干旱半干旱区斑块状植被格局形成模拟研究进展
2021-01-16刘庆生
刘庆生
中国科学院地理科学与资源研究所,资源与环境信息系统国家重点实验室, 北京 100101
斑块状植被格局是世界上干旱半干旱区常见的景观类型,美丽而神秘,一般形成于年降雨量50 mm到750 mm的区域,其发育的土壤可为砂质、粉砂、黏土,构成的植被可为林、灌、草,受区域降水、地质地形、土壤和植被的空间异质性及人类活动干扰的影响,常呈条带状、环状、斑点状等结构。单个斑块大小可从0.5 m2到1200 m2不等,具有生物多样性强、生物量高等特征,能减缓地表径流侵蚀、防风固沙及防止土壤养分流失等,因而受到广泛关注[1- 4]。斑块状植被格局综合反映了干旱半干旱区生态系统的生态水文过程,研究它们的形成、组成结构和演替过程,对于揭示干旱半干旱区生态系统变化的关键过程具有重要意义,且有助于制定适宜的干旱半干旱区退化生态系统修复策略,以及预测气候变化对干旱半干旱区生态系统的影响[5-14]。
斑块状植被格局研究始于20世纪40年代基于航空影像对非洲之角的调查研究,之后吸引了越来越多科学家的关注[14],提出了许多形成理论和假说[15],然而直到现在它们的起源和控制因素仍是未解之谜[16]。研究的热点包括不同尺度斑块状植被格局的精细识别、斑块状植被格局形成过程和机制的模拟,以及降水等自然因素和放牧等人为因素对斑块状植被格局的影响等。研究方法主要包括基于地面调查和遥感技术的定性、定量斑块状植被格局分析,以及基于个体、过程的植被格局形成数学模拟两种[17- 18]。前一种方法所涉及的研究区域和对象有限,尽管当今高分辨率卫星和低空无人机遥感技术能获得较大范围的斑块状植被格局数据,但对于斑块状植被格局研究所需的植被个体数量和生活史、土壤养分、水分等环境信息获取还能力有限,且获取的时间序列数据与植被格局演化时间相比相去甚远,因此利用数学模拟的方法来研究斑块状植被格局形成机制受到了越来越多的重视[19- 22]。已有文献较为详细地综述了基于地面调查和遥感技术的斑块状植被格局定性、定量分析[14,17,23- 24],讨论了斑块状植被格局形成机制及理论(基于对称破缺不稳定性理论的格局形成确定性机制,包括似Turing不稳机制、基于核的短距离合作与长距离抑制相互作用机制和差分流动不稳定性机制;噪声诱导格局形成的随机机制,包括非平衡相变机制和动力学随机切换诱导机制)[14,17,25- 26],论述了综合多种形成机理进行斑块状植被格局形成研究的必要性[15],本文不再赘述,将主要侧重于综述斑块状植被格局形成模拟研究进展,以期为推动干旱半干旱区斑块状植被格局形成机制研究提供科学参考和技术支撑。
当前植被格局模拟方法主要有两种,一为连续模拟,二为离散模拟[18,25,27]。前者利用的是求解偏微分方程的方法,并不针对植被或种群个体,而是用连续生物量面密度来刻画植被种群,更侧重于小尺度确定性的过程,所构建的多为时空连续模型。后者是一种随机计算模拟的方法,多侧重于斑块状植被格局中的植被个体或种群,所构建的多为时空离散模型[18,25,27]。相对来说,偏微分方程的方法是一种更适合于格局形成理论的强有力方法[18,25,27]。
1 斑块状植被格局形成的连续模拟
1.1 斑块状格局形成的反馈机制
