东南亚兰科植物的物种多样性、生活习性及其传粉系统
2021-01-12张哲任明迅向文倩宋希强
张哲 任明迅 向文倩 宋希强





摘 要:兰科(Orchidaceae)植物广布于除两极和极端沙漠地区外的各种陆地生态系统,包括5个亚科800多属28 000多种。东南亚地区兰科植物种数约占世界的1/3,是兰科植物生物多样性热点区域之一。通过查阅文献及书籍等资料,该文系统整理了东南亚兰科植物物种种类及其扩散演化历史,并对其生活习性和传粉系统进行了归类。结果表明:(1)东南亚兰科植物8 855种,分属5亚科17族26亚族240属;(2)主要生活型为附生的有127属6 000种以上,地生97属2 000种以上,腐生13属约100种,藤本4属40余种;(3)根据整理出的东南亚79个属的兰科植物传粉系统发现,有44个属含有自动自交的物种,具报酬物的传粉系统有花粉(仅见于拟兰亚科)、芳香类物质(仅见于香荚兰亚科)和花蜜(5个亚科均有)等报酬物类型。欺骗性传粉系统广泛存在于各个亚科,包括食源性欺骗、性拟态、繁殖地拟态和信息素拟态等类型。东南亚兰科植物在物种、生活习性及传粉系统都展现出极高的多样性,对这些生物学特点的总结将为兰科植物的保育提供一定的理论基础和本底资料。
关键词:东南亚,兰科植物,物种多样性,传粉生物学,保育
中图分类号:Q949.71
文献标识码:A
文章编号:1000-3142(2021)10-1683-16
Abstract:Orchidaceae,widely distributed in various terrestrial ecosystems except for Antarctica,Arctic and extreme desert areas,comprises more than 28 000 species in more than 800 genera of 5 subfamilies. As one of the hotspots of orchid biodiversity in the world,Southeast Asia accounted for about 1/3 of all orchid species. In this paper,we reviewed the species diversity,evolution and dispersal history,as well as classified the habits and pollination systems of orchid species in Southeast Asia. The results are as follows:(1) A total of 8 855 orchid species,which belongs to 5 subfamilies,17 tribes,26 subtribes and 240 genera in Southeast Asia,were enumerated and evaluated. (2) The main habits of orchids in Southeast Asia include more than 6 000 epiphytic species of 127 genera,more than 2 000 terrestrial species of 97 genera,about 100 saprophytic species of 13 genera and more than 40 vine species of 4 genera. (3) According to the pollination system of 79 genera of Orchidaceae in Southeast Asia,44 genera contained automatic self-pollination species. Rewarding pollination systems involve the forms of pollen (only found in subfamily Apostasioideae),fragrance oils (only found in subfamily Vanilloideae) and nectar (found in all five subfamilies) as rewards. Additionally,deceptive pollination systems exist widely in all five subfamilies,including food-deceptive system,sexual mimicry,shelter mimicry,oviposition-site mimicry and pheromone mimicry. Orchidaceae in Southeast Asia show a high diversity of species,habits and pollination systems. The summary of orchid biological characteristics provides some theoretical foundations and context information for the conservation.
