宝天曼中小型土壤动物功能类群及其影响因素研究
2021-01-15徐帅博李艳红冯小燕朱连奇
徐帅博,李艳红,冯小燕,李 理,朱连奇
(河南大学 环境与规划学院,河南 开封 475004)
土壤动物作为陆地生态系统功能发挥的调节者,在分解生物残体、转化矿物质和有机质方面起重要作用,是土壤中物质循环和能量流动的主要驱动者[1-2].土壤动物种类丰富,数量繁多,不同类型土壤动物虽然在体形、活动能力及方式等方面各不相同,但许多类群在功能和食性上相近,占据着相似的生态位,在生态系统中具有相似的生态作用[3-6].土壤动物主要通过各个功能类群间的食物网关系发挥其生态作用[7],单一类群很难完成这一作用过程.按照食性、取食方式、生活史策略等差异将土壤动物划分成不同的功能类群,简化了复杂的生物群落研究环节,便于系统性地认识和理解不同功能类群的生态作用过程[8-9].
目前,国内外学者围绕土壤动物功能类群已经开展了大量研究工作.国外学者Root针对同功能类群物种首次提出了“种团”(Guild)一词,意为“以类似的方式利用同一类环境资源”的物种集团.随后又增加“摄食方式”(Mode of Feeding)作为第二个判别标准,在对土壤微型节肢动物的研究中将不同的繁殖方式与代表这些方式的物种所对应环境条件联系起来,总结出12种“生活史策略”(life-history tactics);Brussaard进一步将物种功能群分类标准总结为生活史策略、微生境、主要食物、摄食方式和生理生态学[10-13].近年来欧洲学者使用稳定同位素、脂肪酸标记等在土壤动物功能群研究方面也取得了较大进展[14-16].国内学者借鉴国外的理论,对我国土壤动物功能类群进行了研究.相关学者研究松嫩草原大型土壤动物功能类群,指出大型土壤动物生物量是生态系统功能研究的重要依据[6,17];还有一些学者从林分结构、经营措施等方面探讨其对次生林土壤动物功能群的影响[18-20];王仙攀等研究认为,喀斯特地区潜在和轻度石漠化生境不利于捕食性土壤动物的生存、繁殖活动[21].总体而言,当前国内对土壤动物功能类群的研究主要集中在东北和西南地区,而对南北过渡带土壤动物功能类群的报道还相对缺乏.
宝天曼国家级自然保护区地处北亚热带向暖温带的过渡区,也是我国第二、三阶梯的过渡区,是对气候变化较为敏感的区域[22-24].目前,该区域物种功能类群研究多集中于植物群落方面,针对土壤动物的报道并不多见[25-27].为此,本文对宝天曼自然保护区南坡中小型土壤动物功能类群开展调查研究,旨在揭示保护区不同海拔梯度土壤动物功能类群特征,为保护区森林资源的保护和物种多样性的维持提供理论依据,同时为南北过渡带森林生态系统的调控提供理论参考.
1 材料与方法
1.1 研究区概况
宝天曼国家级自然保护区地处河南省南阳市内乡县北部(33°20′12″~33°35′43″N,111°46′55″~112°03′32″E).该区位于北亚热带向暖温带的过渡地区,属于季风型大陆性气候,是河南省生物多样性的分布中心,年均温15.1 ℃,年均降水量885.6 mm,相对湿度68 %[23,28];平均海拔800~1800 m;土壤分为棕壤、黄棕壤和褐土3个类型,土壤pH值约为6.5;植被类型主要为落叶阔叶林和针阔混交林,是我国中部地区保存最为完好的天然阔叶林区[29].