植被群落构建机制一直是生态学研究的核心问题,前人已对其研究进展进行了系统评述[28-29]。尽管有研究表明社会昆虫生态系统工程师群体之间的区域干扰竞争可造成六边形巢址和相关植被的规则离散格局分布[15],但对于干旱半干旱区斑块状植被格局的形成来说,一般地认为是小尺度上植物和水反馈作用的结果[18,20,25,27,30]。为了揭示其形成机制,所构建的基于偏微分方程的模型必须能反映植物和水之间的反馈作用[25]。一般地,存在五种反馈作用[20,31]:一是渗透反馈,这是植物生物量与水之间的一种正向反馈,其增加了植被斑块区地表水渗入土壤的速率,加大了植被斑块区与周围裸土区的渗透率差异,进而促使地表水向植被斑块区流动,进一步促进植被生物量的增加;二是根扩展反馈,这是植物地上生物量与地下根之间的正向反馈,其增加了植物可利用水量,从而促使植物生物量的进一步增加;三是抽吸反馈,这是植物生物量与水之间的一种负向反馈,是由于植物根系抽吸水分而引起的。植物生长消耗水分,水分的消耗限制了植被生长;四是土壤水扩散反馈,是一种由于植物抽吸反馈引起的水分消耗促使植被斑块周边土壤水向植被斑块扩散移动的作用,其与植物的根茎比属性有关,间接反映了植物根系吸水能力;五是遮荫反馈,这是植物生物量与土壤水之间的一种正向反馈,其减少了植被斑块区的蒸发,促进了植物生长,但并未引起土壤水分的运移。五种反馈作用促进了植被斑块区植物生物量的增加,抑制了植被斑块区周围植物的生长,从而造就了干旱半干旱区斑块状植被格局。当然,放牧、气候变化、土壤异质性、降雨脉动、有毒化合物、动物和人类干扰等外部因素也能造就斑块状植被格局[14,32],特别是能改变斑块状植被的形态和格局,但其并非是必要因素[8,33-34],对这些因素的研究模拟,一般是在反馈机制模拟的基础上,增加特定项来进行。
1.2 斑块状格局形成的经典连续模型
最早的、现在依然经典的整合了格局形成理论的连续模型是Lefever和Lejeune[33]仅考虑了单一生物量动态变量构建的偏微分方程模型[14,25,31,35],其描述了植物之间交互作用引起的连续生物量动态变化,揭示了植被斑块群落内存在的近距离激励和远距离竞争的机制,模拟了干旱半干旱区常见的条带状(虎斑)格局的形成。该模型并没有清晰地分析土壤水分的动态变化,只是模糊地进行了环境假设。因此,难以用该模型研究环境对格局形成的影响[13]。其最基本的模型方程如下,详细内涵及进一步分解推导[14,25,31,33]。
(1)
式中,B(X,T)为T时刻X位置的单位面积生物量,FG反映植物繁殖和生长,FI代表植物之间的竞争和交互作用,FM反映植物的死亡或损失率。
随后,Klausmeier[36]构建了一个简单的包含两个动态变量(分别代表生物量和水)的连续模型,清晰地描述了植被和土壤水分的动态变化及其相互作用,模拟了均匀山坡上条带状植被格局的形成。与Lefever和Lejeune[33]的模型相比,该模型考虑了与生态水文过程相联的土壤水分动态变化,有助于理解地形、降水、放牧等对植被格局的影响。然而,该模型对土壤水分动态的模拟只是反映综合的跨时间尺度的水文过程,没有区分地表水和土壤水,这样其模拟结果与实际观测结果在许多方面并不一致。其基本的无量纲的模型方程如下,详细内涵及进一步推导[14,36]:
(2)
(3)
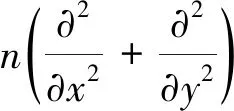
2001年,Von Hardenberg 等[38]构建了一个包含两个动态变量(生物量和水)的连续模型,其新的特征是引入了模拟由于植物根系抽吸水导致的植被斑块对于水分的竞争。该模型能模拟水分受限区域的多种植被格局形成,包括从低降雨条件下裸土到中等降雨条件下条带状、迷宫和斑点状植被格局,一直到高降雨条件下均匀植被。然而,其与Klausmeier[36]的模型一样,也未区分地表水和土壤水。其基本的模型方程如下,详细内涵及进一步推导[38]:
(4)
(5)
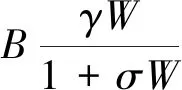
