龙眼DlTRF2-like的克隆及表达模式分析
2021-01-13李浩然张树伟王金英黄小雄彭宏祥徐炯志潘介春朱建华谭春露李平丁峰
李浩然 张树伟 王金英 黄小雄 彭宏祥 徐炯志 潘介春 朱建华 谭春露 李平 丁峰





摘 要:以龙眼成熟叶片为材料,克隆龙眼MYB-related基因家族TRF2-like基因,命名为DlTRF2-like,并对其进行生物信息学和表达模式分析。DlTRF2-like基因属于MYB-related家族的TRF-like亚家族,ORF长度是909 bp,编码含有302个氨基酸残基的蛋白,预测该基因定位于细胞核,同源基因进化分析表明龙眼DlTRF2-like与阿月浑子PvTRF2-like亲缘关系最近。利用实时荧光定量PCR检测DlTRF2-like基因在‘四季蜜’龙眼不同组织及乙烯利和多效唑处理后不同时间的相对表达量。结果表明:DlTRF2-like在龙眼不同组织中均有表达,在花芽中表达量最高;乙烯利和多效唑处理后,DlTRF2-like基因的表达量明显高于对照,推测DlTRF2-like响应乙烯利和多效唑信号促进龙眼成花。
关键词:龙眼;MYB-related;龙眼成花;TRF-like亚家族;表达模式分析
中图分类号:S667.2 文献标识码:A
Abstract: The TRF2-like gene of MYB-related family was cloned from the mature leaves of longan and named DLTRF2-like. Bioinformatics and the expression pattern of DLTRF2-like were analyzed. DLTRF2-like gene belongs to the TRF-like subfamily of MYB-related family, with an ORF length of 909 bp, encoding a protein of 302 amino acids which was predicted to be located in the nucleus. Evolution analysis of homologous genes indicated that DLTRF2-like was most closely related to PvTRF2-like in Pistacia vera L. The expression pattern of DLTRF2-like in different tissues and leaves treated by ethephon and paclobutrazol was analyzed by real-time quantitative PCR. The results showed that DLTRF2-like was expressed in all the samples, and the highest expression level was found in flower buds. The expression level of DLTRF2-like gene in the leaves treated by ethephon and paclobutrazol was significantly higher than that of the control. The results indicated that DLTRF2-like might response to ethephon and paclobutrazol signal to promote the flowering of longan.
Keywords: longan; MYB-related; longan flowering; TRF-like subfamily; expression pattern analysis
DOI: 10.3969/j.issn.1000-2561.2021.12.012
龍眼(Dimocarpus longan Lour.)是无患子科龙眼属常绿果树,在我国栽培历史悠久。广西是龙眼主产地之一,采收期在7月中下旬至8月下旬。产期过于集中是我国龙眼生产面临的主要问题之一,为了有效调节龙眼产期,提高龙眼生产效益,生产上多采用修剪和化学调控等方法促进龙眼成花。化学调控主要采用植物生长调节剂和氯酸钾诱导龙眼成花,但是不同品种及不同地区的调控效果存在差异。‘四季蜜’可一年多次成花,常出现花果同期现象,生产上常用乙烯利、多效唑等处理,调控‘四季蜜’龙眼成花坐果时间。因此,解析龙眼成花的分子调控机制具有重要的理论和生产意义。
