酿酒酵母酚类抑制物耐受性脂质组学研究
2021-01-22顾翰琦邵玲智刘冉刘晓光李玲刘倩李洁张雅丽
顾翰琦 邵玲智 刘冉 刘晓光 李玲 刘倩 李洁 张雅丽
(1. 河北民族师范学院生物与食品科学系,承德 067000;2. 河北琢酒集团有限公司,承德 067600)
木质纤维素具有环境友好性和可再生性的特点,是纤维素乙醇最具发展潜力的生产原料[1]。但在利用木质纤维素原料之前需要进行预处理破坏其复杂致密的结构[2]。在预处理过程中会产生酚类、呋喃类和有机弱酸类毒性物质,这对微生物细胞活性以及乙醇发酵具有很大的抑制作用。其中酚类物质的毒性最强,特别是酚酸抑制物种类多、浓度高,对纤维素乙醇生产过程产生严重影响[3]。因此有必要筛选或构建酚酸耐受性酿酒酵母,并对其耐受机制进行研究。
为应对木质纤维素预处理衍生抑制物的毒害作用,已有研究通过脱毒法、基因工程改造和适应进化等策略提高酵母细胞对抑制物的耐受性[4-5]。其中,适应进化方法是在无需相关遗传信息和分子作用机制的背景下,通过定向的适应和筛选能够有效获得耐受菌株的策略[6]。其具有能够在较短的时间内有效定向的改变菌株的某些表型或者生理特性并且基本不会影响目的表型以外的其他优良性状的优点。在前期研究中已经通过适应进化策略构建了酚酸耐受性酵母菌株,并且发现适应进化菌株在酚酸胁迫下能表现良好的细胞膜完整性[7],但是其耐受性提高的作用机制尚不明确。
细胞膜作为细胞与外界环境的屏障,对于维持细胞膜内环境的稳态、控制物质进入、信息传递及能量流动都有着重要作用[8]。在工业微生物抗逆性研究方面,已有大量研究从酵母细胞质膜组分代谢和调控应答、细胞膜信号途径、质膜抗逆基因等方面揭示酵母细胞质膜抗逆应答机制[9]。当外界环境中存在抑制物、高渗、低渗、pH等胁迫因素时,首先对细胞膜造成严重的破坏。研究发现木质纤维素衍生的酚酸化合物对发酵微生物抑制作用主要因其具有疏水性芳香环,从而容易进入细胞膜并破坏膜的完整性[7,10]。巩林林等[11]研究发现乙腈会溶解细胞膜表面脂质,增加细胞膜的流动性,导致酵母细胞膜部分出现裂痕,从而增加致死率。郭红等[12]研究表明极端高糖产生的高渗胁迫下通过干预脂肪酸的组成和含量影响细胞膜的流动性和渗透性,从而提高细胞对高渗的耐受性。其他多项研究表明脂肪酸饱和度和酰基长度的变化可以维持细胞膜的流动性,提高酵母细胞对乙醇的耐受性[8]。Tian等[13]研究发现通过降低细胞中饱和脂肪酸和短链脂肪酸含量,同时提高麦角固醇含量能够抵御环境胁迫。上述研究说明压力胁迫下细胞膜的性质功能和结构组成变化对微生物抗逆性起到非常重要作用。