为了更好地反映土壤水动态相关的水文过程,HilleRisLambers 等[39],Rietkerk 等[40]提出了包含三个动态变量(生物量、地表水、土壤水)的连续模型,模拟植被扩散及其与周边地表水和土壤水的净交换。该模型适合于研究植被斑块区植被-水的交互作用,但也增加了模型计算的复杂性。这里仅给出Rietkerk 等[40]的模型方程,其他详细内涵及进一步推导[14,39- 40]:
(6)
(7)
(8)
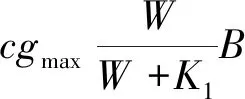
1.3 经典连续模型的改进和扩展
当前,斑块状植被格局形成的连续模拟逐渐向能更好地解释斑块状植被格局形成过程中起关键作用的多种生态水文过程的连续模型构建发展。Gilad[35]构建了更为复杂的包含三个动态变量(生物量、地表水、土壤水)的连续模型,能更好地反映水分受限系统中植被格局形成过程中影响植物生物量-水相互关系的渗透反馈、抽吸反馈和遮荫反馈机制。Saco 等[41]在Rietkerk 等[40]模型基础上,耦合了地表水路径,增加了地貌演化方程,解释了地表径流和沉积物分布引起的种子扩散,不仅模拟了植被格局,而且也模拟了区域径流和侵蚀格局。Zeng 等[42]研究了干旱半干旱草地生态系统植被格局的多样性和转换特征,构建了包含三个动态变量的连续模型,与前述模型不同的是,该模型的三个变量分别为植物鲜叶质量密度、土壤有效湿度、凋落和死叶的质量密度,模拟了随着湿度减少从均一草地到规则和不规则植被格局,最后到纯沙漠的转换,指出过渡区可能存在无穷多个平衡态。Liu 等[43]在Klausmeier[36]模型基础上,引入了一个反映由于植物根系抽吸导致的水资源再分布的正向反馈机制项到公式(3)中,模拟了植被格局形成,认为树比草对该正向反馈机制更敏感。Thompson 等[44]在Rietkerk 等[40]模型基础上,考虑了植物种子的变距离扩散,模拟结果表明种子扩散行为能改变植被的空间结构,修订后模型的模拟结果与实际观测更吻合。Barbier 等[45]构建了单一植被生物量动态变量的积分微分方程,成功模拟了尼日尔西南部空斑植被格局的形成,认为植物之间相互促进和竞争可以诱导景观尺度植被格局,并形成环境和植被之间的双向反馈循环。Van der Stelt 等[46]通过增加地表水的可能非线性扩散项,修订了Klausmeier[36]模型,使之也可用于平坦地形区植被格局形成的模拟。Escaff 等[47]考虑了强非局部耦合和均匀植被态与裸土态之间的双稳态特性,构建了单一植被生物量动态变量的变程非局部纳古莫型方程(Nagumo-type equation),模拟了仙女环和斑点状植被格局的形成,结果表明仙女环的宽度随着干旱程度的增强而强烈增加,相反地,斑点状植被斑块的宽度随着干旱程度的增加而减小。Bordeu 等[2]提出一种利用具有非局部项的逻辑斯蒂(logistic)方程来描述归一化生物量的时空演化的特殊方法,其考虑到了植物间的激励和竞争及种子扩散反馈机制,模拟结果表明干旱程度减少引起的植物自我复制和竞争性相互作用引起的植物分布共同促成了干旱区生态系统植被的空间组织格局。Kinst 等[48]提出了一个耦合植被-结皮的模型,包含了植被生物量、结皮生物量、土壤水和地表水四个动态变量,模拟了沿降雨梯度,不同植被-结皮状态下干旱区斑块状植被从空间均匀到周期性景观格局的条件,认为结皮动态的加入使得植被格局的出现需更高的降水量,同时增加了生物量的变化幅度。Tlidi 等[49]给出了Bordeu 等[2]模型的积分微分方程,考虑了非局域植物间激励和竞争相互作用,模拟了各向同性环境条件下平地上单一植物种格局的形成,结果很好地解释了赞比亚和莫桑比克斑点状植被格局源于植被的自我复制机制。Wang 等[32]构造了包含两个动态变量(生物量和水)的连续模型,考虑了扩散和交叉扩散来模拟植被和水之间的相互作用。