Key words:Southeast Asia,Orchidaceae,species diversity,pollination biology,conservation
蘭科(Orchidaceae)植物广布于除两极和极端沙漠地区外的各种陆地生态系统,是被子植物中与菊科并列的最大科,有800多属28 000多种,约占世界维管束植物总数的10%(Joppa et al.,2011)。目前的研究显示,兰科植物可能起源于早白垩纪的澳大利亚地区,此时非洲、印度和马达加斯加已与南极洲和澳大利亚分离,而澳大利亚和南美洲通过南极洲相连(Givnish et al.,2015,2016)。约90 Mya (million years ago)时,兰科植物从澳大利亚地区穿越南极洲向新热带区扩散,逐渐分化出五个亚科,包括拟兰亚科(Apostasioideae)、香荚兰亚科(Vanilloideae)、杓兰亚科(Cypripedioideae)、兰亚科(Orchidoideae )和树兰亚科(Epidendroideae) (Givnish et al.,2015,2016)。
东南亚地区是世界三大热带雨林分布区之一和生物多样性最高的区域之一,以约占全球陆地5%的面积分布着全球20%~25%的高等植物(Brooks et al.,2006; Buerki et al.,2014)。东南亚地区兰科植物预计超过10 000种,约占世界兰科植物的1/3,是世界上兰科植物最为丰富的区域之一。现有研究显示,历史上兰科植物的长距离扩散速率相对其他植物类群较低,从而限制了物种在跨洋大洲间的基因交流,加剧了物种在独立大陆板块内的分化(Givnish et al.,2016)。东南亚复杂的地质历史,如大陆板块碰撞和连接、陆地隆起形成高山、火山喷发、海平面的反复波动等条件下,加剧了这一区域物种分化和多样性的提高(Thomas et al.,2012; Guo et al.,2012,2015 )。此外,蘭科植物多样的花部特征和传粉系统(Cozzolino,2005; Schiestl,2009; Ramírez et al.,2011)、附生习性及与之相关的景天酸代谢途径(CAM,crassulacean acid metabolism pathway)(Gravendeel et al.,2004; Silvera et al.,2009)等,都被认为驱动了兰科植物的物种形成和分化。因此,对东南亚地区兰科植物的物种多样性、生活习性及其传粉系统方面的总结有助于了解这一地区兰科植物的特点,对于兰科植物的保育具有重要的理论和实践意义。
根据地块历史及植物区系属性,东南亚地区包括以下4大区域(谭珂等,2020)。(1)印度-缅甸区:主要包括中南半岛、海南岛、云南南部、广西西南部及广东南部沿海;(2)巽他区:主要包括马来半岛、婆罗洲、苏门答腊岛等;(3)华莱士区:主要由苏拉威西岛、爪哇岛、小巽他群岛、马鲁古群岛等组成;(4)菲律宾群岛:包括吕宋岛、米沙鄢和棉兰岛等7 000多个大小岛屿。