1.2 样品采集与分离
2018年9月份在宝天曼自然保护区南坡沿海拔梯度设置样带进行取样(表1).考虑到低海拔地区人类干扰较强,样带选择在海拔900~1700 m之间,沿坡面每间隔100 m左右设置1个20 m×20 m的样地,每个样地采用对角线法选择5个样方作为重复样,按O层(凋落物层)、A层(0~5 cm)、B层(5~10 cm)、C层(10~15 cm)分4层进行取样,共取160份土壤动物样品,编号后装入自封袋(敞口)带回实验室.同时,在样地内按“蛇形布点法”采集混合土壤样品约500 g,用于土壤理化性质测定.采用Tullgren法分离中小型土壤动物,并用95 %浓度酒精固定.使用LeicaM125双目体视显微镜,参照《中国土壤动物检索图鉴》对土壤动物进行分类鉴定[30].中小型土壤动物一般鉴定到亚目和科级,同时记录个体数量.因为土壤动物幼虫和成虫生态功能可能不同,因此将幼虫和成虫分别统计[31].
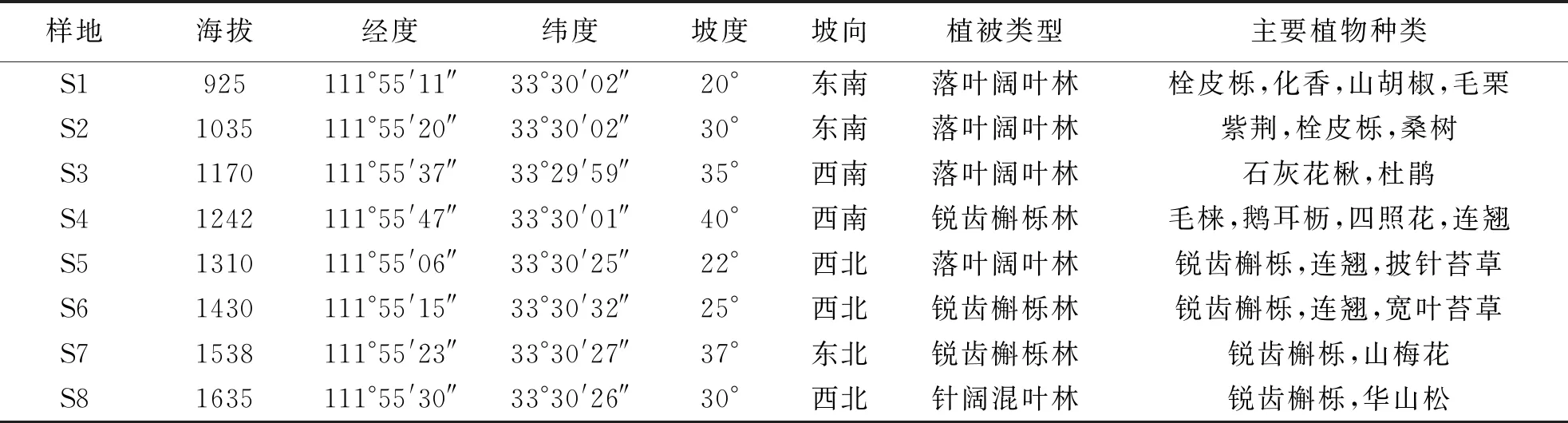
表1 研究样地基本概况
1.3 土壤理化因子测定
土壤理化指标参照鲁如坤《土壤农业化学分析方法》进行测定[32].其中土壤容重采用环刀法测定;土壤含水率采用烘干法测定;pH值使用水浸提法测定;有机质含量采用重铬酸钾—硫酸溶液氧化法测定;土壤全氮采用凯氏蒸馏法测定;土壤全磷采用钼锑抗比色法测定.
1.4 数据处理
根据土壤动物的生活型和食性差异,将宝天曼中小型土壤动物划分为腐食性(Saprophygous,Sa)、植食性(Phytophagous,Ph)、捕食性(Predatory,Pr)、杂食性(Omnivorous,Om)4个功能类群[9,33-34].采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(E)、Simpson优势度指数(C)、Margalef丰富度指数(M)分析中小型土壤动物各功能类群多样性特征,各指数计算公式如下:
(1)
E=H′/lnS,
(2)
(3)
M=(S-1)/lnN.
(4)
上式中,Pi=ni/N,ni为第i个类群的个体数,N为群落中所有类群的个体总数,S为总类群数.
