得到广泛栽培,呈现出良好的生态适应性和竞争优势,成为云南省人工草地建植的重要草种[4],并用于草坪建植、高速公路护坡、园林绿化、脆弱生态环境修复、水土保持及恶性杂草防控等方面[5]。
研究表明:东非狼尾草能自然侵入白茅(Imperata cylindrica)、石生繁缕(Stellaria vestita)和牛尾蒿(Artemisia dubia)等群落,促进这些群落的良性发展,且能够替代入侵物种紫茎泽兰(Ageratina adenophora)[6]。同时,东非狼尾草与白三叶(Trifolium repens)混播能维持草地群落稳定性,获得较高草地生产力[7]。东非狼尾草对氮肥敏感,新枝长出后,施适量氮肥促能其生长[8]。
目前,对于东非狼尾草的研究大多集中于其生长特性及牧草品质[9-10]等方面,而将其用于石砾区域天然草地或草山草坡改良的研究相对较少。由于中国南方喀斯特山区许多区域属严重石砾地,缺乏土壤,难于耕作和建植人工草地,因此该地形的草地改良一直是人工草地建植的难点。本研究通过对东非狼尾草改良草地和未改良天然草地的植物种重要值、物种多样性、生物量构成、草土养分特征和群落稳定性特征的分析,研究南方喀斯特石砾区域天然草地适宜的建植方法,为东非狼尾草在中国南方喀斯特山区石砾地的开发利用和退化草地改良提供实践依据。
1 材料与方法
1.1 试验地概况
试验地在云南省寻甸回族彝族自治县种羊场(N25°36′42″,E103°13′38″),平均海拔2 008 m,降水多集中在6—8 月,年均降雨量1 043 mm,年均温为13.5 ℃,土壤类型为砖红壤,土壤相对黏重,保水能力较差,pH 值为5.6~6.1。研究地属低纬高原季风气候,冬春两季受平直西风环流控制,大陆季风气候明显,干旱少雨;夏秋季主要受太平洋西南或印度洋东南暖湿气流控制,海洋季风突出,多雨,凉爽潮湿。
1.2 样地设置
2018 年6 月,在试验地随机选择改良草地样地和未改良天然草地样地各3 个,面积分别为0.3 和0.1 hm2。
改良草地于2014 年8 月播种,改良前为不平整坡地,采用机械平整原生草地后拣出较大石块,播种草种数量比例为东非狼尾草∶鸭茅(Dactylis glomerata)∶多年生黑麦草(Lolium perenne):白三叶=6∶2∶1∶1。草地播种后每年9—10 月中旬施肥1 次,即:施钙镁磷复合肥(过磷酸钙) 300 kg/hm2、硫酸钾(K2SO4) 75 kg/hm2和尿素(含氮46.2%) 75 kg/hm2;在牧草生长季(4—11 月)的每月中旬进行家畜轮牧利用,每次放牧8~10 d,每次轮牧时间和家畜数量依据牧前牧草生长状况而变化,牧后草层高度为5~8 cm。