该文根据Kew Garden的世界植物名录(World Checklist of Selected Plant Families) (http://wcsp.science.kew.org/)公布的兰科植物物种数及分布信息,以及GBIF (Global Biodiversity Information Facility,http://www.gbif.org/)收集兰科植物的分布地点信息,并查阅书籍或文献(Pridgeon et al.,1997,1999,2001,2003,2005,2009,2014; Chen et al.,2009; Chase et al.,2015; Givnish et al.,2015,2016; Angiosperm Phylogeny Group et al.,2016)获取兰科的分类系统、物种多样性和物种系统发育位置,对东南亚区域内的兰科植物名录进行整理和分析,并对其生活习性和传粉生物学进行概述。兰科植物的起源由于缺乏化石证据,不同作者基于不同的证据,在起源时间和地点等方面都有不同观点,为保证统一性,本文采用Givnish et al.(2015,2016)的研究成果作为科属之间的起源断代参考。
1 东南亚兰科植物起源及其多样性
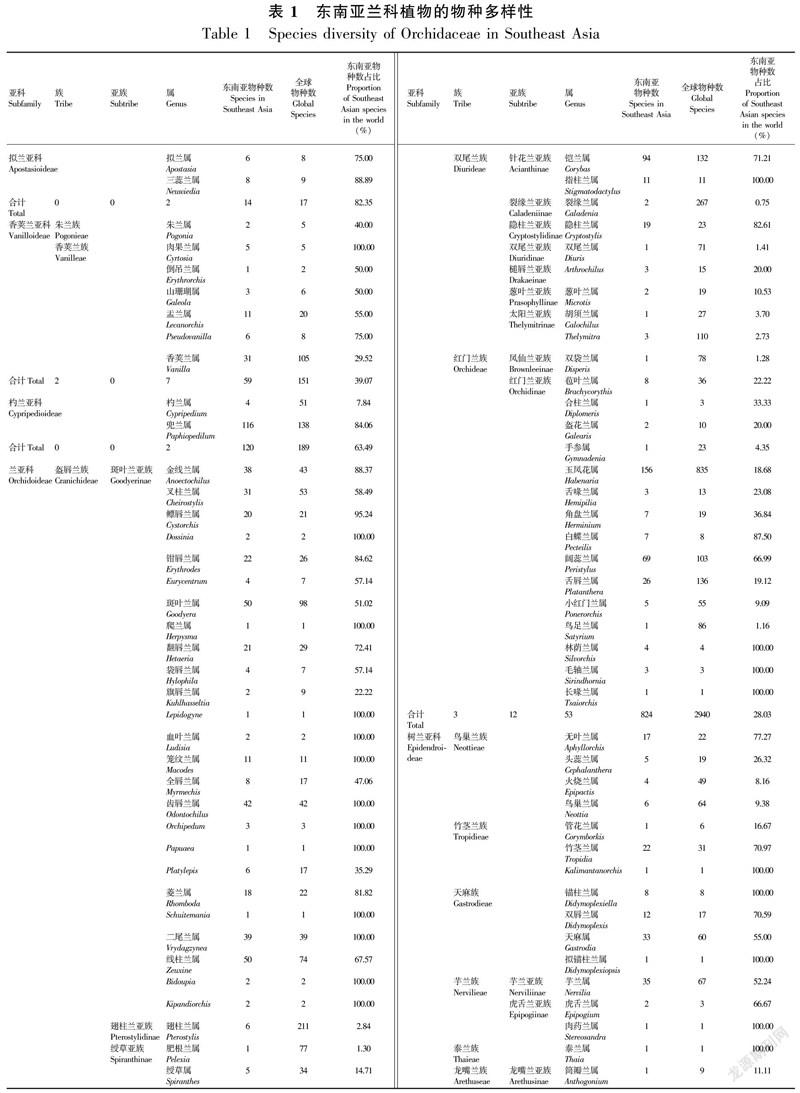
本文共整理出东南亚兰科植物8 855种,分属5亚科17族26亚族240属。东南亚地区的兰科植物表现出极强的地域特有性,有118个属下所有种在东南亚地区均有分布,有197属的种类占全球分布种数在50%以上(表1)。
1.1 拟兰亚科(Apostasioideae)
拟兰亚科是兰科的基部类群,90 Mya时最早于澳大利亚地区从兰科祖先类群中分化出来。在25 Mya时,由澳大利亚通过新几内亚并穿过华莱士线到达东南亚地区(Givnish et al.,2015,2016)。东南亚是世界拟兰亚科的分布多样性中心,有2属14种,主要分布于婆罗洲中部和北部、中南半岛西部、马来半岛南部和苏门答腊岛。
该亚科全球分布有2属17种,主要分布于东南亚、日本和澳大利亚北部的湿润地区。其中拟兰属(Apostasia)世界分布有8种,除深圳拟兰(A. shenzhenica)和A. fogangica外,其余6种在东南亚地区均有分布;三蕊兰属(Neuwiedia)世界分布9种,除麻栗坡三蕊兰(N. malipoensis)外,其余8种在东南亚均有分布(表1)。
1.2 香荚兰亚科(Vanilloideae)
香荚兰亚科起源于84 Mya时的新热带区,其下的朱兰族(Pogonieae)于44 Mya从新热带区向北美洲扩散,分化出Cleistesiopsis、Isotria和朱兰属(Pogonia),随后这些类群于11 Mya向欧亚大陆扩散;本亚科香荚兰族(Vanilleae)自新热带区起源后,于64~59 Mya通过长距离扩散穿越太平洋到达新喀里多尼亚,分化出Clematepistephium和Eriaxis; 而Pseudovanilla和肉果兰属(Cyrtosia)于61 Mya从新热带区长距离扩散至澳大利亚和东南亚地区;随后,肉果兰属进一步扩散到东亚地区,Pseudovanilla又于31~6 Mya扩散至波纳佩和斐济(Givnish et al.,2016)。香荚兰属(Vanilla)于61 Mya自新热带区分化形成,于26~18 Mya扩散至非洲,于13 Mya自非洲扩散至印度洋区域又于12~4 Mya自新热带区扩散至加勒比海区域(Givnish et al.,2016)。
东南亚是世界香荚兰亚科分布中心之一,共有2族7属59种(表1),主要分布在马来半岛、婆罗洲北部和菲律宾中部。
1.3 杓兰亚科(Cypripedioideae)