MYB转录因子参与调控植物的生长发育、生长代谢和细胞形态建成等生物学过程,在植物响应逆境胁迫[1-5]、花色素苷合成[6-7]、调控开花时间[8]等过程中发挥重要作用。根据高度保守的结构域的特征,可将MYB转录因子分为4个亚类,1R-MYB(MYB-related)、R2R3-MYB、3R-MYB、4R-MYB(4个R1/R2的重复)。1R-MYB转录因子亚类,是重要的端粒结合蛋白,在生长发育过程中起重要作用[9]。MYB-related家族可划分为5个亚家族:CAA1-like/R-R、TBP-like、CPC-like、I-BOX-like和TRF-like。各亚家族结构高度保守,其中CAA1-like/R-R亚家族成员结构均含有1个保守的氨基酸基序(SHAQK(Y/F)F);TBP-like亚家族成员结构均含有1个LKDKW(R/K)(N/T)基序;CAA1-like/R-R和TBP-like亚家族都含有红藻同源基因且最古老,而CPC-like和I-BOX- like亚家族在被子植物中才形成;其中CPC-like是从2R-MYB类的R3重复区域演化而来,I-BOX-like则是从含有2个MYB结构域的R-R类型的第一个MYB重复区域演化而来;TRF-like亚家族在进化过程中非常保守,且数量最少。
端粒结合蛋白TRF(telomere repeat binding factor, TRF)在人和动物中的功能研究较多,端粒结合蛋白TRF1、TRF2(telomere repeat-binding factor, TRF)与抑制因子激活蛋白Rap1(repressor activator protein 1)、TRF1相互作用的核蛋白2(TRF1-interacting nuclear protein 2, TIN2)、端粒保护蛋白1(protection of telomeres 1, POT1)和POT1结合蛋白1(telomere-binding protein POT1- interacting protein 1, TPP1)形成复合物,参与维持端粒DNA结构稳定和损伤修复、细胞的衰老凋亡、增殖和分化等[10]。TRF1和TRF2均以同源二聚体的形式结合到端粒双链DNA的YTAGGGTTR序列。TRF2和Ku70相互作用可以抑制端粒重组[11]。在拟南芥、水稻和玉米中,端粒长度与开花时间呈负相关,端粒越长,开花更早[12]。在拟南芥中,TRB通过结合telobox基序招募PRC2(polycomb repressive complexes)到靶基因介导H3K27me3表观修饰[13]。在植物春化过程中PRC2与VIN3(vernalization insensitive 3)共同参与开花抑制基因FLC的表观沉默,从而促进植物开花[14]。
为探索龙眼DlTRF2-like基因在成花调控中的生物学功能,本研究以龙眼叶片为材料克隆该基因,对其进行生物信息学分析,采用实时荧光定量PCR方法分析DlTRF2-like基因在‘四季蜜’龙眼不同组织及乙烯利和多效唑处理后不同时期的表达,为深入探索DlTRF2-like基因在龙眼成花过程中的作用机制提供参考依据。
1 材料与方法
1.1 材料
在广西大学农学院龙眼园基地采集‘四季蜜’龙眼的根、茎、叶片、叶柄、花芽、果肉、果皮和种子等8个不同组织的样品,‘四季蜜’龙眼喷施乙烯利和多效唑[15]后0、1、8、15、22(开花)、29、36、43 d等8个不同时期的叶片,以清水处理为对照。采样后擦拭叶片表面灰尘后,迅速置于液氮中速冻备用,各处理3次重复。
1.2 方法
1.2.1 总RNA的提取和反转录 样品总RNA用多糖多酚植物总RNA提取试剂盒(TIANGEN)提取,1%琼脂糖凝胶电泳检测提取质量,cDNA第一链的合成使用反转录试剂盒Prime ScriptTM RT Reagent Kit(Takara),随后将产物置于–20 ℃冰箱保存备用。