未改良天然草地主要植物为白茅、黑穗画眉草(Eragrostis nigra)、牛尾蒿、石生繁缕、百里香(Thymus vulgaris)和猪殃殃(Galium aparine)等[11]。由于临近改良草地,随着家畜放牧采食和轮牧转场,畜粪和皮毛将鸭茅和多年生黑麦草种子携带入未改良天然草地,进而未改良天然草地中也出现少量鸭茅和多年生黑麦草。
1.3 测定指标和方法
1.3.1 植物群落特征调查
2018 和2019 年的7 月中旬—8 月中旬,在各个改良和未改良草地样地上均匀设置15 个50 cm×50 cm 的样方,调查各样方内所有植物种的高度、密度(以株计算)、盖度和地上生物量。地上生物量齐地刈割后,将收获的各样方地上生物量的死物质(凋落物+立枯物)和活物质分开;再将活物质按不同植物种分开后,于65 ℃下烘干称干质量。
1.3.2 植物种重要值
基于样方植物种的高度、盖度和密度数据,按公式IV=(RH+RC+RD)/3 计算植物种重要值。式中,RH、RC 和RD 分别为植物种的相对高度、相对盖度和相对密度。
1.3.3 植物种多样性指数
基于样方草地群落植物物种数和植物种重要值数据,按以下公式计算群落植物丰富度指数、Shannon-Wiener 指数、Simpson 指数和Pielou 均匀度指数[12]。
丰富度指数:R0=S;

式中,S指出现在某一草地类型中的物种数;Pi指某个草地类型中第i个物种的相对重要值,Pi=Ni/N(Ni指该草地类型中的第i个物种的重要值;N指该草地类型中所有物种重要值之和)。
1.3.4 地下生物量测定及生物量构成分析
在测定完地上生物量的各样方内,用直径9 cm的根钻分别采集0~15 cm 深度根样,捡出石砾,放入尼龙网袋中,流水冲洗干净,于65 ℃下烘干后称干质量。
将1.3.1 节中测定的地上生物量按东非狼尾草、鸭茅+多年生黑麦草、白三叶、非播种禾草和其他5 类经济类群,进行各类群地上生物量占总地上生物量比例分析,并以地上生物量和地下生物量进行地上(活物质和死物质)和地下生物量构成分析。
1.3.5 草土养分分析
将1.3.1 节中地上样方分种牧草的活物质与死物质混合,并将地上混合牧草和地下根系样品分别粉碎后用于牧草养分分析。在各样方内用直径5 cm 的土钻分别采集0~10 cm 土样,捡出石子和草根后,风干过筛用于土壤养分分析。