杓兰亚科包括5属,包括杓兰属(Cypripedium)、兜兰属(Paphiopedilum)、Mexipedium、Phragmipedium和Selenipedium。该亚科物种广泛分布于欧亚大陆的温带至热带地区及南北美洲地区,其中杓兰属主要分布于北半球的温带和亚热带地区,有些种类延伸到北美热带地区;Mexipedium、Phragmipedium和Selenipedium则主要集中在新热带区;兜兰属主要集中在旧热带区。
目前的研究表明,大陆分裂及随后的冰期气候变冷导致了该亚科物种的不连续分布,杓兰属是该亚科的基部类群,于76 Mya左右在新热带区分化形成并长距离扩散至欧亚大陆,少量到达东南亚,随后又扩散返回北美洲数次。Selenipedium、Phragmipedium和Mexipedium、分别于约31、28、21 Mya分化形成。兜兰属于46 Mya经历了长距离扩散,可能的扩散路线为新热带区—穿越白令陆桥—东亚—东南亚,并在东南亚地区分化出众多种类(Givnish et al.,2016)。
东南亚杓兰亚科有2属,包括兜兰属和杓兰属。兜兰属世界分布有130多种,东南亚是兜兰属的世界多样性中心,有116种,主要分布于中国西南部至越南中部、婆罗洲中部和菲律宾中部。杓兰属全球分布有51种,主要分布于北半球温带和亚高山带至美洲中部,东南亚分布仅4种(表1)。
1.4 兰亚科(Orchidoideae)
兰亚科是兰科中物种数量仅次于树兰亚科的类群,于64 Mya在新热带区与树兰亞科分化开来,包括银钟兰族(Codonorchideae)、盔唇兰族(Cranichideae)、双尾兰族(Diurideae)和红门兰族(Orchideae)。该亚科中的兰族沿新热带区—非洲—欧亚大陆的路线扩散,最后到达日本和北美洲;银钟兰族仅在南美洲分布;双尾兰族沿新热带区—澳大利亚—东南亚、新西兰和新喀里多尼亚的路线扩散;盔唇兰族的物种主要在新热带区分化形成,一部分扩散至北美地区,如盔唇兰属(Cranichis)和绶草属(Spiranthes),另一部分向澳大利亚和太平洋区域扩散,如Pachyplectron、翅柱兰属(Pterostylis)及它们的近缘类群(Givnish et al.,2016)。
东南亚分布有兰亚科共3族12亚族53属824种(表1)。
1.4.1 盔唇兰族(Cranichideae) 含3个亚族,28属395种。①斑叶兰亚族(Goodyerinae),25属381种,包括斑叶兰属(Goodyera)、线柱兰属(Zeuxine)、齿唇兰属(Odontochilus)、二尾兰属(Vrydagzynea)、金线兰属(Anoectochilus)、叉柱兰属(Cheirostylis)、菱兰属(Rhomboda)、钳唇兰属(Erythrodes)、鳔唇兰属(Cystorchis)、翻唇兰属(Hetaeria)等。②翅柱兰亚族(Pterostylidinae),仅含翅柱兰属(Pterostylis)8种。③绶草亚族(Spiranthinae),仅含绶草属(Spiranthes)和肥根兰属(Pelexia)2属6种。
1.4.2 双尾兰族(Diurideae) 含7个亚族,9属138种。①针花兰亚族(Acianthinae),2属105种。②裂缘兰亚族(Caladeniinae),仅含裂缘兰属(Caladenia)2种。③隐柱兰亚族(Cryptostylidinae),仅含隐柱兰属(Cryptostylis)19种。④双尾兰亚族(Diuridinae),仅含双尾兰属(Diuris)1种。⑤槌唇兰亚族(Drakaeinae),仅含Arthrochilus 3种。⑥葱叶兰亚族(Prasophyllinae),仅含葱叶兰属(Microtis)2种。⑦太阳兰亚族(Thelymitrinae),仅Thelymitra和胡须兰属(Calochilus)共4种。