采用Sorensen相似性指数分析各样地中小型土壤动物功能类群相似性特征.
Cs=2c/(a+b).
(5)
式中,a、b分别为群落的类群数,c为两群落共有的类群数.
相对多度:个体数量占群落总个体数10.00% 以上为优势类群;占1.00 %~10.00 %为常见类群;占比小于1.00 %为稀有类群.
分析中小型土壤动物数据之前,先进行log(x+1)对数转换.对服从正态分布的数据使用单因素方差分析(One-way ANOVA)比较不同海拔、土层中小型土壤动物功能类群个体、类群数之间的差异,并用最小显著差异法 ( LSD )比较均值;运用排序软件CANOCO 5.0对土壤动物各功能类群与土壤理化因子先进行DCA分析,由于第一、二排序轴长度均小于3,因此选择冗余分析 (Redundancy analysis, RDA).所有数据使用Microsoft Excel 2010、IBM SPSS Statistics 22.0和CANOCO 5.0进行分析处理,最后使用OriginPro 9.1软件作图.
2 结果与分析
2.1 中小型土壤动物功能类群组成
宝天曼自然保护区共捕获中小型土壤动物48类、3673只(表2).其中腐食性、植食性、捕食性、杂食性4种功能类群土壤动物个体数分别为2413、186、82、992只,分别占个体总数的65.70%、5.06%、2.23%、27.01%;类群数分别为16、11、8、13个,分别占类群总数的33.33%、22.92%、16.67%、27.08%.研究区中小型土壤动物功能类群个体数、类群数大小依次为:腐食性>杂食性>植食性>捕食性.
2.2 中小型土壤动物功能类群结构
2.2.1 不同海拔中小型土壤动物功能类群结构
研究区8个样地中小型土壤动物不同功能类群个体、类群总数如图1所示.其中腐食性和杂食性个体、类群总数均随海拔升高呈现先增后减的“单峰型”分布趋势,植食性和捕食性随海拔的上升其个体和类群总数无明显变化规律.方差分析结果显示,海拔对腐食性和杂食性土壤动物个体总数影响显著(P<0.05),对腐食性、植食性和捕食性土壤动物类群总数影响显著(P<0.05).
