茶足柄瘤蚜茧蜂蛹滞育过程中胰岛素信号通路及其相关途径的初探
2021-01-18韩海斌刘爱萍高书晶徐林波黄海广
刘 敏,韩海斌,刘爱萍*,高书晶,徐林波,黄海广
(1. 中国农业科学院草原研究所,呼和浩特 010000;2. 内蒙古自治区林业科学研究院,呼和浩特 010000)
茶足柄瘤蚜茧蜂LysiphlebustestaceipesCresson,隶属膜翅目Hymenoptera蚜茧蜂科Aphidiidea,是一种寄生性昆虫,可寄生于多种蚜虫,包括麦二叉蚜SchizaphisgraminumRondani、棉蚜AphisgossypiiGlover、玉米蚜RhopalosiphummaidisFitch、大豆蚜AphisgylcniesMatsumura等经济作物上的重要害虫和危害豆科牧草及沙生植物的主要蚜虫-苜蓿蚜AphiscraccivoraKoch(Rodrigues and Bueno, 2001; Silvaetal., 2008; 刘兴龙等,2009),是优良的天敌昆虫。
在生物体内,胰岛素信号通路(Insulin signaling pathway)是一条非常重要的信号通路,通过调节糖代谢、脂代谢和蛋白质代谢,从而影响生物的生长、代谢、生殖、衰老等过程(Wu and Brown, 2006)。在昆虫体内,存在与脊椎动物胰岛素序列相似的蛋白,通常命名为胰岛素样蛋白(Insulin-like peptides, ILPs)。除埃及伊蚊AedesaegyptiILP3外,昆虫胰岛素样蛋白只有一个受体-胰岛素受体(Insulin receptor,INSR或IR)(Wenetal., 2010)。
胰岛素样蛋白与胰岛素受体结合后,胰岛素信号启动,激活的受体紧接着磷酸化胰岛素受体底物(Insulin receptor substrate,IRS);继而激活两条主要通路,第一条通路是通过磷脂酰肌醇-3-激酶(phosphatidylisonsitol 3-kinase,PI3K),激活蛋白激酶B(protein kinase B,PKB/Akt),蛋白激酶B磷酸化下游的靶标蛋白,如叉头状转录因子O家族FoxO(Forkhead box-containing protein, FoxO),来达到调节的目的,从而影响生命体的代谢调控过程,如糖代谢、脂类合成和基因表达等(Frame and Cohen, 2001; Tzivion and Hay, 2011; Leto and Saltiel, 2012)。FoxO转录调控因子是胰岛素信号通路中的关键分子,无刺激因子时,FoxO停留在细胞核中,处于去磷酸化状态,细胞核中的FoxO转录因子可以与相关的靶基因结合从而影响细胞周期、细胞凋亡、延长寿命、新陈代谢和应激反应、能量需求等(Accili and Arden, 2004)如果FoxO不能完成磷酸化,则会影响昆虫生长甚至导致细胞死亡(Rhodes and White, 2002);第二条通路是通过Ras(rat sarcoma, Ras)调控有丝分裂原激活的蛋白激酶(mitogen-activated protein kinase,MAPK)信号路径(Taniguchietal., 2006;Slacketal., 2015),最终调控细胞内的蛋白合成和细胞增殖分化(Cantley, 2002)。