磷脂、甾醇和鞘脂作为酵母细胞膜的主要组成成分,对于维持细胞膜结构和生物学功能有重要作用。脂质组学研究方法作为代谢组学的一个分支主要对细胞内脂质代谢物进行检测、鉴定和系统的比较分析。本研究在适应进化构建酚酸耐受性酿酒酵母的基础上,通过脂质组学分析方法对酚酸胁迫下野生型和适应进化型菌株的脂质成分进行系统分析。主要从细胞膜磷脂的种类分布及其脂酰链结构变化等方面进行统计分析。并通过统计学分析方法筛选酚酸耐受性酵母的显著差异的磷脂成分。为细胞膜工程改造理性构建抗逆性菌株提供前期研究基础。
1 材料与方法
1.1 材料
1.1.1 菌种 酿酒酵母(Saccharomyces cerevisiae),购自安琪酵母股份有限公司。以其作为原始菌株经过混合酚酸适应进化得到耐受性菌株S. cerevisiaePAT01(能耐受导致酵母细胞产生50%生长抑制的酚酸浓度),保藏于中国微生物菌种保藏管理委员会普通微生物中心,编号CGMCC No.18021。
1.1.2 试剂 甲醇、乙腈为色谱纯(德国Merck公司);甲酸、L-2-氯-苯丙氨酸为色谱纯(美国Sigma-Aldrich公司);香草酸、对羟基苯甲酸、丁香酸为分析纯(上海阿拉丁化工有限公司);Yeast Extract为生物试剂(英国Oxoid公司);其他试剂均为分析纯,购自天津科密欧化学试剂公司。
1.1.3 培养基 合成培养基:20 g/L葡萄糖、2 g/L K2HPO4、1 g/L MgSO4、1 g/L(NH4)2SO4、10 g/L Yeast Extract溶于水后121℃高压灭菌20 min。混合酚酸培养基:合成培养基中加入适量香草酸、对羟基苯甲酸和丁香酸溶于二甲基亚砜的母液,使培养基中最终浓度分别1.44 g/L、0.87 g/L和0.72 g/L。PBS缓冲液:8.00 g/L NaCl,0.20 g/L KCl,1.42 g/L Na2HPO4,0.24 g /L KH2PO4,pH7.4。
1.2 方法
1.2.1 酵母细胞培养收集 将酿酒酵母野生型菌株WT和适应进化菌株PAT01分别在20 mL合成培养基中活化18 h,取适量菌液调节菌体密度至600 nm吸光值(OD600)为7-8,以10%(V/V)接种量转接至混合酚酸培养基中,培养6 h后收集菌体。2 mL菌液与等体积预冷的60%(V/V)甲醇溶液混合[10],4427×g,4oC离心10 min,弃上清。称取菌体沉淀(湿重)不少于200 mg,-80oC保存备用。参考相关脂质组学研究[14-15],确定每种菌株制备3个平行样品。
1.2.2 细胞脂质提取 称取酵母细胞样品200 mg,加入甲醇溶液(含5 μg/mL L-2-氯-苯丙氨酸作为内标)300 μL,在50 Hz条件下匀浆2 min,冰水浴超声20 min。在4℃,11167×g离心10 min,最后取200 μL上清液待测。