Tlidi 等[50]考虑了资源积累和周边植被引起的蒸散减少产生的植物生长、种子繁殖和扩散等造成的生物量增加、资源竞争造成的生物量减少和由于干旱增加造成的植物生存比降低等,构建了单一植被生物量的通用交互再分配方程,模拟了干旱景观中各向同性环境条件下螺旋形和弧状植被格局的形成。Eigentler 等[51]构建了一个非局域的Klausmeier[36]模型,其增加了一个描述植被扩散的非局域卷积积分项,解释了植物种子长程扩散的可能性,更好地模拟了半干旱区条带状植被格局的形成。Huang 等[52]考虑了土壤层对植被格局形成的影响,对Klausmeier[36]模型进行修订,模拟了沉积层中水资源对植物格局的改变,模拟结果为严重侵蚀区自然植被恢复提供了理论支撑。
1.4 连续模型的优势与局限性
斑块状植被格局的连续模拟依据自然界的格局形成理论,基于植被斑块中植物之间激励与竞争机制、植物与水之间的相互作用机制来构建模型,模型多用偏微分方程、常微分方程、积分微分方程等数学函数来表述。现在大多数的连续模型可以或者至少部分用计算机数值技术来解决,适合于解析描述,一旦确定了描述模型各种状态的解,就可以更好地理解各种过程及参数对植被格局形成的影响以及对外部环境的响应机制[17,25]。依据考虑变量的多少和对关键过程描述的详细程度,模型各有不同。一般地,所构建的连续模型越详细,对斑块状植被格局的形成解释地越好,同时复杂性也增加,需要更多的数学分析和数值模拟工作。空间上考虑一维的研究较多,二维的较少;考虑一种植被类型的较多,两种以上的较少,很少考虑植物个体的生命周期及行为对格局的影响,往往把植被看作一个整体,用植被生物量来代替[53];从均匀植被退化到裸土的正向模拟较多,而从退化土地恢复植被的反向模拟较少[52]。纵观文献,目前连续模型仍然是斑块状植被格局形成模拟的主体,相关研究层出不穷。值得注意的是,已有的许多连续模型均能模拟出合理的植被格局,模型简化太注重普遍性,而可能忽视了植被格局形成的真正原因,而这正是应对气候变化和人类活动的影响所需要的。模型模拟结果大多是与其他模型模拟结果相比,与实际观测的植被格局比较和验证较少[16],今后应注重这方面的研究。
2 斑块状植被格局形成的离散模拟
2.1 离散模拟的主要方法
与连续模型相比,斑块状植被格局形成的离散模拟出现地更早。第一批植被格局形成的离散模拟出现在20世纪90年代初期到中期[14,17]。Sato和Iwasa[54]构建了以一排树的高度为单一变量的离散集群模型,从树的随机分布开始,模拟亚高山冷杉锯齿状空间格局的形成,结果表明该格局以不变的形态按固定速率向下风向移动,与一维模型相比,二维模型趋向于产生更为规则的波浪再生格局。Mauchamp 等[55]基于斑块内动态、交错带动态,以及斑块与资源流之间的交互作用等规则构建了一个简单的时空离散元胞自动机模型,通过对每个元胞的水和碳收支功能、植物的补充和生存过程的模拟,实现了干旱区双斑块系统(植被镶嵌格局)的格局动态与功能模拟。Thiery 等[56]考虑了单元的四种状态(无树或死树、幼树或衰老的树、小树或受胁迫的成年树、生长好的成年树),以及临近位置树的状态对当前位置的树的影响,构建了尼日尔西南部树的不规则条带状格局形成的时空离散元胞自动机模型,结果表明模拟结果与初始树的密度无关,树的结构和格局变化仅与反映上坡向资源竞争、横向协同作用的参数和模型迭代次数有关。Wiegand 等[57]基于五种主要植物详细的生活史数据,构建了南非干燥台地高原灌丛群落时空动态的基于个体的离散模型,揭示了降水对灌丛组成及格局的影响。