草土养分分析指标和方法为:草土全氮含量采用凯氏定氮法测定;草土全磷含量采用钼锑抗比色法测定;土壤有机质(organic matter,OM)含量采用重铬酸钾硫酸法测定;牧草中性洗涤纤维(neutral detergent fiber,NDF)、酸性洗涤纤维(acid detergent fiber,ADF)、纤维素(cellulose,CL)、半纤维素(hemicellulose,HCL)、酸性洗涤木质素(acid detergent lignin,ADL) 和酸不溶灰分(acid-insoluble ash,AIA) 含量采用范氏(Van Soest)洗涤纤维分析法和灼烧法测定[13-15]。所有指标数据均换算为干物质基础。
1.3.6 草地稳定性评价
用Raunkiaer 频度系数评价草地植物群落稳定性。Raunkiaer 将植物种频度系数(R) 划分为A、B、C、D 和E 5 个等级,对应的R值分别为1%~20%、>20%~40%、>40%~60%、>60%~80%和>80%~100%,5 个等级物种数占比呈现反“J”形则稳定性高,反之稳定性低[16]。
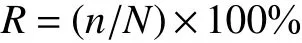
式中,n为某一物种在全部取样中出现的次数,N为全部取样数。
1.4 数据统计分析
在Excel 中处理基础数据并制图,用SPSS 16.0中的独立样本t检验,对草地植物群落物种数、生物量构成及草土养分等进行改良和未改良天然草地之间的差异显著性分析,数据表示为均值±标准误。重要值、植物物种多样性指数和草地稳定性分析数据均以2 年的均值进行计算。
2 结果与分析
2.1 群落植物种重要值
由表1 可知:改良草地群落中东非狼尾草重要值最高,在群落中占绝对优势;其次是白三叶和鸭茅;天然草地中植物物种重要值较高的是黑穗画眉草、白茅和百里香。改良草地与天然草地的物种构成差异较大,改良草地中特有物种为东非狼尾草、狗牙根、金色狗尾草和长柔毛野豌豆等7 种,而天然草地中特有植物种有鼠尾粟、砖子苗、百里香和积雪草等22 种,其草地群落植物物种数(30 种) 大于改良草地(15 种)。说明改良草地中东非狼尾草播种比例高,且其对石砾地适应性较强,从而导致改良草地东非狼尾草重要值占绝对优势。

表1 改良和天然草地植物种重要值Tab.1 Important value of plant species in improved and natural grasslands