1.2.3 液相色谱-质谱(LC-MS)分析 使用Agilent 1290 Infinity和Agilent 6545 UHD Accurate-Mass Q-TOF系统(美国Agilent公司)进行LC-MS分析。液相色谱条件:色谱柱为Waters XSelect HSS T3(2.5 μm,100 mm×2.1 mm)(美国Waters公司),柱温35oC,流动相A为0.1%甲酸的水溶液,流动相B为0.1%甲酸的乙腈溶液,流速350 μL/min,梯度 洗 脱:0-2 min,5% B;2-10 min,5%-95% B;10-15 min,95% B;15-18 min,95%-5% B, 进样量2 μL;质谱条件:毛细管电压为3.5 kV,干燥气体流速10 L/min,温度325℃,喷雾器压力为20 psig。正、负离子模式下质谱的扫描范围分别为100-3000 m/z和100-1700 m/z。
1.2.4 数据分析 LC-MS数据处理:将LC-MS检测的原始数据通过Agilent Masshunter Qualitative Analysis B.08.00软件(美国Agilent公司)转换成mz.data格式。然后使用XCMS 3.2.0程序对数据进行峰识别、保留时间校正、峰匹配等操作,得到包含m/z、RT和样品特征信号强度匹配信息的.csv格式数据包。将数据包通过LipidFinder在线数据库进行比对,获得样品脂质信息,然后对上述数据进行内标归一化处理。最后通过正交偏最小二乘判别分析(OPLSDA)方法筛选WT和PAT01两酵母菌株显著性差异脂质分子。
2 结果
2.1 酵母细胞主要脂质成分分析
脂质分子是细胞膜的酵母细胞膜组成成分,不同种类脂质分子结构和理化性质存在明显差异直接影响细胞膜的结构和功能。因此,通过LC-MS方法对酿酒酵母WT和PAT01两菌株脂质成分进行检测,进一步分析适应进化酵母对酚酸耐受性的分子机制。结果如图1所示,同一菌株的平行样品在正、负离子模式总离子流谱图中的色谱峰信号强度和保留时间均表现良好的重现性。而两菌株在特定保留时间区域的峰信号强度存在明显差异。这说明检测系统稳定,所得数据可靠,而且两菌株脂质成分含量存在差异。进一步对两菌株脂质成分进行鉴定和统计分析,结果如图2所示。WT和PAT01菌株在正、负离子模式下共检测到565种脂质分子,所有脂质分子共涉及21种亚类,分别归属于甾醇(Ste、C24-Pr-S、Sec、Stigma、Ste-con、Erg、Other-ST),鞘脂(Cho、Sul、Other-SP、N-GLS、Cer、PSL、Sph、GM)和磷脂(PA、PC、PE、PG、PI、PS)三大类脂质。其中,磷脂的种类最多,占总脂质种类的80%以上,这与酵母细胞膜中磷脂含量基本一致[16]。
2.2 酵母细胞膜磷脂组成分析