研究所采用的集群模型、元胞自动机模型和基于个体的模型是斑块状植被格局形成离散模拟的三种主要方法,它们的共同点是将空间和时间划分为一定数量的离散单元,每个单元代表植被个体或植被群,每个单元被赋予一定的模型过程。集群模型基于非常简化的框架来模拟复杂系统的动态,其每个单元代表植被群,每个单元有其自身的动态变化,与其它单元的动态变化无关,单元间植被短距离合作-长距离竞争的相互作用导致植被格局的形成。元胞自动机模型基于一组表征系统动态过程的规则来模拟系统动态,植被种群的行为表征为每个单元的状态特征,每个单元的状态仅与上一时刻该单元及其相邻单元的状态有关,上一时刻每个单元状态变量根据确定的规则发生改变,每个单元一个时刻只能拥有一个特定状态的,模型中的单元根据局部规则同步更新,每个单元与其相邻单元之间的相互作用强弱导致了不同植被格局的形成。基于个体的模型中每个单元代表成熟植物个体的平均尺寸或最大尺寸,对于仅包含植物个体的单元考虑个体的行为,对于其它单元考虑单元的内部功能,单元的状态及其变化对相邻单元的响应用一组规则来表示,每个单元中植物个体的行为以及每个单元与其相邻单元植物个体之间的相互作用导致植被格局的形成。离散模型假设离散单元内的空间变化和过程可以忽略,注重离散单元间的空间变化和动态过程,这样大大简化了模型,特别是对离散单元间相互作用关系的模拟和计算上,由此也具有计算模拟上的优势[19]。离散模拟大多是空间显式的,这也促进了随后其在斑块状植被格局形成模拟中的广泛应用。
2.2 基于植被-土壤水内在反馈机制的离散模拟
Dunkerley[58]构建了一个简单的离散元胞自动机模型,每个单元依据水的分配和植物生存规则来确定其植被覆盖率和土壤湿度,通过多次迭代模拟了一致降雨条件下条带状植被格局的形成。Keymer 等[59]构建了一个简单的离散集群模型,每个单元的状态(植被、非植被)由该单元及其相邻单元的状态的局部转换规则来确定,结果表明:尽管环境在空间上是均匀的,但由于局部的相互作用和特定的定殖和灭绝参数,自组织时空模式呈现出分形聚类。Rietkerk 等[5]基于修订的Thiery 等[56]简单元胞自动机模型,论述了利用尺度依赖反馈机制如何解释生态系统中的格局多样性,结果表明不同强度和尺度的反馈作用产生了不同结构和尺度的自组织斑块状空间格局。Feagin 等[60]构建了美国德克萨斯州加尔维斯顿岛沙丘植被群落格局形成的元胞自动机模型,每个单元的状态(裸地、定殖植物、土壤粘合剂植物、竞争性植物)由相邻植物的促进作用和先前演替阶段对环境约束的改善确定的十个规则来决定,结果表明植物个体尺度和群落尺度的过程共同决定了沙丘植被群落格局。Shoshany 等[61]利用元胞自动机模型和概率转换规则模拟了裸坡上植被的恢复,认为土壤和植被格局特征(斑块数、密度、破碎度、优势度和形状分布参数)的协同变化是植被格局形成的关键过程。Scanlon 等[62]考虑了相邻树密度的局部作用和总体植被覆盖率的全局作用对于单元两种状态(树冠、非树冠)间可能转换的影响,构建了元胞自动机模型,发现观测到的树冠丛大小的幂律分布可能是由于全球尺度的资源约束(即水的有效性)和局部尺度的激励机制相互作用的结果。Cipriotti 等[21,63]构建了一个空间显式的基于个体的模型,模拟了三种优势草和优势灌木的时空动态,考虑了植被地上生物量、六个0.1 m厚的土壤层剖面(包含各层的植被根和土壤水),以及土壤种子库。对于每个植被单元,模拟植被个体的种子生成、种子扩散、植物生长和死亡过程,对于非植被单元考虑出苗和补充过程,对于所有单元模拟土壤种子库的动态,结果表明植物的出苗和补充是半干旱区草-灌草原长期植被格局动态的关键过程,内部水分动态导致灌木与邻近草的竞争减少足以产生灌木被草环绕形成的环形斑块结构,灌-草共生机制对干旱草原格局至关重要,火和草食并不是它们形成的关键驱动力。