2.2 群落植物种多样性指数
由表2 可知:植物丰富度指数、Shannon-Wiener 指数、Pielou 均匀度指数和Simpson 指数均为改良草地<天然草地(P<0.05),说明草地改良降低植物种多样性和植物种分布均匀性。

表2 改良和天然草地物种多样性指数Tab.2 The species diversity indices of improved and natural grasslands
2.3 群落生物量构成
由图1 可知:2018 年和2019 年活物质、地上(活物质+死物质) 生物量、地下生物量及地上+地下(根系) 生物量均为改良草地>天然草地(P<0.05 或P<0.01),而死物质生物量在改良与天然草地之间差异不显著(P>0.05)。
由图2 可知:2018 年和2019 年东非狼尾草和白三叶地上生物量比例为改良草地>天然草地(P<0.001);其中,改良草地中东非狼尾草仍然占绝对优势(78.58%~82.23%),且改良草地白三叶地上生物量比例为天然草地的34.86~76.23 倍。2018 年,鸭茅+多年生黑麦草地上生物量比例为改良草地>天然草地(P<0.05),前者为后者的28.04 倍。2 年期间,非播种禾草和其他类群植物地上生物量比例为改良草地<天然草地(P<0.01或P<0.001),且天然草地的非播种禾草地上生物量占绝对优势(49.71%~64.56%),主要植物为菊科和唇形科。说明东非狼尾草改良石砾地区域效果较好,能有效抑制天然非播种禾草和其他杂类草植物的生长。
2.4 草地草土养分
由表3 可知:2018 和2019 年牧草的中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)、纤维素(CL)、酸性洗涤木质素(ADL)、酸不溶灰分(AIA)含量和土壤有机质含量以及2019 年土壤全氮含量均为改良草地<天然草地(P<0.05);2018 和2019 年牧草全氮和全磷含量、2019 年根系全氮以及2019 年土壤全磷含量均为改良草地>天然草地(P<0.05);2 年牧草半纤维素(HCL)和根系全磷含量在两种草地之间差异不显著(P>0.05)。说明改良草地由于牧草品质改善使家畜采食加强,进而加速养分循环,降低土壤有机质积累。
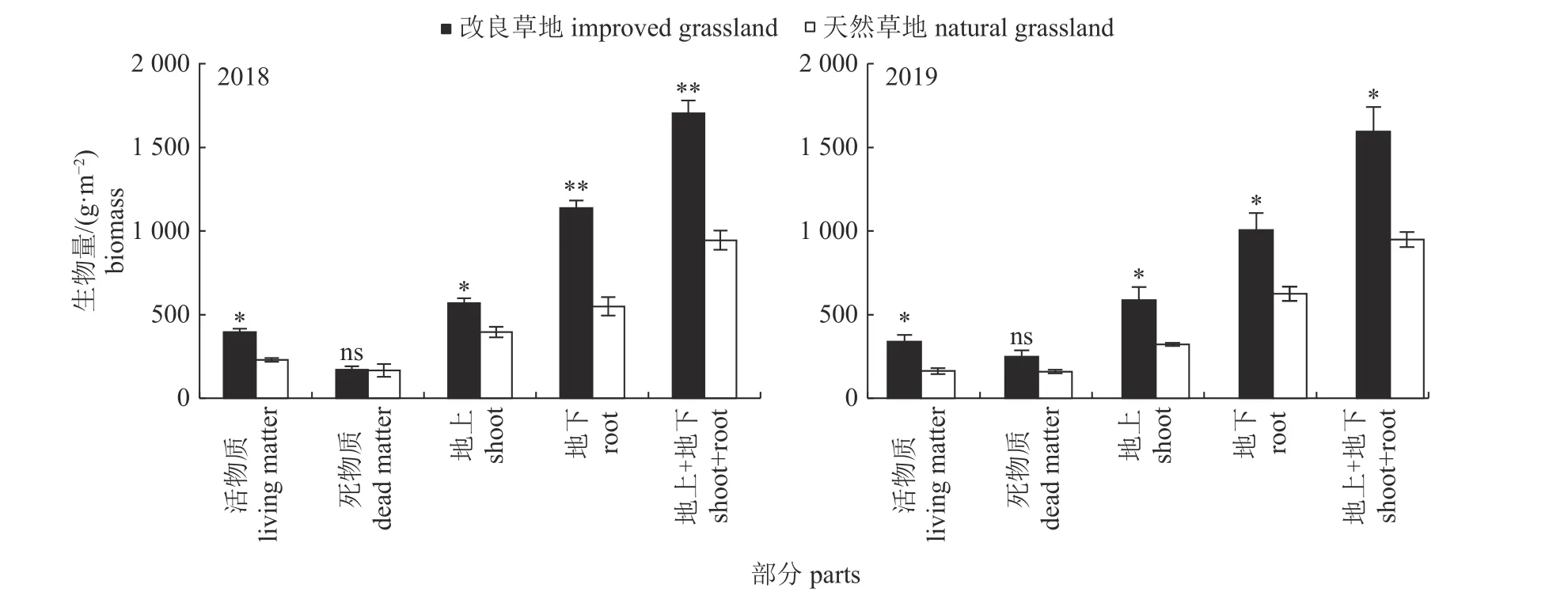