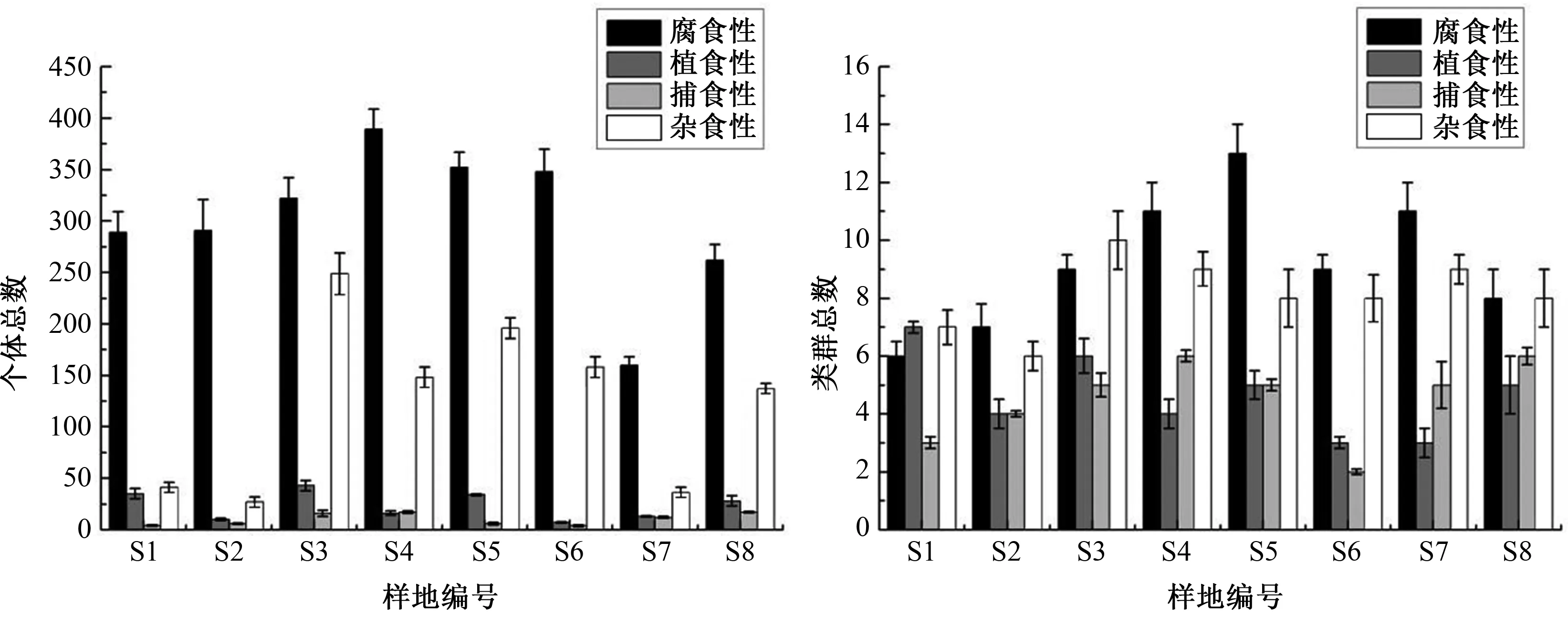
图1 宝天曼不同海拔中小型土壤动物功能类群个体、类群结构(平均值±标准误)
2.2.2 不同土层中小型土壤动物功能类群结构
各样地不同土层深度对4种功能类群土壤动物个体、类群分布影响不同(图2).除杂食性土壤动物外,各样地均为O层和A层个体总数显著大于B层和C层(P<0.01),且O层个体总数最多;腐食性和杂食性类群总数O层最大,A、B、C层无显著差异(P>0.05),植食性和捕食性各层差异显著(P<0.05).各样地4种功能类群均表现出明显的“表聚性”分布特征.
2.3 中小型土壤动物功能类群多样性特征
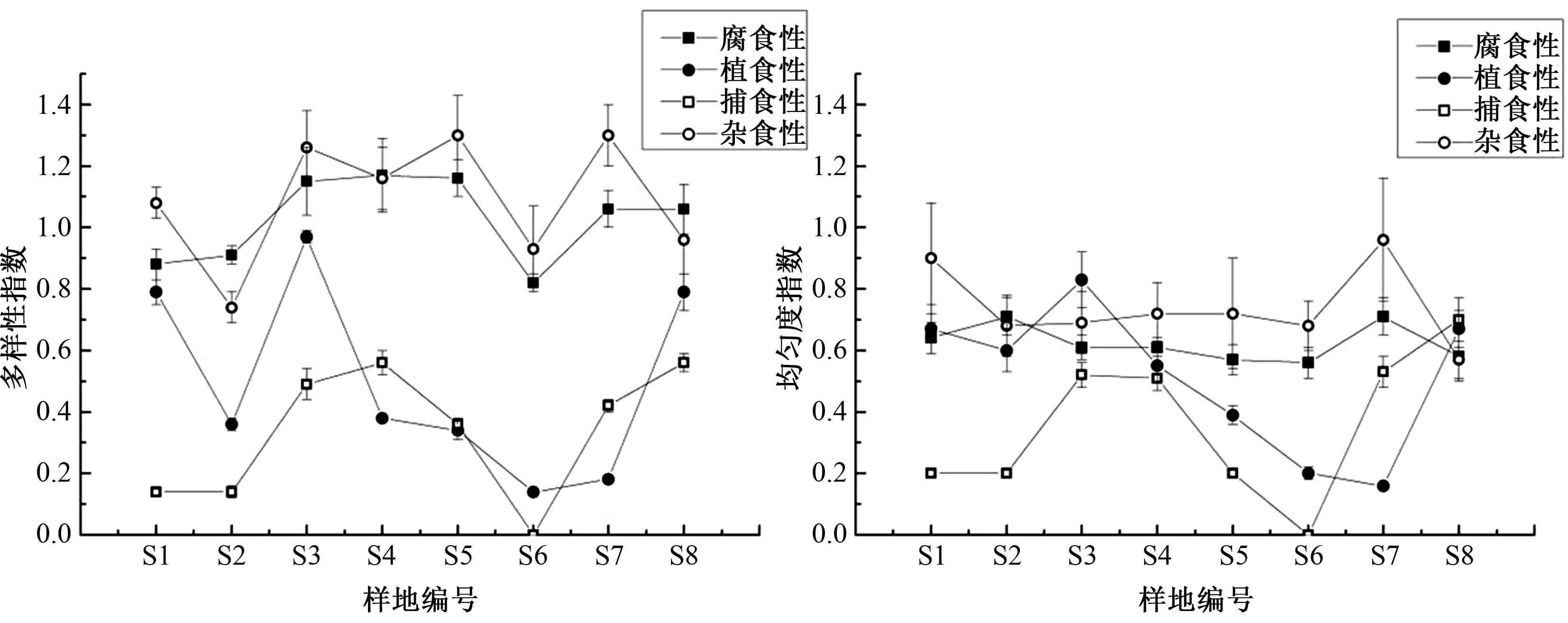
如图3所示,杂食性和腐食性土壤动物多样性指数最高,植食性土壤动物优势度指数最高,腐食性土壤动物丰富度指数最高;捕食性土壤动物多样性和均匀度指数最低,腐食性土壤动物优势度指数最低.