水稻长日照开花因子Lfm1的图位克隆
2021-01-22陈冰陈国鑫张治国
陈冰 陈国鑫 张治国
(中国农业科学院生物技术研究所,北京100081)
水稻(Oryza sativaL.)是我国主要的粮食作物,提高水稻粮食产量是农业科研中最主要的目标之一[1]。抽穗期(开花时间)与地区适应性以及产量密切相关,是受遗传和环境影响的最大的性状之一[2]。适度地调节水稻材料的开花时间将有助于充分利用当地的温度和光照条件,进而提高水稻产量[3]。
水稻属于短日照开花植物,在长期驯化过程中,逐渐从短日照向长日照、低纬度向高纬度繁衍,水稻已进化出适应长日照条件下的开花调控途径。水稻成花素两个基因RFT1与Hd3a,在水稻中也是关系最近的同源基因,但是RFT1在长日照下起作用,而Hd3a却在短日照行使功能。目前,已鉴定一些长日照的开花调节因子SDG724、EHD4、DTH2等。SDG724编码组蛋白甲基转移酶,通过调控MADS50和RFT1位点的H3K36me2/3甲基化水平,影响“MADS50/MADS51-Ehd1-Hd3a/RFT1”途径,促进水稻开花[4]。Ehd4基因编码一个水稻特有的CCCH类锌指蛋白,该基因通过诱导Ehd1调节成花素基因Hd3a和RFT1的表达从而控制抽穗[5]。DTH2是一个位于水稻第2染色体的在长日照下促进水稻抽穗的微效QTL,编码一个CONSTANS类似蛋白,其表达受时钟节律调控,DTH2 通过诱导成花素基因Hd3a和RFT1的表达而促进水稻抽穗,且独立于已知的Hd1和Ehd1起作用[6]。值得一提的是,突变体rid1在中日照、短日照和长日照条件下均不开花,始终停留在营养生长阶段,Rid1基因编码Indeterminate domain(IDD)转录因子成员[7]。虽然鉴定了一些影响水稻长日照的开花基因如SDG724、RFT1、EHD4、DTH2,但是对挖掘水稻长日照开花基因还十分有限。
本课题组前期开展了长日照条件下光周期突变体的筛选,获得了一批与长日照相关的突变体。本研究以一份长日照条件开花延迟的突变体lfm1为材料,利用图位克隆的方法克隆Lfm1基因,探讨了Lfm1基因在生产应用中可能存在的潜力,为培育适应不同生态区域的水稻品种提供参考。
1 材料与方法
1.1 材料
突变体lfm1材料来源于本实验室前期创制的以日本晴为背景水稻突变体库,突变性状稳定遗传且高代纯合[8]。材料种植于海南陵水和河北廊坊基地,用于统计材料的生育期。PCR反应使用的聚合酶是南京诺维赞公司的2×Rapid Taq Master Mix(P222-AA)。分子标记由北京擎科新业生物技术有限公司合成。
1.2 方法
1.2.1 遗传分析与群体构建 对突变体lfm1和野生型日本晴进行正反交,获得F1代种子,并统计F2后代生育期,明确控制突变体lfm1表型的遗传规律。同时以突变体lfm1母本,以籼稻广亲和材料Dular为父本进行杂交,配制基因定位群体。遗传分析与群体构建在海南陵水和河北廊坊实验基地进行。
1.2.2 水稻基因组DNA提取与PCR体系 日本晴、Dular、F2代群体的叶片基因组DNA提取参照改进的CTAB法[9]。采用10 μL反应体系:2×Rapid Taq Master Mix 5 μL,10 μmol/L正反向引物各0.5 μL,模板DNA 50 ng,ddH2O 3 μL。PCR反应程序:95℃预变性3 min;95℃变性20 s,60℃退火20 s,72℃延伸30 s,30个循环;72℃再延伸5 min。根据PCR产物差异大小,选择4%的琼脂糖凝胶(170 V,15 min)或者10%的聚丙烯胺凝胶(200 V,120 min)跑胶。
1.2.3 分子标记设计与基因定位 根据水稻籼粳亚种间基因组插入与缺失差异,在Gramene网站(http://www.gramene.org)上进行粳稻品种日本晴与籼稻品种9311基因组序列比对,寻找两亚种间存在插入与缺失变异的片段,利用Primer3web(http://primer3.ut.ee/)在线设计引物,产物大小在100-200 bp左右。开发的初定Indel标记共296对,实际可用Indel标记157对,平均每隔2-3 M覆盖水稻12条染色体。初定位用4个混池基因组DNA,每个混池有3个单株基因组DNA。同时提取200个F2代隐形表型单株基因组DNA用于精细定位和图位克隆。
1.2.4 定位引物序列 设计引物,对候选区间Lfm1基因进行定位,定位引物如表1。
2 结果
2.1 长日照晚花突变体lfm1的发现
2015年5 月至10月在河北廊坊实验基地种植水稻突变体库(光照为长日照,容量10000份),在材料收获时,发现一批在长日照下开花期有差异的材料。其中一个株系出现生育期延长现象,为了研究方便,后续将其命名为lfm1(图1-A-C),将突变体lfm1移栽至大棚中(防止降温对育性影响),~250 d后突变体才能正常结实(图1-D)。为了进一步判断突变体lfm1是否受短日照条件影响,我们将其种植于海南陵水基地(2015年12月-2016年4月生长),在短日照条件下,突变体lfm1的花期与野生型差异不太显著,约95 d可开花结实(图1-A、E)。