1.4.3 红门兰族(Orchideae) 含2个亚族,16属306种。①凤仙兰亚族(Brownleeinae),仅含双袋兰属(Disperis)1种。②红门兰亚族(Orchidinae),15属305种,包括玉凤花属(Habenaria)、阔蕊兰属(Peristylus)、舌唇兰属(Platanthera)、苞叶兰属(Brachycorythis)、角盘兰属(Herminium)等。
1.5 树兰亚科(Epidendroideae)
树兰亚科是兰科中的最大亚科,与兰亚科于64 Mya时分化形成。基部类群鸟巢兰族(Neottieae)最先从新热带区扩散至欧亚大陆、东南亚和北美洲,目前主要分布于北半球的温带和亚热带地区,个别种类扩散到热带高山地区(Pridgeon et al.,2005; Chen et al.,2009; Zhou & Jin,2018)。箬叶兰族(Sobralieae)、垂帽兰族(Triphoreae)、竹茎兰族(Tropideae)、芋兰族(Nervileae)也由祖先类群从新热带区长距离扩散至东南亚地区分化形成。现存的树兰族(Epidendreae)的大部分类群均为祖先类群于30 Mya从东南亚回迁至新热带区后逐渐分化形成,包括布袋兰亚族(Calypsinae)的Coelia、禾叶兰亚
族(Agrostophyllinae)、拟白及亚族(Bletiinae)、蕾丽兰亚族(Laeliinae)、腋花兰亚族(Pleurothallidinae)、蔺叶兰亚族(Ponerinae)及不包括兰亚族(Cymbidinae)外的兰族(Cymbideae)类群;而兰亚族一枝于17 Mya时从东南亚开始扩散,分别到达澳大利亚、太平洋海域和欧亚大陆地区;禾叶兰亚族(Agrostophyllinae)中的Earina于30 Mya从东南亚扩散至太平洋区域;布袋兰亚族中的大部分类群于32 Mya扩散至北美洲地区,随后从北美洲扩散回欧亚大陆和东南亚地区(Givnish et al.,2016)。龙嘴兰族(Arethuseae)于15 Mya从东南亚长距离扩散至北美洲,随后又扩散回东南亚地区。万代兰族(Vandeae)中的彗星兰亚族(Angraecinae)是由祖先类群于21 Mya从东南亚长距离扩散类群至非洲分化而形成(Givnish et al.,2016)。吻兰族(Collabieae)、柄唇兰族(Podochileae)、沼兰族(Malaxideae)中的石斛亚族(Dendrobiinae)也发生过多次较长距离的扩散,从东南亚扩散至澳大利亚和太平洋海域(Givnish et al.,2016)。
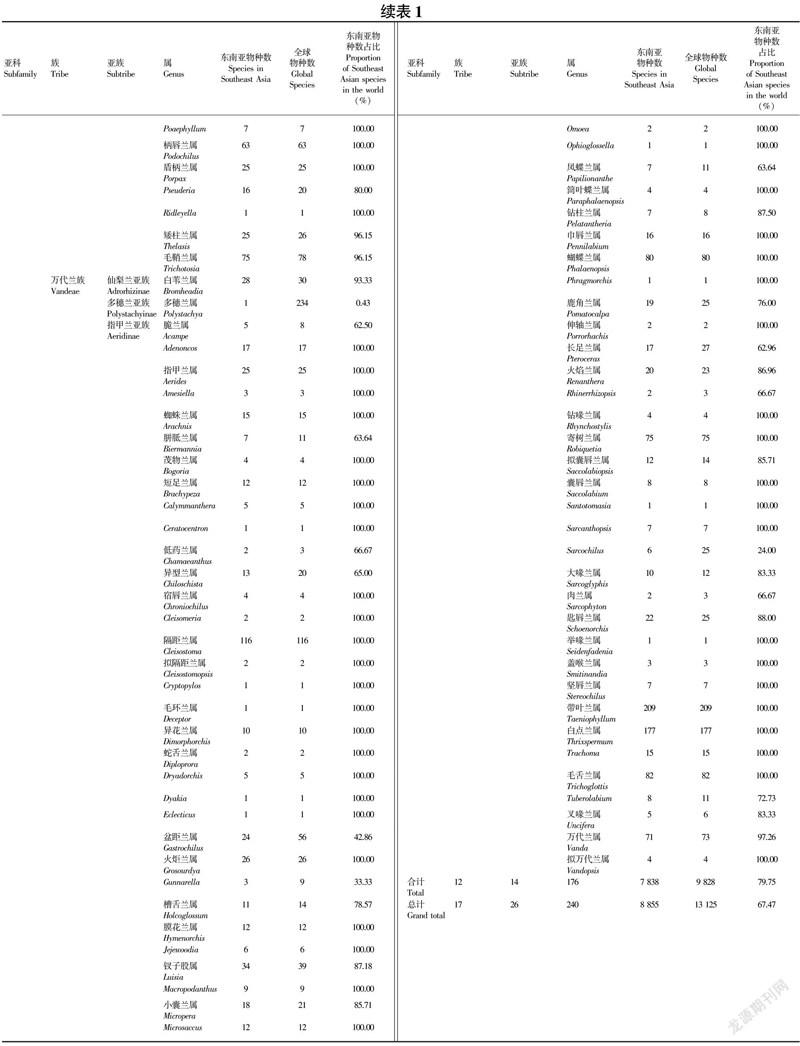
东南亚树兰亚科共12族14亚族176属7 838种(表1)。
1.5.1 鸟巢兰族(Neottieae) 含无叶兰属(Aphyllorchis)、鸟巢兰属(Neottia)、头蕊兰属(Cephalanthera)和火烧兰属(Epipactis)共4属32种。
1.5.2 竹茎兰族(Tropidieae) 含竹茎兰属(Tropidia)、管花兰属(Corymborkis)和甜薯兰属(Kalimantanorchis)共3属24种。