单因素方差分析表明,海拔变化对4种功能类群多样性指数影响显著(P<0.05),对捕食性土壤动物均匀度指数影响显著(P<0.05),对植食性土壤动物优势度指数影响显著(P<0.05),对腐食性土壤动物丰富度指数影响极显著(P<0.01).
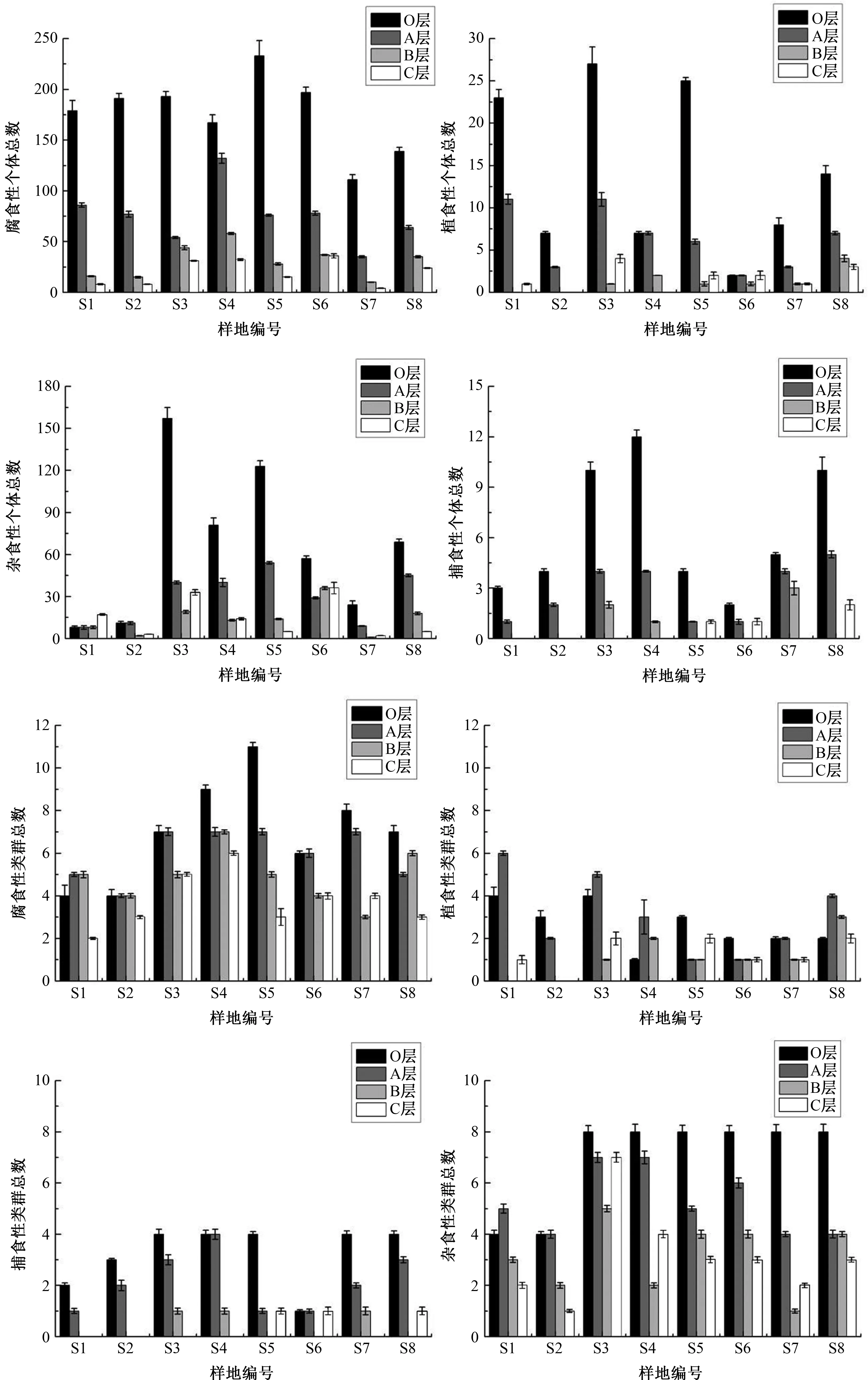
图2 宝天曼不同土层中小型土壤动物各功能类群个体、类群结构(平均值±标准误)
2.4 中小型土壤动物功能类群相似性特征
研究区中小型土壤动物4种功能类群相似性指数存在差异(表3).腐食性土壤动物Sorensen相似性系数为0.63~0.92,杂食性土壤动物Sorensen相似性系数为0.57~0.94,各样地相似性水平在中等相似以上,说明各样地腐食性、杂食性土壤动物组成和结构相近,共有类群较多;植食性土壤动物Sorensen相似性系数为0.29~0.89,捕食性土壤动物Sorensen相似性系数为0.00~0.91,表明这两个功能类群在各样地中具有较多的特有类群.