1.2.2 ‘四季蜜’龙眼DlTRF2-like基因的克隆 根据已公布的龙眼基因组信息,设计DlTRF2-like基因引物kl-DlTRF2-like F、kl-DlTRF2-like R1、kl-DlTRF2-like R2(表1),以合成的cDNA第一链为模板,采用Prime star Max Premix 2X高保真酶(Takara)进行ORF全长扩增。经1.5%琼脂糖凝胶电泳检测正确的条带使用试剂盒Gel Extraction Kit(Takara)回收,回收产物连接到pMD18-T载体后转化至大肠杆菌DH5α感受态细胞中,随机挑选阳性克隆菌液测序,获得DlTRF2-like基因的序列信息[16]。
1.2.3 DlTRF2-like蛋白的生物信息学分析 利用NCBI-CDS在线软件预测蛋白保守结构域,使用软件MEGA 5.0邻近相接法(neighbour joining)构建系统进化树,DNAMAN V6软件进行同源基因的多重序列比对。通过在线软件(http://www. soft-berry.com/berry.phtml?topic=index&group= programs &subgroup=proloc)对DlTRF2-like进行亚细胞定位预测。
1.2.4 龙眼DlTRF2-like基因的表达分析 DlTRF2- like基因的相对表达量,采用SYBR Premix Ex Taq(TaKaRa)在LightCycler 480实时荧光定量PCR仪(roche, switzerland)检测。反应程序如下95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火1 min,40个循环;循环结束后95 ℃ 5 s,60 ℃ 1 min进行溶解,50 ℃冷却30 s。以ActinF、ActinR为内参基因,3次重复,相对表达量的计算采用2–ΔΔCT法[17]。
2 结果与分析
2.1 DlTRF2-like基因的序列分析及和多重序列比對
‘四季蜜’龙眼DlTRF2-like基因ORF长度为909 bp(图1),编码的蛋白含有302个氨基酸残基(图2)。蛋白保守结构域分析表明,DlTRF2-like蛋白属于SANT-TRF/Myb-related超家族,在多肽链的N端存在Myb-related结合位点(图3),且存在含有SANT/MYB-like家族的端粒重复结合因子DNA结合域SANT_TRF,通过在线软件预测DlTRF2-like蛋白定位在细胞核。
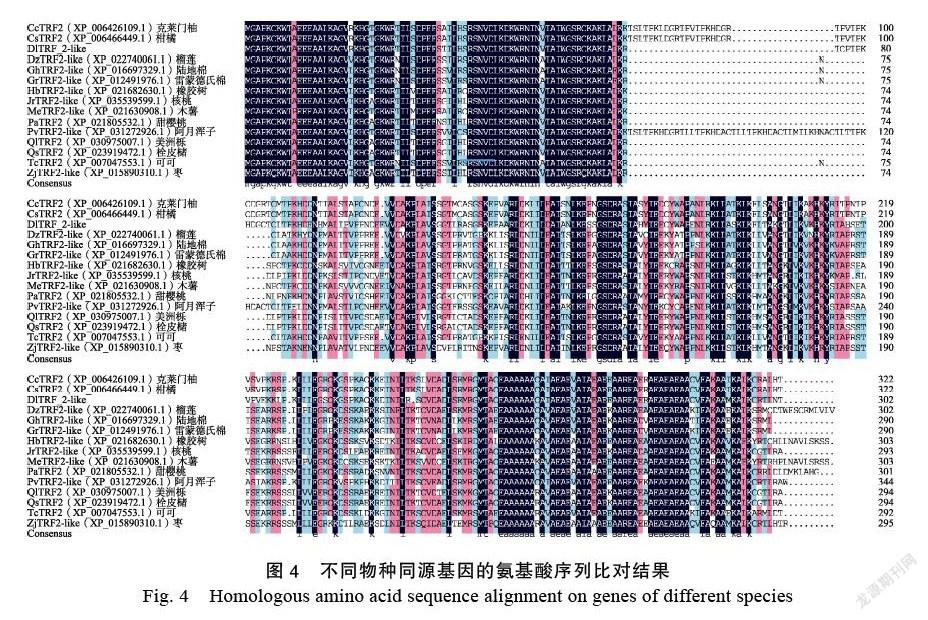
使用软件DNAMAN V6对龙眼DlTRF2-like和其他物种同源基因进行多重氨基酸序列比对(图4),发现龙眼DlTRF2-like中含有MYB- related基因家族典型保守序列LKDKWRN,确定该序列为MYB-related基因家族中的一员。
2.2 构建龙眼DlTRF2-like蛋白质进化树
将龙眼DlTRF2-like蛋白与NCBI中相似度较高的14种植物进行系统进化树的构建与分析[阿月浑子(Pistacia vera)PvTRF2-like(XP_ 031272926.1)、柑橘(Citrus sinensis)CsTRF2(XP_006466449.1)、克莱门柚(Citrus clementina) CcTRF2(XP_006426109.1)、橡胶树(Hevea brasiliensis)HbTRF2-like(XP_021682630.1)、美洲栎(Quercus lobata)QlTRF2(XP_030975007.1)、雷蒙德氏棉(Gossypium raimondii)GrTRF2-like(XP_012491976.1)、枣(Ziziphus jujuba)ZjTRF2- like(XP_015890310.1)、栓皮槠(Quercus suber)QsTRF2(XP_023919472.1)、陆地棉(Gossypium hirsutum)GhTRF2-like(XP_016697329.1)、可可(Theobroma cacao)TcTRF2(XP_007047553.1)、木薯(Manihot esculenta)MeTRF2-like(XP_ 021630908.1)、核桃(Juglans regia)JrTRF2-like(XP_035539599.1)、甜樱桃(Prunus avium)PaTRF2(XP_021805532.1)、榴莲(Durio zibethinus)DzTRF2-like(XP_022740061.1)。发现龙眼DlTRF2-like蛋白与阿月浑子PvTRF2- like的亲缘关系最接近,其次是柑橘CsTRF2和克莱门柚CcTRF2,它们的功能可能更为相似(图5)。