滞育是昆虫的一种生命现象,具体表现为在特定时期发育停滞,不食不动,新陈代谢降低,是昆虫在漫长进化中长期适应不良自然环境而获得的一种生存本领(徐卫华,1994)。滞育的生理指标不仅仅表现在发育停止,也体现在行为方式转变、代谢抑制以及对逆境抵抗力增强(And and Tauber,1976;Denlinger, 2002)。通过诱导茶足柄瘤蚜茧蜂滞育,可以延长其产品的货架期。滞育的分子机理十分复杂,这一生理现象是众多信号通路协同作用而形成的复杂网络,接收来自感觉神经元的各种环境信息,如温度、光照、水分、食料等,再通过信号传导作用于各组织器官而影响代谢、生殖、抗性等,最终调控滞育的形成和维持。
胰岛素信号通路和FoxO是控制滞育的关键,影响昆虫的代谢和脂质储存,使卵巢发育停滞,提高越冬存活率。在短日照下,胰岛素信号通路被关闭,并且解除对下游FoxO的抑制,从而导致脂质的积累,呈现出滞育状态,胰岛素信号通路在其他休眠形式中也表现出相同的作用机制(安涛,2015)。
在双翅目昆虫果蝇Drosophilamelanogaster中,胰岛素信号通路异常可致使成虫滞育(Sim and Denlinger, 2013)。将哺乳动物胰岛素注射给欧洲粉蝶Pierisbrassicae(Arpagaus, 1987)的蛹后,粉蝶滞育终止并恢复正常发育。敲除果蝇PI3K(Dp110)后,果蝇滞育率升高,而在神经系统中过表达Dp110,滞育率会呈现下降趋势。滞育的雌尖音库蚊Culexpipiens体内储备脂肪大量,卵巢停止发育;而干扰未滞育雌蚊的胰岛素受体,其卵巢也停止发育;胰岛素样蛋白1和胰岛素样蛋白5在将要滞育的雌蚊中的表达量比雄性低,敲除胰岛素样蛋白1,未滞育雌蚊卵巢停止发育(Williams and Sokolowski, 2009)。这些研究表明,未激活的胰岛素信号通路可引发滞育,激活后滞育终止或缓解。
由此可以看出,胰岛素信号通路在滞育进程中起到关键作用,茶足柄瘤蚜茧蜂蛹滞育的研究中,胰岛素信号通路及其相关途径起到什么作用,是否与已研究的结果相符,仍有待探讨。本实验通过对茶足柄瘤蚜茧蜂滞育蛹与非滞育蛹进行转录组测序,重点研究与胰岛素信号通路及其相关途径有关的基因在滞育状态与非滞育状态下表达量的差别,从而探索胰岛素信号通路及其相关途径在茶足柄瘤蚜茧蜂滞育过程中所起的作用,旨在为茶足柄瘤蚜茧蜂等小型寄生蜂提供滞育机理研究的方向,为害虫防治提供新思路。在实际生产应用中,可以通过寻找胰岛素替代物来取代传统通过温光等环境条件诱导昆虫滞育的方式,来控制寄生蜂滞育,从而更高效地应用天敌昆虫防治害虫。
1 材料与方法
1.1 供试虫源、供试寄主植物与滞育诱导
茶足柄瘤蚜茧蜂、寄主蚜虫苜蓿蚜采自中国农业科学院草原研究所沙尔沁基地,供试寄主植物为蚕豆Viciafaba。
苜蓿蚜采自基地的羊柴Hedysarummongolicum植株上,并转接在室内的水培蚕豆苗上繁殖,接虫后对蚕豆苗进行笼罩(100目防虫网笼,55 cm×55 cm×55 cm),确保苜蓿蚜未被天敌寄生,试验用2~3龄的苜蓿蚜若蚜作为寄主,在温室内饲养5代以上作为供试虫源。
从基地采集被寄生的苜蓿蚜僵蚜,从中挑取未羽化破壳的僵蚜置于人工气候箱,温度25±1℃,相对湿度70%±1%,光周期L ∶D=14 h ∶10 h条件下培养,待蜂羽化后,挑选茶足柄瘤蚜茧蜂转移至试管(10 cm×3 cm)内,用20%蜂蜜水作为补充营养,接入具有苜蓿蚜的蚕豆苗上,建立茶足柄瘤蚜茧蜂种群作为供试虫源,并在室温下用苜蓿蚜有效扩繁10代以上。