磷脂双分子层作为细胞膜的基本支架,磷脂的亲水头部基团种类和疏水尾脂酰链的长度与饱和度等对细胞膜的性质和功能起决定作用[17]。因此,进一步从以上3个方面对WT和PAT01菌株细胞膜磷脂分布进行比较分析。主要包括:磷脂酸(PA)、磷脂酰胆碱(PC)、磷脂酰乙醇胺(PE)、磷脂酰甘油(PG)、磷脂酰肌醇(PI)和磷脂酰丝氨酸(PS)。结果如图3所示,不同头部基团种类的磷脂分布方面,WT菌株中PE、PG和PS相对含量较高,含量分别超过20%,而酚酸耐受的进化菌株PAT01中相对含量最高的磷脂分别为PE、PC和PI,比WT中相应磷脂增加1.8、3.8和2.22倍(图3-a)。根据脂酰链不饱和键数量对磷脂分布统计,WT菌株中含饱和脂酰链、一个不饱和键和两个不饱和键脂酰链的磷脂含量分布相对平均,相对含量均在30-40%。而适应进化菌株PAT01中含两个不饱和键的磷脂含量明显增加,占总磷脂含量的70%以上(图3-b)。进一步根据磷脂脂酰链碳的原子数量统计含有不同脂酰链长度的磷脂分布,WT菌株磷脂脂酰链长度主要分布在C28、C32、C34和C36,另外少量分布在超长链C40以上,其中C28的磷脂含量最高。而PAT01菌株集中分布在含C32、C34和C36的磷脂组分,大部分由C16、C18和C20的长链不饱和脂肪酸构成(图3-c)。结果表明,适应进化酵母细胞膜磷脂发生了明显的重塑现象。
2.3 显著性差异磷脂成分分析
利用正交偏最小二乘判别对WT和PAT01菌株的脂质测数据进行统计学分析(OPLS-DA),筛选对适应进化菌株酚酸耐受性起重要作用的显著差异磷脂标志物。结果如图4所示,OPLS-DA得分结果表明WT和PAT01菌株组间具有明显的分离趋势,x轴第一主成分t[1]反映了组间差异最大化,y轴to[1]正交主成分表现出两组数据的组内变异情况(图4-a-b)。Permutation检验图用于反映模型的可靠性,其中R2代表模型解释率,Q2代表模型预测率,数值越大说明模型具有更好的解释和预测能力。正离子模式下两组累积R2Y=0.99,Q2=0.82;负离子模式累积R2Y=1.00,Q2=0.90(图4-c-d)。该模型的S-plot图中,分布在右上角和左下角离原点越远的点表明对组间差异的贡献度越大,模型得到的变量权重值(VIP)也越大,这些点代表WT和PAT01两菌细胞中存在显著差异的脂质分子(图4-e-f)。
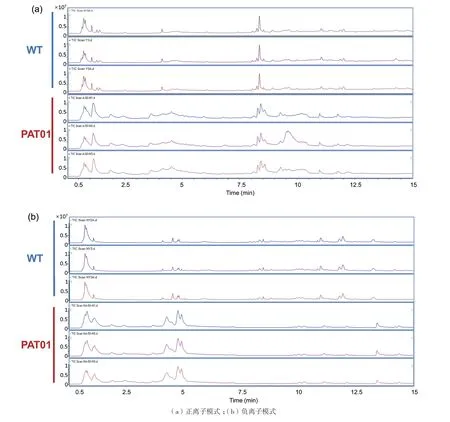
图1 酚酸胁迫下酿酒酵母脂质成分LC-MS分析总离子流图(TIC)

图2 酚酸抑制物作用下酿酒酵母脂质种类分布

图3 酚酸胁迫下酵母菌株细胞膜磷脂相对含量分布
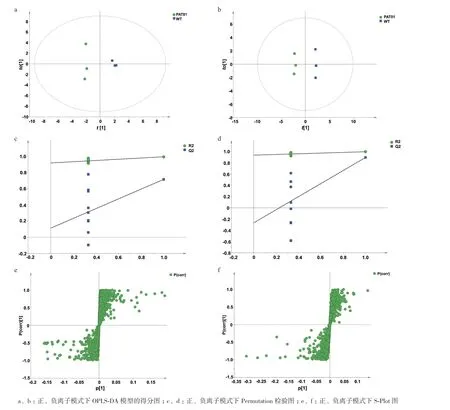