总体来看,海拔是影响研究区植食性、捕食性土壤动物分布的重要限制性因素,这两个功能类群对生境改变具有较强的敏感性.
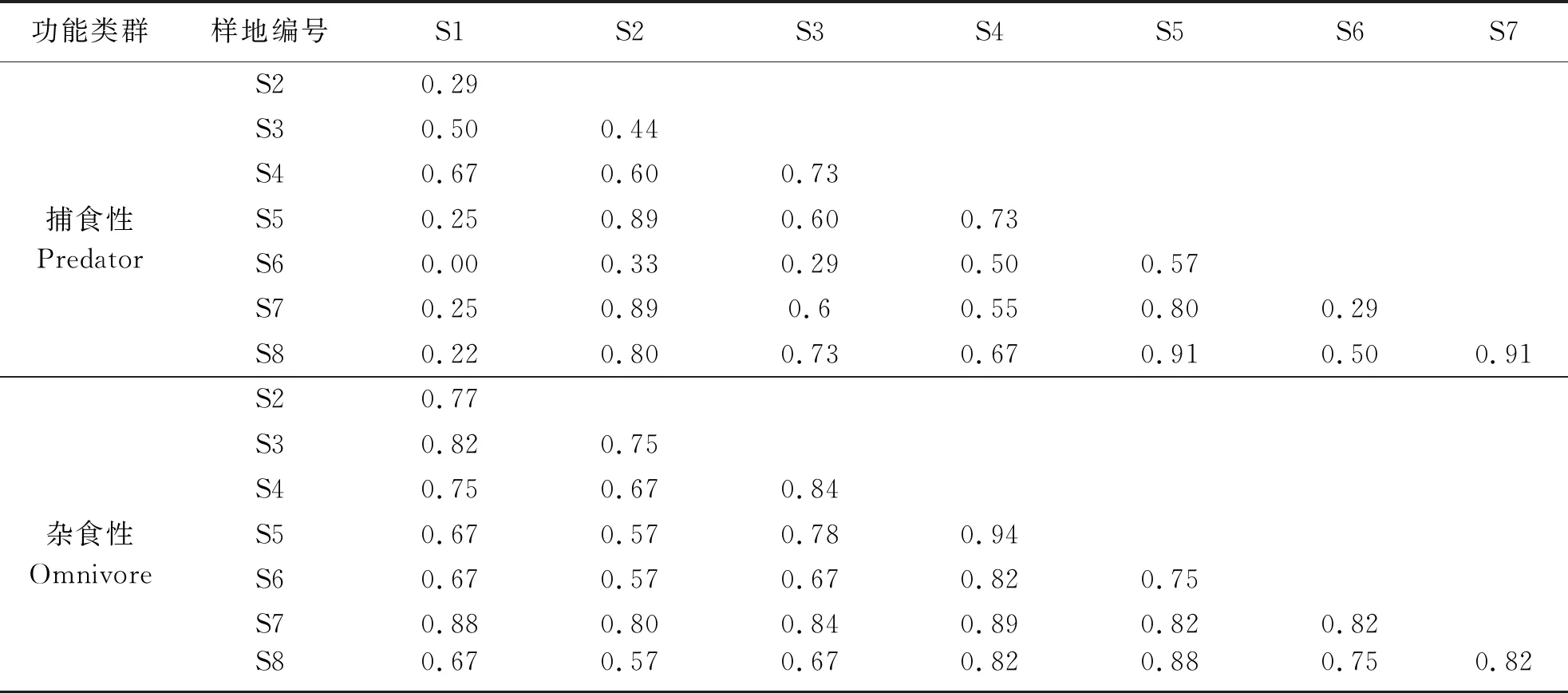
续表
2.5 各功能类群与土壤环境因子的RDA分析
选择土壤容重、含水率、pH值、有机质、全氮、全磷6种土壤理化指标作为土壤环境因素与研究区中小型土壤动物4个功能类群进行排序分析.在排序之前,先对各功能类群进行DCA分析,分析结果中最大轴长度小于3,因此可以使用RDA分析.经蒙特卡洛检验各排序轴均达到显著水平(P<0.05),说明RDA排序结果可信.RDA分析(图4)表明,第一排序轴与土壤含水率呈正相关关系,与土壤容重负相关;第二排序轴与全氮呈正相关.其中,腐食性和捕食性功能类群与土壤含水率呈显著正相关,与容重无明显相关性;杂食性土壤动物与有机质和全磷正相关;植食性土壤动物功能类群受全氮含量影响较明显,与pH值相关性不大.