在室温下养虫笼中将刚羽化成蜂按1 ∶100的蜂蚜比释放成对茶足柄瘤蚜茧蜂,根据实验室前期研究基础可知,苜蓿蚜若蚜被茶足柄瘤蚜茧蜂寄生后,寄生蜂卵继续发育120 h,此时僵蚜体内寄生蜂处于高龄幼虫(3~4龄)阶段,高龄幼虫为茶足柄瘤蚜茧蜂感受滞育信号的敏感虫态,将此时的僵蚜放入人工气候箱中进行滞育诱导。高龄幼虫处于滞育环境条件时,并不会立刻停止发育,而是继续发育一段时间,经试验验证,当发育至蛹时,便不再继续发育(孙程鹏,2018)。本实验中,诱导茶足柄瘤蚜茧蜂滞育的温光组合为温度8℃、光周期L ∶D=8 h ∶16 h,诱导时长为30 d。选取经过30 d滞育诱导的僵蚜进行解剖,选择虫态是蛹的茶足柄瘤蚜茧蜂进行收集,以获得滞育组样品;苜蓿蚜若蚜被茶足柄瘤蚜茧蜂寄生后,放置在25±0.5℃、RH 70%±5%、光周期L ∶D=14 h ∶10 h、光照强度8 800 Lx(人工气候箱,上海一恒公司MGC-HP系列)条件下,寄生蜂卵继续发育168 h(此时蚜茧蜂处于蛹态),对僵蚜进行解剖,挑选饱满、有活力的蛹作为正常发育组样品。
1.2 测序和组装
茶足柄瘤蚜茧蜂滞育组与非滞育组的转录组测序工作由北京诺禾致源科技股份有限公司完成。测序进行3个生物学重复,共构建6个文库。测序完成后,经数据预处理,采用Trinity(Grabherretal., 2011)对clean reads进行拼接。
1.3 序列筛选
1) 拼接得到的茶足柄瘤蚜茧蜂转录组测序结果与Nr数据库进行Blastx比对(将核酸序列翻译为蛋白,再进行比对),筛选条件E-value<1e-5,得到注释基因。
2) 应用每个样本的RPKM (reads per kilo base of exon model per million mapped reads),以每个样本的总表达量为内标进行Fisher-test差异检验,将正常发育组和滞育组转录组结果进行比较,采用FDR与表达差异倍数Fold-Change方法进行差异基因挑选,筛选条件为:FDR≤0.05,Fold-change≥2。
3) 对筛选到的差异基因进行序列提取。
1.4 功能注释
将筛选到的序列与KEGG数据库进行比对。KEGG(Kyoto Encyclopedia of Genes and Genomes)是有关Pathway的主要公共数据库(Kanehisa, 2008)。Pathway显著性富集分析以KEGG Pathway为单位,应用超几何检验,找出差异基因相对于所有有注释的基因显著富集的pathway。该分析的计算公式:

在这里N为所有基因中具有pathway注释的基因数目;n为N中差异表达基因的数目;M为所有基因中注释为某特定Pathway的基因数目;m为注释为某特定Pathway的差异表达基因数目。FDR≤0.05的Pathway定义为在差异表达基因中显著富集的Pathway,使用KOBAS(2.0),设置参数—fdr为BH(即使用BH校正)进行Pathway富集分析。
2 结果与分析