图4 酚酸适应进化菌株脂质正负离子模式下OPLS-DA模型分析
根据OPLS-DA模型的VIP值(≥1),并结合独立样本t检验P值(<0.05)以及差异倍数(FC>2或<0.5)作为标准,进一步筛选WT与PAT01菌株之间的显著性差异脂质代谢物[18]。FC代表两菌株中不同磷脂分子相对含量的比值,以2为底取对数得到log2FC,可以直观判断不同磷脂分子相对含量的变化情况,log2FC>0,表示磷脂相对含量增加,反之降低[19]。结果如表1所示,两菌株显著性差异变化的磷脂种类主要集中在PC和PE。PAT01菌株相比WT菌株,PC类中含有短脂酰链、饱和脂酰链的磷脂分子,以及只含有单个脂酰链的溶血磷脂酰胆碱(LPC)相对含量减少(LPC 4:0、LPC 5:0、LPC 14:0),而PC和PE含有长链和不饱和脂酰链的磷脂相对含量增加(PC 34:2、PC 35:2、PC 36:1、PC 36:2、PE 34:2、PE 36:2、PE 40:2)。这与上述磷脂组成分布统计结果一致,说明适应进化菌株PAT01对酚酸耐受性提高可能是由于细胞膜磷脂重塑导致,特别是PC和PE磷脂分子结构和分布变化起重要作用。
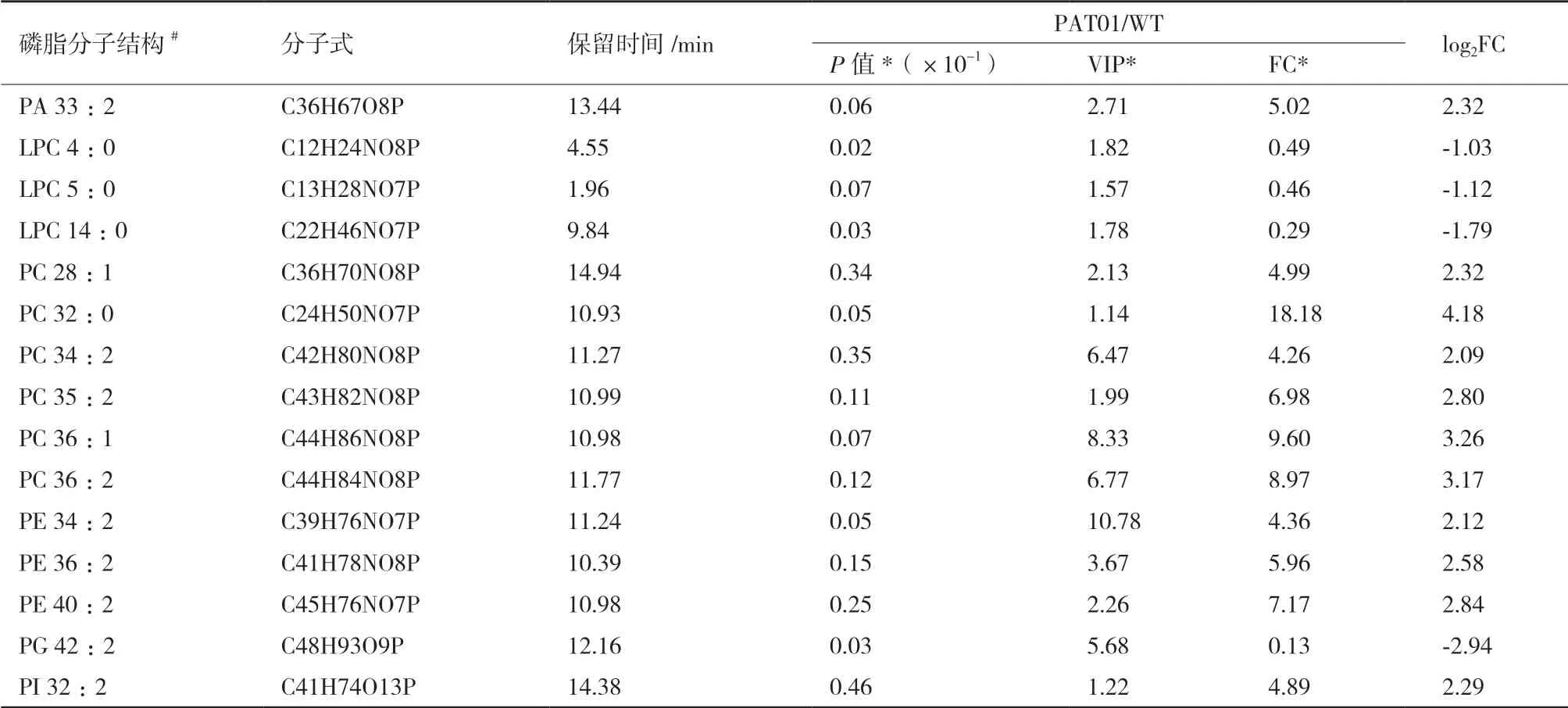
表1 PAT01/WT显著性差异磷脂分子
3 讨论
木质素降解产生酚酸化合物对发酵微生物抑制作用机制主要表现在其疏水性芳香环能够进入细胞膜并破坏其完整性,同时酚酸进入细胞后解离释放质子,从而导致细胞内物质外泄、ROS增加、pH紊乱及严重的细胞自噬等。我们前期研究发现通过酚酸适应进化的酿酒酵母菌株PAT01与初始菌株WT相比,在酚酸作用下能够保持更高的细胞膜完整性和较低的渗透性。细胞膜的这些性质与其磷脂成分的头部亲水基团种类和尾部脂酰链结构密切相关。本研究通过脂质组学分析方法对酿酒酵母细胞膜磷脂成分进行系统分析,结果发现对酚酸有耐受性的进化菌株磷脂种类分布和脂酰链结构发生明显变化,其显著性差异的磷脂分子主要是PC和PE类含长链不饱和脂酰链的磷脂分子。首先,进化菌株PAT01与WT菌株相比,PE、PC和PI相对含量增加。PC和PE是细胞膜中主要的磷脂成分,说明进化菌株PAT01在酚酸作用下仍能保持相对正常的细胞膜组分,而WT菌株则可能由于细胞膜被破坏导致磷脂成分分布发生明显变化。PE和PC合成代谢之间存在密切联系,PE主要在线粒体中合成,部分运送到内质网经过连续甲基化转化为PC,因此PE的含量直接影响PC的合成。酵母细胞PC和PE相对含量对调节细胞膜曲率、维持细胞膜的稳定方面起重要作用。PE由于头部乙醇胺基团很小呈圆锥形,自由组装成膜时容易形成负向弯曲,而PC头部基团为带有3个甲基乙醇胺基团,相对较大,与脂酰链形成圆柱形,趋于形成平行的双层膜。