2.3 DlTRF2-like基因的表达分析
2.3.1 DlTRF2-like基因‘四季蜜’龙眼不同组织中的表达 实时荧光定量PCR结果表明(图6),DlTRF2-like基因在‘四季蜜’龙眼不同组织中的表达存在显著性差异,DlTRF2-like基因在花芽中的表达量最高,是根中表达量的8倍;其次是叶柄,表达量约为根中的6倍;然后是叶片,表达量是根中的3.3倍,DlTRF2-like基因在茎、果肉、果皮和种子的表达量与根相近,说明DlTRF2-like基因主要在‘四季蜜’龙眼的叶片和花芽中发挥重要功能。
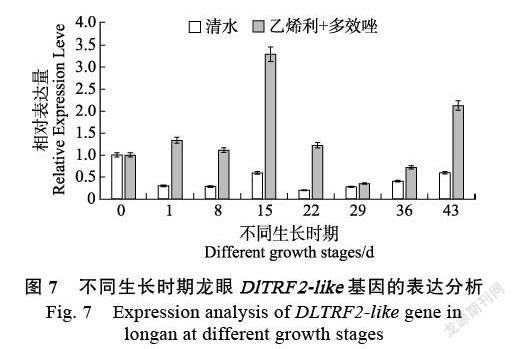
2.3.2 喷施乙烯利和多效唑的叶片中DlTRF2- like基因的表达情况 喷施乙烯利和多效唑后DlTRF2-like基因的表达量与对照存在较大差异(图7),喷施乙烯利和多效唑后的表达量明显高于同时期的清水对照,说明喷施乙烯利和多效唑可以诱导DlTRF2-like基因在‘四季蜜’龙眼叶片中的表达。喷施乙烯利和多效唑后15 d,DlTRF2-like基因表达量最高,其次是43 d。DlTRF2-like基因在开花前的0、1、8 d表达量相近,在15 d表达量迅速上升至3倍,喷施乙烯利和多效唑后22 d开花,而清水对照未开花,且22 d表达量下降至1倍水平,说明DlTRF2-like基因在临近开花时起重要作用。43 d末花期表达量增高原因暂时不明确。
3 讨论
本研究获得了龙眼DlTRF2-like基因,DlTRF2-like属于TRF-like亚家族,全长909 bp,编码302个氨基酸,含有一个Myb-related保守结构域LKDKWRN。亚细胞定位预测显示DlTRF2-like蛋白定位于细胞核中。NCBI比对发现龙眼DlTRF2-like与阿月浑子PvTRF2-like的亲缘关系最接近,其次是柑橘CsTRF2和克莱门柚CcTRF2,目前暂无阿月浑子PvTRF2-like、柑橘CsTRF2和克莱门柚CcTRF2功能相关报道,龙眼DlTRF2-like基因功能有待进一步分析。
PcG(polycomb group)介导的表观修饰在动植物的发育过程中发挥重要作用。拟南芥PcG与PWO1(pwwp domain interactor of polycombs1)相互作用调控开花和发育[18]。在拟南芥中TRB可以结合telobox基序,并与PcG的核心催化酶CLF(curly leaf)和SWN(swinger)相互作用,招募PRC2介导靶基因的表观修饰[13]。
多效唑和乙烯利具有抑制果树营养生长和促进成花的作用[19-20]。在龙眼栽培中,乙烯利有明显的促花和增加开花数的作用,高浓度乙烯利(CEPA)能在冬季有效地调控龙眼冬梢促进花芽分化,低浓度乙烯利在龙眼花序迅速分花期(2月下旬)使用能使多年不开花的成年树开花结果[21]。热带生态型龙眼‘四季蜜’是一个易于调控成花的龙眼品种,喷施多效唑和乙烯利具有调控其成花的作用,且使用浓度范围较广,多种浓度组合以及不同施用次数都能促进开花[15]。本研究分析了‘四季蜜’龙眼不同组织中DlTRF2-like基因表达,结果显示它在花芽中表达量最高,其次是叶柄、叶片。有研究发现MdFT基因在苹果花、叶片和幼果中的表達均高于种子等其他组织[22],桑树MaFT基因在成熟叶的表达高于花、果实、嫩叶、腋芽顶芽等组织[23]。而FT基因在植物花形成过程中起关键作用[24],故推测龙眼DlTRF2- like基因可能在‘四季蜜’龙眼花芽中发挥重要作用。根据定量结果,DlTRF2-like基因在喷施乙烯利和多效唑的表达量明显高于在清水对照中的表达量,说明该基因可能参与乙烯利和多效唑促进龙眼成花调控,促进花芽分化。但该基因具体作用机制尚不明确,有待进一步研究证明。
参考文献
[1] Zhao P C, Hou S L, Guo X F, et al. A MYB-related tran-scription factor from sheepgrass, LcMYB2, promotes seed germination and root growth under drought stress[J]. BMC Plant Biology, 2019, 19(14): 237-251.