对茶足柄瘤蚜茧蜂蛹进行转录组测序发现,KEGG注释到的与胰岛素信号通路相关的基因有64个,与该通路相关的滞育组与非滞育组差异表达基因有31个。胰岛素信号通路激活的重要途径有两条,分别是PI3K途径和MAPK途径,胰岛素信号主要通过这两条途径传递。胰岛素(INS)激活胰岛素受体(INSR),胰岛素受体磷酸化形成胰岛素受体底物(IRS),磷酸化的胰岛素受体底物可以激活PI3K和RAS,激活的物质不同,也决定了进入不同的信号传导途径。PI3K激活后,促使磷脂酰肌醇三磷酸(PIP3)的生成,而PIP3被认为是胰岛素的第二信使,PI3K与细胞内含有Pleckstrin Homology(PH)结构域的信号蛋白PDK1/2结合,继续磷酸化导致Akt活化,调节下游靶标蛋白(如FoxO, GSK3, GLUT等)影响糖代谢、脂类合成等。磷酸化的受体底物LAR间接结合并磷酸化含有src同源区2(SH2)结构域的蛋白质SHC后,再激活GRB2,GRB2与SOS结合使之活化,激活的SOS即可与膜上的Ras相结合。Ras使Raf定位于质膜,激活其活性,Raf结合并磷酸化MEK1/2使之激活,继续进行信号传导,调控MAPK通路,最终达到调控蛋白合成、细胞增殖分化的目的(图1)。分别有55个和28个差异表达基因与PI3K途径和MAPK途径相关,与胰岛素信号通路密切相关的FoxO信号通路,共有21个差异表达基因(表1),由于基因数量较大,根据前人的研究,挑选出与信号通路密切相关的部分基因进行分析。参与上述4条信号通路的基因有Sos,FASN,TSC1,JNK,Rac1,PRKAB,Akt,NLK等基因。在这些基因中,除PRKAB基因上调表达外,其他基因均下调表达。它们与茶足柄瘤蚜茧蜂滞育密切相关,共同影响茶足柄瘤蚜茧蜂蛹的滞育,主要体现在影响虫体能量代谢、脂质积累、细胞增殖等方面。

图1 差异基因在胰岛素信号通路富集的通路图Fig.1 Pathway map of differentially expressed genes enrichment in insulin signaling pathway注:包含上调基因的KO节点边框标红色,包含下调基因的KO节点边框标绿色,包含上下调的边框标黄色;实心箭头代表分子间的相互作用或关系,圆圈代表化学分子;方框-实心箭头-圆圈相连,代表表达;方框-实心箭头-竖线-圆圈相连,代表抑制;方框-虚线箭头-圆圈相连,代表间接作用;直线-方框-实心箭头-圆圈-直线-方框-实心箭头相连,代表两个连续的反应步骤;带箭头的直线上标有“+P”代表磷酸化,标有“-P”代表去磷酸化。Note:The KO node border containing the up-regulated genes is red, the KO node border containing the down-regulated genes is green, and the border containing the up-and-down regulated is yellow; a solid arrow represents an interaction or relationship between molecules and a circle represents a chemical molecule; box-solid arrow-circle linked, for expression; box-solid arrow-vertical line-circle linked, for inhibition; box-dashed