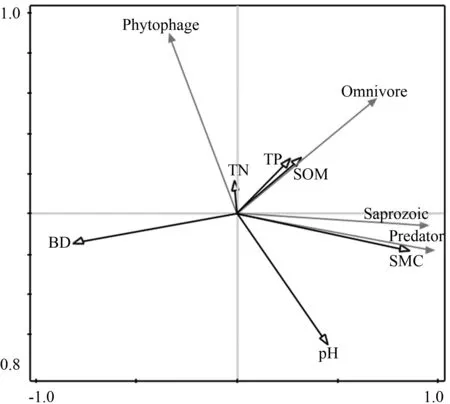
图4 宝天曼中小型土壤动物各功能类群与 土壤环境因子RDA分析注:Saprozoic—腐食性土壤动物;Phytophage—植食性土壤动物;Predator—捕食性土壤动物;Omnivore—杂食性土壤动物;BD—土壤容重;SMC—土壤含水率;pH—土壤酸碱度;SOM—土壤有机质;TN—土壤全氮;TP—土壤全磷
3 讨论
本研究发现,宝天曼中小型土壤动物功能类群以腐食性土壤动物为主,森林生态系统凋落物丰富,枯枝落叶和动物残体是腐食性土壤动物的主要食物来源,因此该区腐食性土壤动物个体、类群总数占据优势,这与唐本安、李娜等的研究结论相同[35-36].植食性土壤动物通过取食植物,为其他功能类群提供食物和能量来源,具有较高的优势度[37].捕食性土壤动物活动能力强,流动性较大,作为群落中的不稳定类群,均匀度和优势度指数最低[6].杂食性土壤动物因兼具多种食性,多样性和均匀度指数较高,可以看作是土壤动物对不稳定生境条件适应的表现,而生境内部越稳定也越有利于单一食性功能类群的生存[6].腐食性、杂食性土壤动物个体、类群总数随海拔上升呈“单峰型”分布模式,这与研究区中间海拔梯度水热充足、人为干扰程度低有关[38].相似性系数分析发现海拔是影响植食性土壤动物分布的重要限制性因素,可能是随着海拔的升高,样地植被类型由落叶阔叶林逐步变为针阔混交林,对植食性功能类群取食活动产生影响有关.
大量研究表明,土壤环境因素对土壤动物功能类群具有重要影响作用[1,39].研究区腐食性和捕食性土壤动物功能类群与土壤含水率呈显著正相关,这与Bengtsson等学者的研究结论相符[40].本研究还发现,土壤有机质、全氮、全磷等营养元素指标对不同功能群影响不同,如有机质和全磷对杂食性土壤动物影响比较明显,植食性土壤动物功能类群受全氮含量影响较大.孙儒泳研究指出,土壤pH值通常是影响土壤动物分布的限制性因素,本研究发现宝天曼中小型土壤动物各功能类群与pH值相关性并不显著,这可能与研究区各样地pH值差异较小有关[41].
通过划分土壤动物功能类群为深入研究土壤动物生态系统功能提供了科学的研究思路和研究方法,近年来已成为土壤生态学关注的热点[13].鉴于研究区生境的复杂性和土壤动物类群的多样性以及研究技术的限制,国内外当前对土壤动物功能类群并没有统一的划分标准,未来还需要运用新技术(如同位素示踪技术)和方法,对土壤动物食性进行细分,深入研究土壤生态系统功能及调控机制[6,36].
4 结论
本研究选取宝天曼自然保护区南坡海拔900~1700 m的8个样地为研究区,以中小型土壤动物为研究对象,基于土壤动物功能类群划分方法,探讨了宝天曼中小型土壤动物功能类群组成和结构特征,并分析了影响其分布格局的环境影响因素.
4.1 宝天曼自然保护区共捕获中小型土壤动物48类、3673只.基于食性差异,可划分为腐食性、植食性、捕食性、杂食性土壤动物4种功能类群,且其个体、类群总数以腐食性、杂食性为主,植食性和捕食性次之.
4.2 水平结构上,腐食性和杂食性个体、类群总数均随海拔上升呈先增加后减小的“单峰型”分布模式,植食性和捕食性土壤动物分布无明显规律;垂直分布上,各样地4种功能类群均表现出“表聚性”分布特征.
4.3 腐食性和杂食性土壤动物多样性指数、丰富度指数较高,植食性土壤动物优势度指数较高;海拔变化对4种功能类群多样性指数影响显著(P<0.05).Sorensen相似性指数表明研究区各样地腐食性、杂食性土壤动物组成和结构相近,海拔是影响植食性、捕食性土壤动物分布的重要限制性因素.
4.4 土壤环境因素对各功能类群影响程度具有差异.腐食性和捕食性功能类群与土壤含水率呈显著正相关,与土壤容重无显著相关性;杂食性土壤动物分布与有机质和全磷正相关;植食性土壤动物功能类群受全氮含量影响显著,与pH值无明显相关性.