2.3 降水、放牧等外部环境对植被格局形成的影响离散模拟
Dunkerley[64]构建了一个简单的离散元胞自动机模型,包含了干旱对土壤含水量的影响和放牧压力对植被生物量的影响规则,结果表明:除非极度放牧,否则放牧对条带状植被格局的生存影响很小,干旱与极端干旱位置处植物的系统损失会导致大范围条带状植被格局的破碎化。Puigdefabregas 等[65]构建了一个离散元胞自动机模型,模拟了风和树生长之间的相互作用,认为条带状植被格局是由树的生长率和风致死率共同决定的,树木生长速度的增加会导致条带状植被格局的波长变长和波的传播速度变快,相反,风致死率的增加会导致条带状植被格局的波长变短和波的传播速度变慢。Yokozawa 等[66]利用耦合映像格子模型(时空离散,状态变量连续)模拟了单个树木之间的竞争模式和外部干扰强度对亚高山林分波浪再生空间格局动态的影响,结果表明:与不对称竞争模式相比,波浪状空间格局更容易在对称竞争模式下出现。Adler和Raff[67]构建了一个离散元胞自动机模型探究放牧如何持续地抑制竞争植物物种的精细尺度空间格局,结果表明:与放牧影响或其他环境异质性性比,植物个体尺度相邻单元之间的相互作用是导致斑块状植被格局的主要原因。Peters[68]为了探讨草地-灌丛过渡区植物生活史性状、优势格局和植被组成之间的关系,以预测随着气候变化优势格局的转变,构建了一个混合植物生活史的基于个体的空斑动态离散模型,模拟了多个优势种之间在多尺度空间的相互作用,结果表明:在年温度和夏季降雨增加的气候情景下,植被格局将向单一草地控制的格局转变,土壤水的时空分配对于草-灌之间的相互作用至关重要,基于个体的模型能够很好地模拟草本-灌木植物种之间复杂的相互作用。Esteban和Faireén[69]构建了离散元胞自动机模拟半干旱区异质性缓坡条带状植被格局的自组织形成,模拟结果表明,在降雨较少的条件下,植被条带会变窄,但其生产力高,在降雨增加的条件下,植被条带会变宽,但其生产力会降低。与前人条带状植被格局的研究结果一致,总体上看,条带状植被格局会向上坡迁移。皱梅[70]构建了黄土高原半干旱丘陵区羊道斑状植被格局形成的元胞自动机模型,假设羊道路径的形成主要受微地形坡度、多年生植物、土壤紧实度、位置偏离、采食搜索范围等因素影响,模拟了羊道景观格局的形成,结果发现羊的采食宽度小会形成密集的羊道格局,采食宽度大则形成稀疏的羊道格局。Franz 等[71]基于三类典型的裸土、草和树斑块的水平衡,构建了空间显式的元胞自动机模型,通过改变激励-竞争的范围,模型展示了一系列最有效的山坡水利用模式(从随机到高度组织的静态植被模式)。King和Franz[72]在Franz 等[71]模型基础上耦合了一个基于元胞自动机构建的动态植被模型,基于随机降雨,模拟了旱季-生长季循环过程中每日土壤湿度,进而模拟了不同放牧强度下植被格局动态。结果表明:在生长季结束时,依据生长季平均土壤有效湿度,在旱季末,依据放牧强度和前期生长季土壤湿度,斑块植被类型可发生两次转换。
2.4 离散模拟的优势和局限性
离散模型没有明确的数学理论,大多基于制定的规则来进行数值模拟[18],因此对于植被格局的野外观测要求较高,需要从中获得大量数据支撑,并由此制定出单元状态转换的规则。离散的时间和空间,计算相对简单,可操作性强,有利于提高计算机模拟的效率,便于模拟结果的空间显式表达,可同时模拟多种植被,每个单元可以有多个状态特征,因其是以规则的形式来模拟过程而不是数学公式,对于某些复杂的植被格局过程来说,可直接利用专家知识,而不受到所需数据的限制,在斑块状植被格局形成模拟中具有一定的优势[73]。斑块状植被格局的离散模拟以元胞自动机模型为主,集群模型、耦合映像格子模型和基于个体的模型相对较少,但随着对植物个体尺度相互作用机制在植被格局形成中的作用的重视,以及基于个体的模型建模方法等方面的逐渐提高,基于个体模型模拟斑块状植被格局的形成研究将逐渐增多。总体上看,与之前Tietjen和Jestsch[74]对1995—2005年半干旱区放牧系统和气候变化模拟模型的研究综述结果相似,即呈现离散模拟下降-连续模拟研究增长的态势。