arrow-circle linked, representing indirect action; line-box-solid arrow-circle-straight line-box-solid arrow linked, representing two consecutive reaction steps; lines with arrows are marked “+p” for phosphorylation and “-p”for dephosphorylation.
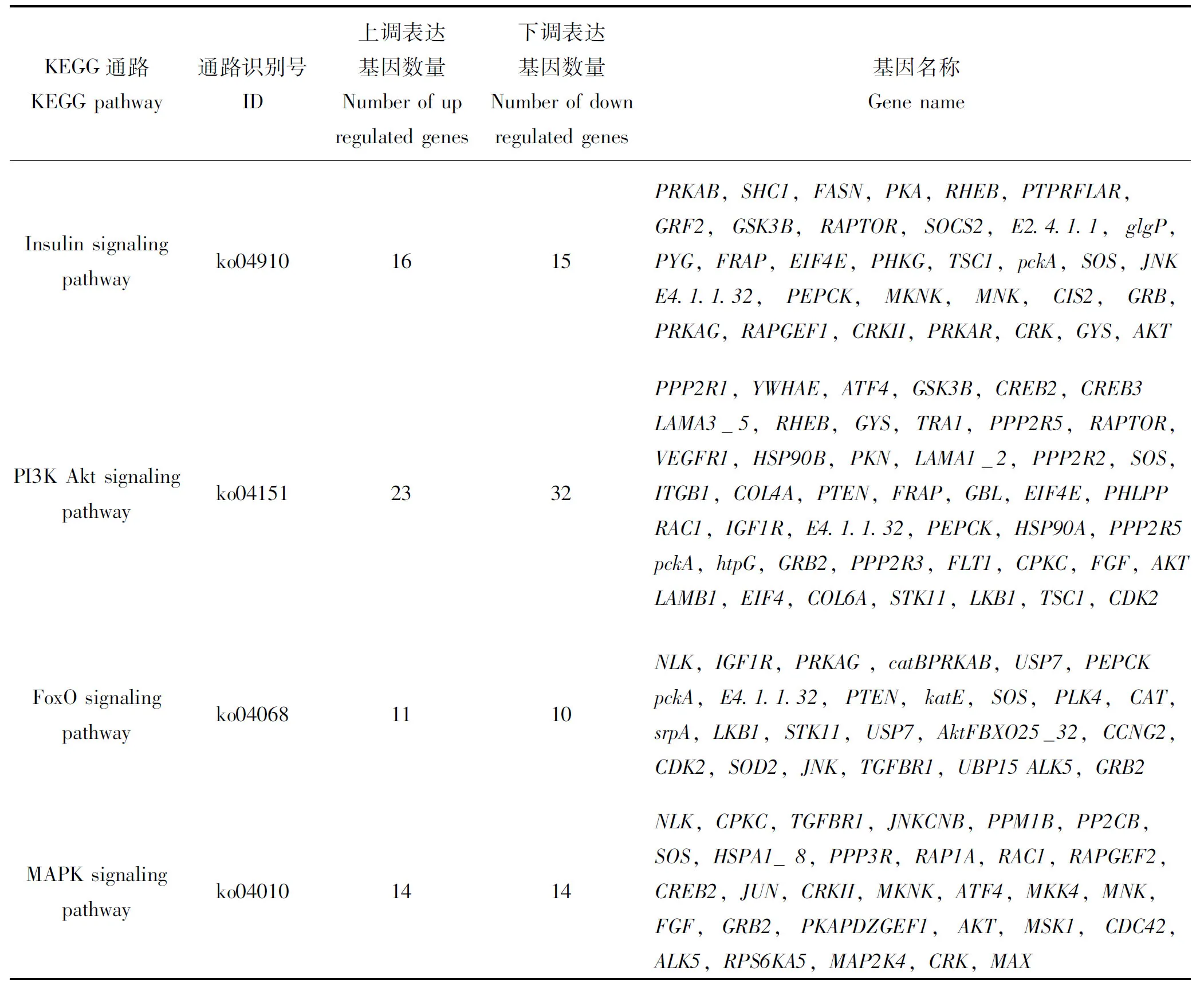
表1 胰岛素信号通路相关路径差异表达基因情况
3 结论与讨论
鉴于胰岛素样蛋白在昆虫生长发育、代谢、生殖以及衰老等生命活动中的重要性,本文对茶足柄瘤蚜茧蜂滞育组蛹与非滞育组蛹进行转录组测序研究,筛选出与胰岛素信号通路相关及相关途径的基因,并对其功能进行探索。研究结果为深入挖掘胰岛素信号通路及其相关途径有关基因的功能奠定基础。对果蝇(Tateretal., 2001;Williamsetal., 2006),库蚊Culexpipiens(Sim and Denlinger, 2008)和线虫Caenorhabditiselegans(Leeetal., 2001)进行研究发现,胰岛素信号可能是调控滞育的主要发育通路。胰岛素信号受抑制后,会导致这些生物体发育停滞。敲除果蝇编码胰岛素样蛋白、胰岛素受体和胰岛素受体底物的基因,或者过表达下游转录因子dFoxO,或者使用PIP3抑制剂PTEN,这些措施都能抑制胰岛素信号,最终导致寿命延长(Clancy, 2001)。敲除滞育昆虫的FoxO后,脂质积累立刻终止(Sim and Denlinger, 2008)。本试验研究发现,参与胰岛素信号通路,PI3K-Akt信号通路,FoxO信号通路,MAPK信号通路的重要基因,Sos、FASN、TSC1、JNK、PRKAB等基因在滞育的茶足柄瘤蚜茧蜂蛹中呈现不同程度的上调或下调表达。
Sos(Son of sevenless)基因最早在果蝇复眼神经发育中发现。该基因转录翻译成178 kDa的蛋白,在果蝇各个发育期均有表达。遗传学实验结果表明,表皮生长因子结合细胞生长因子受体激活结合蛋白GRB2,将Sos固定到膜上,随后Sos作为转化因子激活Ras绑定GDP形成GTP。从而开启下游的一系列级联蛋白磷酸化,最终激活MAPK信号通路。茶足柄瘤蚜茧蜂滞育蛹中Sos基因的下调表达,势必会导致MAPK信号通路受到抑制。ERK是MAPK家族成员,在豆长刺萤叶甲Atrachyamenetriesi应对低温胁迫和家蚕Bombyxmori调节胚胎滞育过程中起作用(Fujiwaraetal., 2006; Kidokoroetal., 2006)。研究人员发现,在家蚕滞育过程中,ERK通路调节类固醇和山梨醇的合成物,来终止家蚕幼虫的滞育(Fujiwaraetal., 2006)。Iwata发现ERK与家蚕滞育与再次发育有关(Iwataetal., 2005)。因此推测,在低温条件下,Sos对MAPK信号通路的影响主要是影响ERK活性。ERK通过参与昆虫在低温条件下的代谢,控制山梨醇、甘油等醇类物质的合成,来给出逆境保护措施,从而协助昆虫度过难关或逆境。茶足柄瘤蚜茧蜂滞育蛹中Sos基因对影响ERK在耐寒机制中的作用,还需要进一步的探究。