[2] Yin X M, Cui Y C, Wang M L, et al. Overexpression of a novel MYB-related transcription factor, OsMYBR1, confers improved drought tolerance and decreased ABA sensitivity in rice[J]. Biochemical and Biophysical Research Commu-nications, 2017, 490(4): 1355-1361.
[3] Cheng L Q, Li X X, Huang X, et al. Overexpression of sheepgrass R1-MYB transcription factor LcMYB1 confers salt tolerance in transgenic arabidopsis[J]. Plant Physiology and Biochemistry, 2013, 70: 252-260.
[4] Li T, Sun J K, Bi Y P, et al. Overexpression of an MYB- Related Gene FvMYB1 from Fraxinus velutina increases to-lerance to salt stress in transgenic tobacco[J]. Plant Growth Regulation, 2016, 35(3): 632-645.
[5] Xiong H Y, Li J J, Liu P L, et al. Overexpression of Os-MYB48-1, a novel MYB-Related transcription factor, en-hances drought and salinity tolerance in rice[J]. PloS One, 2014, 9(3): e92913.
[6] Aly R, Lati R, Bari V K, et al. Use of a visible reporter marker-myb-related gene in crop plants to minimize herbi-cide usage against weeds[J]. Plant Signaling & Behavior, 2019, 14(4): e1581558.
[7] Kobayashi S, Ishimaru M, Hiraoka K, et al. Myb-related genes of the Kyoho grape (Vitis labruscana) regulate antho-cyanin biosynthesis[J]. Planta, 2002, 215(6): 924-933.
[8] Zhang L C, Liu G X, Jia J Z, et al. The wheat MYB-related transcription factor TaMYB72 promotes flowering in rice [J]. Journal of Integrative Plant Biology, 2016, 58(8): 701- 704.
[9] 杜 海. 植物MYB转录因子家族的分子进化机制及调控类黄酮生物合成MYB基因的鉴定[D]. 雅安: 四川农业大学, 2013.
[10] Titia D L. Shelterin: The protein complex that shapes and safeguards human telomeres[J]. Genes & development, 2005, 19(18): 2100-2110.
[11] Song K, Jung D, Jung Y S, et.al. Interaction of human Ku70 with TRF2[J]. FEBS Letters, 2000, 481: 81-85.
[12] Young C J, Abdulkina L R, Yin J, et al. Natural variation in plant telomere length is associated with flowering time[J]. Plant Cell, 2021, 33(4): 1118-1134.
[13] Zhou Y, WangY J, Krause K, et al. Telobox motifs recruit CLF/SWN–PRC2 for H3K27me3 deposition via TRB factors in Arabidopsis[J]. Nature Genetics, 2018, 50(5): 638-644.
[14] Zhao Y, Antoniou-Kourounioti R L, Calder G, et al. Tem-perature-dependent growth contributes to long-term cold sensing[J]. Nature, 2020, 583: 825-829.
[15] 朱建华, 黄凤珠, 徐 宁, 等. ‘四季蜜’龙眼成花调控技术研究[J]. 中国农学通报, 2011, 27(25): 288-293.
[16] 李 琳, 丁 峰, 潘介春, 等. 龙眼DlZAT10基因克隆与植物超表达载体构建[J]. 热带作物学报, 2021, 42(1): 33- 39.
[17] Livak K J, Schmittgen T D. Analysis of relative gene ex-pression data using real-time quantitative PCR and the 2 (-delta delta C (T)) method. Methods, 2001, 25(4): 402-408.
[18] Hohenstatt M L, Mikulski P, Komarynets O, et al. PWWP- DOMAIN INTERACTOR OF POLYCOMBS1 interacts with Polycomb-group proteins and histones and regulates Arabidopsis flowering and development[J]. Plant Cell, 2018, 30(1): 117-133.
[19] 叶明儿. 植物生长调节剂在果树上的应用[M]. 北京: 化学工业出版社, 2011.
[20] 卢美英, 徐炯志, 欧世金, 等. 荔枝龙眼杧果沙田柚控梢促花保果综合调控技术[M]. 北京: 金盾出版社, 2009.
[21] 陈香玲. 乙烯利对龙眼成花的生理生化作用研究[D]. 南宁: 广西大学, 2005.
[22] 郑小一, 王三红, 张计育, 等. 苹果FT同源基因MdFT的表达特性[J]. 江苏农业学报, 2011, 27(2):390-395.
[23] 李四军. 桑树FT基因的克隆及表达分析[D]. 杨凌: 西北农林科技大学, 2013.
[24] 张 婧. 高等植物开花基因FT研究进展[J]. 现代农业科技, 2012(3): 31-32.
責任编辑:沈德发