3 结语与展望
3.1 结语
斑块状植被格局综合反映了干旱半干旱区生态系统的生态水文过程,它们的形成、组成结构和演替机理研究,对于揭示干旱半干旱区生态系统变化的关键过程具有重要意义。从20世纪90年代初期发展至今,斑块状植被格局形成的模拟研究层出不穷,然而,我们必须清醒地认识到,斑块状植被格局的形成理论永远不可能简化为简单的数学方程,也不可能获得离散模型所需的植被格局的全部参数信息,而更可能是一组过程机制的简化规则和知识;不要试着将所有过程整合到一个模型中,不仅是因为各种过程依据的理论不同,而且其研究的尺度等也很难统一[53],至今,还没有一个模型能完美地解决斑块状植被格局形成模拟中的所有问题,斑块状植被格局形成的模拟研究仍将是一个长期而艰巨的任务。当前,无论是连续模拟还是离散模拟,模型共享有待加强,一方面可以吸引更多的斑块状植被格局形成模拟的研究者加入,另一方面可促进已有模型在更多区域的应用和验证,由此进一步推进斑块状植被格局形成模拟的深入研究。
3.2 展望
综合已有研究成果,将来应着重注意和加强以下几个方面的研究:
(1)斑块状植被格局的连续模拟今后应加强与实际观测的植被格局比较和验证[16]。无论是连续模型还是离散模型,应将降水等气候条件、土壤异质性等外部因素加入到模型中,使得模型能更好地刻画和揭示植被格局动态,应进一步把植被格局和功能联系起来,以应对气候变化对干旱半干旱区生态系统的影响[75-78]。
(2)研究连续模型和离散模型之间的关系,考虑连续模型的离散化。相关研究表明,离散化的经典Klausmeier模型在定量描述半干旱区多样性植被格局方面更有优势,能更好地预测降雨和植被类型变化引发的植被格局转换,离散后模型的非线性机制能更好地揭示半干旱区植被格局形成的复杂性[79]。
(3)综合连续和离散模型各自的优点,构建混合模型。连续模型能成功模拟植物新陈代谢和土壤水文等连续过程,离散模型对植物个体异质性和离散时空相互作用概念能很好地模拟。结合两者的优点,混合模型能更好地模拟斑块状植被格局的形成。目前已出现耦合系统动力学与基于个体模型的混合模型研究植被自组织空间结构对气候胁迫的响应[53],以及整合Logistic模型和元胞自动机进行群落动态与规则植被格局之间的相互关系的研究[80],是今后值得重视的研究方向之一。
(4)应采用面向格局的模拟方法建模。面向格局模拟聚焦复杂系统的内部结构和过程的最基本信息,避免模拟模型结构和机制过于简单或过于复杂而增加的不确定性。利用植被格局去测试和对比植被个体或群落的行为或小尺度过程是对现实植被格局系统如何形成以及它们对外部因素影响如何响应研究更为聪明的办法[81-82]。
(5)应构建空间显式斑块状植被格局形成模拟模型。尽管非空间显式模型仍然是连续模拟的主流,但空间显式模型能解决特殊情况下植物种群或群落过程由于空间异质性、植被个体行为和其他偶然因素产生的无法融入非空间显式模型的情形[83]。
(6)应注重引入新的方法对模型中的状态和参数进行更为精确地预测。深度神经网络已被成功用于预测植被状态的转换概率以及对变化环境条件的响应[84]。
(7)应注重研发斑块状植被格局形成过程中的标准子模型及模型开发和集成平台。将来的离散模拟应采用标准开发协议基于已有模型来研发,以此促进模型的交流、共享、应用和验证。基于个体的模型在该方面已提供了很好的示例[19,85],值得借鉴。