表1 引物序列
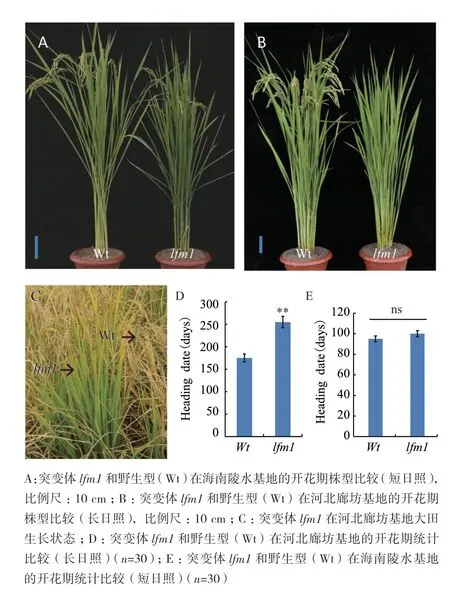
图1 突变体lfm1开花期表型鉴定
2.2 突变体lfm1遗传分析
由于突变体lfm1(生育期超长)在廊坊基地无法进行杂交,我们在海南基地对其进行了杂交配组,将lfm1与日本晴进行正反杂交,将获取的F1代种植于廊坊基地(长日照条件),F1代均表现为正常开花表型(生育期~170 d),F2代出现晚开花和正常开花分离表型,对正反交植株按3∶1分离比进行了卡平方检测,卡平方检测值均小于Х20.05(1)=3.84(表2)。上述F1和F2代的遗传分析表明,控制突变体lfm1在长日照下晚开花的表型受一对单隐性核基因控制,为下一步进行图位克隆打下基础。

表2 F2分离群体统计结果
2.3 Lfm1基因定位
为了定位Lfm1基因,在海南陵水实验基地,配制突变体lfm1与籼稻材料Dular杂交组合,获得大容量的F2代杂交种子,用于后续的定位试验。首先,我们开发了一套分布在水稻12条染色体的180对具有多态性的Indel标记,平均每隔3 M。用这套引物对Lfm1进行初定位,发现该基因与标记8-0.121(图2-A)和标记8-3.512(图2-B)紧密连锁,位于8号染色体的端粒附近(图3-A)。进一步选取了19个单株,初步将Lfm1定位在8号染色体0.121-3.512M之间。
2.4 Lfm1基因的克隆与突变区间分析
利用200个F2代隐形单株对Lfm1基因进行精细定位(图3-B),最终将Lfm1基因定位区间缩小到分子标记8-0.269和与8-0.283之间,范围为12 kb,其中包括3个基因(LOC_08g01410,LOC_08g01420,LOC_08g01430)(图3-C,表2)。然后对突变体lfm1中这3个候选基因进行测序,发现LOC_Os08g01420基因的第六外显子有9 bp缺失(图3-D)。经NCBI比对,发现LOC_Os08g01420基因编码一个含有植物同源结构域的锌指蛋白,是水稻开花的关键促进因子。经检索水稻已报道ehd3突变体,突变位点LOC_Os08g01420处第六外显子缺失11 bp,其生育期比对照显著延长。突变体lfm1等位于突变体ehd3(图3-D)[10]。长日照条件下,对一些开花基因进行了定量分析,结果表明,在lfm1突变体中,Ehd1、Hd3a和RFT1基因表达水平显著下降,Ghd7转录水平提高,而Rid1和Hd1基因没有变化,Ehd3可能依赖于Ehd1基因光周期遗传途径促进水稻抽穗。