PRKAB属AMPK家族。AMPK指AMP激活的蛋白激酶(AMP-activated protein kinase),在真核生物中广泛存在,属丝氨酸/苏氨酸蛋白激酶。AMPK能感知能量代谢状态的改变,并通过影响细胞物质代谢的多个环节,来维持细胞能量供求平衡。滞育昆虫在能量来源紧缺的情况下,能够高效利用能量是非常重要的。昆虫通过在滞育准备阶段储存能源物质,滞育过程中降低代谢,来满足在滞育过程中的能量需求。积累充足的能源物质不仅可以帮助昆虫成功度过不良环境进入滞育阶段,还可以为滞育结束后的发育过程提供能量。营养物质利用在滞育阶段是一个变化的过程,昆虫能够根据自身能量的积累情况调节是否进入滞育以及滞育持续的时间(Hahn and Denlinger, 2011)。目前有实验结果发现,AMPK可调控Rac1(Leeetal., 2008)。Rac1, Ras相关的C3肉毒素底物1 (ras-related C3 botulinum toxin substrate 1),是Rho GTP酶超家族里Rac亚家族中的一员。Rho GTP酶可以在有活性GTP结合形式和无活性GDP结合形式之间循环,Rac蛋白也如此。正是这两种活性形式间的转换使得Rac1成为细胞内重要的信号转导分子。滞育的茶足柄瘤蚜茧蜂蛹中,Rac1基因下调表达,Rac1的激活受到抑制,因此细胞增殖受到抑制,这与茶足柄瘤蚜茧蜂在滞育期间形态不发生变化一致。所以本研究推测Rac1与茶足柄瘤蚜茧蜂滞育及再次发育相关,但具体怎么影响还需要进一步的实验验证。
PRKAB基因在茶足柄瘤蚜茧蜂滞育蛹中上调表达,说明AMPK与茶足柄瘤蚜茧蜂的滞育相关,本研究推测,AMPK主要影响滞育过程中茶足柄瘤蚜茧蜂的能量代谢。滞育过程中,AMPK可通过抑制脂肪酸氧化、葡萄糖转运等,减少ATP的产生,使代谢减缓;同时,通过促进糖原、脂肪、胆固醇的合成,保证有足够ATP以满足生命活动所需要的能量。胰岛素信号参与哺乳动物的糖代谢和脂类代谢的调控,因此推测胰岛素样蛋白也可能参与调控昆虫滞育过程中的能量积累。
在滞育的茶足柄瘤蚜茧蜂胰岛素信号通路中,脂肪酸合成酶(Fatty acid synthase, FASN)是催化脂肪酸合成的一种结合酶,FASN基因下调表达,说明在滞育过程中,脂肪积累减少。在对库蚊的研究中发现,在滞育准备阶段,库蚊增加糖类摄取,积累更多脂肪(Denlinger, 2005)。敲除FoxO后,库蚊雌虫不能像滞育过程中积累大量脂肪(Sim and Denlinger, 2008),将非滞育雌虫个体的胰岛素受体基因敲除后,卵巢发育受到抑制,促进滞育。干扰胰岛素信号,果蝇终止生殖发育并增加能源物质储存(Tatar, 2001)。由此可以认为,胰岛素信号在茶足柄瘤蚜茧蜂脂肪积累中起着关键作用。
本研究重点对胰岛素信号通路及相关途径中的一些基因与昆虫滞育的关系进行分析,明确胰岛素在昆虫中的作用。对天敌昆虫进行滞育诱导通常采用温光周期共同调控的传统方式,本研究找到了胰岛素信号通路中与滞育相关的基因,从基因水平对昆虫进行滞育诱导提供了新思路,同时对寻找胰岛素替代物施用于害虫进行生物防治也具有重要意义。但要明确,在昆虫中并不是单一信号通路对生物功能起作用,而是多种通路相互交错,共同发挥作用。信号通路对昆虫滞育滞育的影响,还需要进一步研究。