图1 改良和天然草地地上、地下生物量Fig.1 Above-and under-ground biomass in improved and natural grasslands
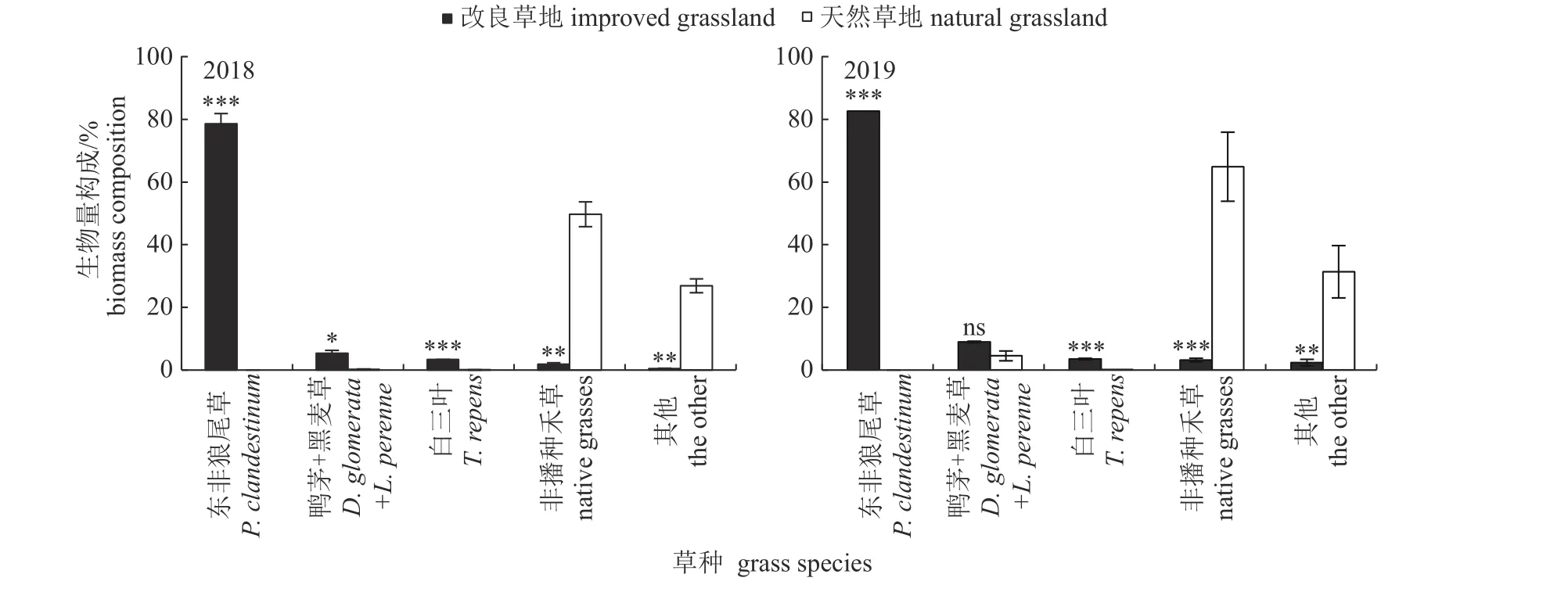
图2 改良和天然草地经济类群地上生物量构成Fig.2 Shoot biomass composition of economic category of improved and natural grasslands
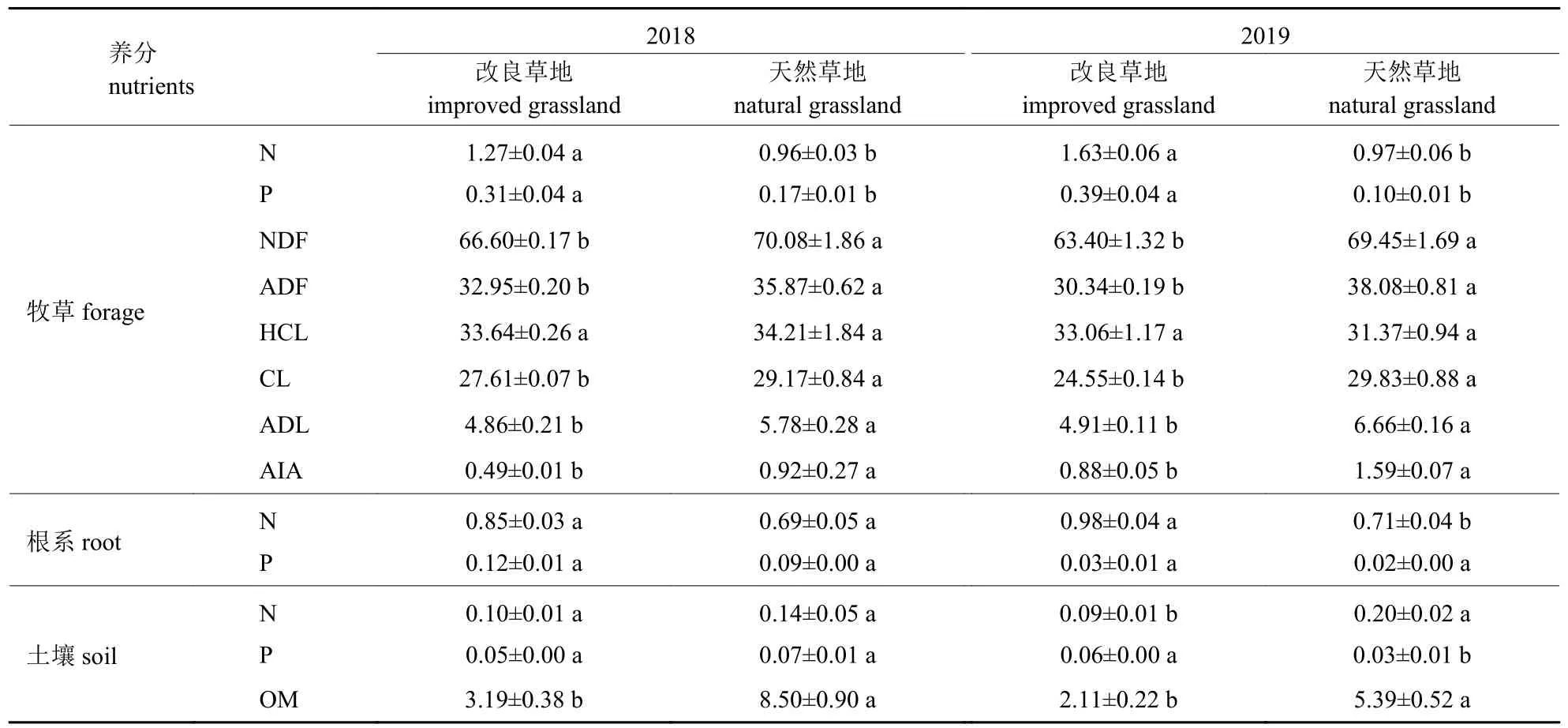
表3 改良和天然草地草土养分含量Tab.3 The nutrient contents in herbage and soil of improved and natural grasslands %
2.5 草地群落稳定性
由图3 可知:改良草地植物群落物种频度图呈反“J”形,符合典型的Raunkiaer 频度定律;而天然草地植物群落物种频度图偏离反“J”形。说明东非狼尾草改良草地植物物种分布较均匀,群落整体处于相对稳定状态;而天然草地群落处于相对不稳定演替阶段。
3 讨论
重要值表示某一种群在群落中的重要性,显示植物种群对环境的适应性[17]。本研究中,改良草地东非狼尾草重要值占绝对优势,其主要原因是改良草地中东非狼尾草播种比例较高,且东非狼尾草具有十分强壮的匍匐茎和根系,根系入土深,匍匐茎能延伸数米长,形成强大地下根茎网,从而其侵占性强,易成为群落优势种[4],由此导致其重要值较高。而天然草地中一些非播种植物种如白茅和黑穗画眉草的重要值相对较高,相较之下,改良草地中播种比例较高的东非狼尾草具有更强的竞争优势,可能在改良草地中抑制了白茅和黑穗画眉草等禾草的生长。这进一步表明:东非狼尾草对石砾地区土壤和其他外界环境具有较强适应性,在喀斯特石砾地区草山草坡改良和禾草/白三叶草地建植中具有较好适应性。
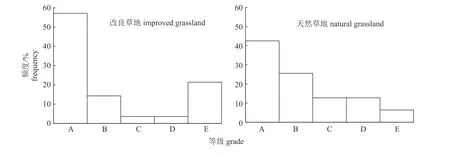
图3 改良和天然草地植物种Raunkiaer 标准频度直方图Fig.3 Raunkiaer standard frequency histogram of plant species in improved and natural grasslands