因此,两种磷脂的分布比例能够有效调节膜的曲率和横向应力,从而保持膜稳定[17]。已有研究表明,酿酒酵母在含有木质纤维素衍生抑制物乙酸、糠醛和苯酚压力胁迫下,PE/PC比例明显增加,有效提高了细胞膜完整性并降低了细胞膜通透性,从而提高酵母对抑制物的耐受性[20]。另外,真核细胞中PE是调节细胞膜流动性的关键因素,PE的头部基团能够与附近磷脂分子的氨基头部基团和磷酸残基形成氢键,因此PE含量增加能够有效提高膜稳定性和刚性[17,21-22],这有助于提高膜对外界毒性分子的屏蔽作用。已有研究报道,酿酒酵母在含有高浓度镉、锌、铁或铝离子的环境中培养时,细胞磷脂含量明显增加,其中PE含量增加最显著,从而维持细胞膜稳态[23-24]。此外,由于磷脂头部基团所带电荷不同,PC和PE均属于两性离子类磷脂,而其余PA、PG、PI和PS都属于阴离子类磷脂,细胞膜中阴离子类与两性离子类磷脂的比例对膜电势又调控作用,增加膜电势有助于细胞去极化从而使细胞具有更大的柔韧性,对环境压力不敏感。已有研究报道,阴离子类与两性离子类磷脂的比值下降时酿酒酵母细胞对盐离子表现出更强的耐受能力[25]。此外,另一种含量明显增加的磷脂PI与细胞活性有关,在调控细胞生长功能等方面起重要作用[13]。PI相对含量增加有助于提高酿酒酵母对木质纤维素预处理抑制物耐受性。已有研究表明,含有长脂酰链的PI对提高细胞膜流动性和降低渗透性有重要作用[20]。通过补充PI的生物合成前体物质肌醇能够有效改善酿酒酵母对木质纤维素衍生抑制物耐受性[26]。
另一方面,磷脂尾部脂酰链结构对膜的性质功能同样起到重要作用。已有大量研究通过基因工程改造调控细胞膜的饱和与不饱和脂肪酸比例或降低短链脂肪酸含量等策略应对细胞膜损伤问题[27]。饱和脂酰链在生理温度下趋于形成不流动紧密结合的凝胶状态,而不饱和脂酰链使膜具有流动性[28]。有研究证明增加不饱和脂肪酸含量不仅能提高酿酒酵母对辛酸的耐受性而且减轻膜渗漏[29]。通过过表达脂肪酸去饱和酶(ELO1和OLE1)能够有效提高油酸含量,增加膜稳定性,进一步提高酵母对乙酸的耐受性[30-31]。脂酰链的长度与磷脂双分子层的厚度呈正相关,细胞膜厚度增加从而增加外界有毒物质跨膜的行程和阻力,从而降低膜的通透性。有研究通过在酿酒酵母细胞过表达脂酰链酰基延长酶能够提高硬脂酸和油酸含量以及平均链长度,从而使其细胞膜完整性增强[27]。
4 结论
通过对酿酒酵母野生型菌株WT与酚酸耐受性适应进化菌株PAT01进行脂质组学分析,发现PE、PC和PI相对含量明显增加,含有长链脂酰链的磷脂成分以及含有两个不饱和脂酰链的磷脂成分相对增加。据此推断适应进化酵母菌株对酚酸耐受性提高的机制在于细胞膜发生磷脂成分重塑,有效提高细胞膜完整性,使细胞膜在酚酸压力胁迫下仍能保持稳态,从而对酚酸抑制物起到选择性屏障作用。