图2 Lfm1基因定位连锁分析

图3 基因Lfm1的图位克隆
2.5 突变体lfm1的潜在育种价值
Lfm1是一个促进植物在长日照条件下开花的正因子,由于突变体lfm1在廊坊开花较晚,我们尝试其在低纬度条件(长沙)进行生长,并进行评估,在长沙,lfm1的生育比对照增加了20 d(对照日本晴170 d,相对应对照增加190 d)。因此,突变体lfm1穗粒数有显著提高(图4-A-B),突变体lfm1结实率和千粒重与野生相比差别不显著(图4-C-D)。但是显示突变体lfm1在适应的生态区具有巨大的生产应用潜力。
3 讨论
开花期是育种家和科研工作者最关注的农艺性状之一。合理的开花期对品种的选育和栽培至关重要。一般,在中国北方,需要选育开花期早的材料,而在南方,需要选育开花期适度晚的材料。水稻开花期受多个数量性状基因影响,因此基因比较难以克隆,第一个控制水稻开花期的基因Hd1就是利用重组自交系进行克隆的[11],也有一些利用染色体代换系进行克隆的[12-15]。而在本研究中,利用F2代材料就对Lfm1进行了成功克隆,有4点启示:第一,对开花期基因的克隆,需要选择后代分离3∶1的,这样就克服亲本背景材料对开花期基因的干扰。第二,对开花期基因的克隆,合理的种植条件也很重要,我们选择了群体在廊坊这个天然的长日照条件下进行生长。第三,定位亲本选择很重要,双亲的生育期差距不要超过10 d。第四,开花期材料如果差别很大,可以选择F2代进行定位克隆,如果差别不大,需要构建高世代材料进行定位。

图4 突变体lfm1与野生型在长沙(正季)的穗性状比较
通过图位克隆Lfm1基因发现,Lfm1基因编码一个含有植物同源结构域的锌指蛋白,是水稻开花的关键促进因子。利用RICEVAR2.0网站对Lfm1基因利用单倍型分析,发现在已有籼稻和粳稻资源材料中,存在多个SNP和INDEL变异,但是这些变异都发生在非编码区(内含子、5'-UTR、3'-UTR),表明LOC_Os08g01420是一个功能比较保守的基因,重要位点的突变如在编码区的突变会影响水稻的生长发育。同时对比突变体ehd3和突变体lfm1的表型和突变方式,突变体ehd3不仅表现为生育期延迟,还比较弱势生长特点[9]。对比突变方式,发现突变体ehd3是一个强突变体,可以用于后续的基因功能分析或作为背景材料的遗传资源。而突变体lfm1虽然突变也发生LOC_Os08g01420的第六外显子,其主要表型为生育期延迟,而无其它不利表型,因此突变体lfm1可能是一个LOC_Os08g01420基因的弱突变体材料。经在长沙种植,发现突变体lfm1的生育期比对照增加了20 d,由于生育期延迟,造成光合产物进一步积累,因此,突变体lfm1穗粒数有显著提高,这个结果暗示突变体lfm1在适应的生态区具有潜在的生产应用潜力。将来,由于突变体lfm1不是转基因材料,可以将其通过回交转育聚合到不同生育期的水稻主栽材料中,培育适合不同地域的育种新材料。
4 结论
本研究通过筛选水稻突变体库,获得一份在长日照条件下晚开花的材料lfm1,其在短日照条件下能够正常开花,通过图位克隆,发现在突变体lfm1中,LOC_Os08g01420基因的第六外显子2800处缺失9个碱基,突变体lfm1等位于突变体ehd3。并发现在适度的光照条件下,突变体lfm1表现为穗粒数增多,生育期略延长,可能存在潜在的生产应用潜力。