本研究中,改良草地植物物种多样性指数比天然草地低,其原因可能是:首先,东非狼尾草交错盘结的匍匐茎会在地面形成致密草皮,且其地上直立茎硬挺、叶量丰富[18],该生长特性将导致草层群落下层透光率减弱,降低不耐阴植物种的萌发率和生长,进而降低其竞争优势[19];其次,因东非狼尾草盖度过高,且其致密草层覆盖土壤表面,使其他植物的种子难以接触土壤萌发,进而降低草地植物物种多样性[11];此外,其他植物物种种子萌发后,与东非狼尾草生态位重叠较大,而在东非狼尾草极强侵占性下也难以获得竞争优势。因此,改良草地由于东非狼尾草侵占性强而导致其植物多样性降低。
已有研究表明:东非狼尾草和白三叶混播可提高牧草产量和品质[20-22],东非狼尾草粗纤维含量低但饲用价值较高[4]。本研究中,东非狼尾草改良草地可提高草地牧草和根系生物量及牧草氮和磷含量,降低牧草NDF 和ADF 含量。两者结果类似。
群落稳定性是在干扰活动、环境压力以及种间相容性3 个因子影响下,草地维持牧草组分稳定、草地生产力和系统功能基本不变的能力[23]。本研究中,改良草地群落整体处于相对稳定状态。其原因是:一方面,东非狼尾草是一种极耐践踏、耐旱涝和耐虫害性牧草[11,18],故以东非狼尾草为主导的草地难以受到人为以及环境因素干扰,更为稳定;另一方面,可能由于东非狼尾草具极强侵占性,能与其共生的植物种较少,草地在合理利用下很快就会到达稳定阶段。
由于中国南方喀斯特石砾地貌地形复杂,土壤贫瘠,不适合机械操作,从而该区域草山草坡常规改良所用草种(多年生黑麦草+鸭茅+白三叶)存在建植效果差、草地不稳定和易退化的缺陷,该区域一直是中国南方喀斯特山区草地改良和草地稳定性维持的难点。东非狼尾草具有匍匐蔓延的生长特点,且侵占性强,在该区域利用东非狼尾草与鸭茅、多年生黑麦草和白三叶混播建植改良草山草坡,东非狼尾草易成为群落优势种[4],不仅能提高草地牧草产量和品质,还能提高其抗逆性[24],有效抑制杂草生长,进而提高草地群落稳定性[20-21]。因此,东非狼尾草在中国南方喀斯特石砾地改良草地中表现出较强的竞争性和适应性,具有较好的草山草坡改良效果,为中国南方喀斯特地貌石砾地天然草地或草山草坡改良提供一种新选择。
4 结论
经过东非狼尾草改良后的草地,降低了植物丰富度指数、Shannon-Wiener 指数、Simpson 指数和Pielou 均匀度指数;此外,提高了牧草生物量及优质牧草禾本科和豆科所占比例;也提高了牧草氮、磷含量,降低了牧草中性洗涤纤维、酸性洗涤纤维、纤维素、酸性洗涤木质素、酸不溶灰分及土壤有机质含量;根据植物物种Raunkiaer频度图显示,改良后草地更加稳定。东非狼尾草改良石砾地效果较好,可作为中国南方喀斯特地区草地优质改良方法。
责任编辑:何謦成