1.5.3 天麻族(Gastrodieae) 含天麻属(Gastrodia)、双唇兰属(Didymoplexis)、锚柱兰属(Didymoplexiella)和拟锚柱兰属(Didymoplexiopsis)共4属54种。
1.5.4 芋兰族(Nervilieae) 含2个亚族,3属38种。包括①芋兰亚族(Nerviliinae):仅含芋兰属(Nervilia)35种。②虎舌兰亚族(Epipogiinae):虎舌兰属(Epipogium)和肉药兰属(Stereosandra)共2属3种。
1.5.5 泰兰族(Thaieae) 仅含单种属泰兰属(Thaia)1种。
1.5.6 龙嘴兰族(Arethuseae) 含①龙嘴兰亚族(Arethusinae):包括筒瓣兰属(Anthogonium)和竹叶兰属(Arundina)共2属2种。②贝母兰亚族(Coelogyninae):包括足柱兰属(Dendrochilum)、贝母兰属(Coelogyne)种、球序兰属(Glomera)、石仙桃属(Pholidota)、Chelonistele等共22属764种。
1.5.7 沼兰族(Malaxideae) 含2个亚族,10属3 691种。①石斛亚族(Dendrobiinae):包括兰科中最大的两个属,石豆兰属(Bulbophyllum)和石斛属(Dendrobium)共2属3 042种。②沼兰亚族(Malaxidinae):包括沼兰属(Crepidium)、鸢尾兰属(Oberonia)、羊耳蒜属(Liparis)、覆苞兰属(Stichorkis)、原沼兰属(Malaxis)等共8属785种。
1.5.8兰族(Cymbidieae) 含2个亚族,10属153种。①兰亚族(Cymbidiinae):包括兰属(Cymbidium)、合萼兰属(Acriopsis)、斑被兰属(Grammatophyllum)、盒足兰属(Thecopus)、紫舌兰属(Porphyroglottis)和盒柱兰属(Thecostele)共6属88种。②美冠兰亚族(Eulophiinae):包括Claderia、双足兰属(Dipodium)、美冠兰属(Eulophia)和地宝兰属(Geodorum)共4属74种。
1.5.9树兰族(Epidendreae) 含3个亚族,6属138种。①腋花兰亚族(Pleurothallidinae):包括Brachionidium仅1种。②布袋兰亚族(Calypsinae):包括杜鹃兰属(Cremastra)、山兰属(Oreorchis)、筒距兰属(Tipularia)和宽距兰属(Yoania)共4属8种。③禾叶兰亚族(Agrostophyllinae):仅含禾叶兰属(Agrostophyllum)129种。
1.5.10吻兰族(Collabieae ) 包括虾脊兰属(Calanthe)、鹤顶兰属(Phaius)、苞舌兰属(Spathoglottis)、卷舌兰属(Plocoglottis)、带唇兰属(Tainia)等共19属366种。
1.5.11 柄唇兰族(Podochileae) 包括馥兰属(Phreatia)、牛齿兰属(Appendicula)、牛角兰属(Ceratostylis)、毛兰属(Eria)、苹兰属(Pinalia)、毛鞘兰属(Trichotosia)、柄唇兰属(Podochilus)、八雄兰属 (Octarrhena)、拟毛兰属(Mycaranthes)、拟石斛属(Oxystophyllum)等共26属1 277种。
1.5.12 万代兰族(Vandeae)
含3个亚族,69属1 348种。①仙梨兰亚族(Adrorhizinae):仅含白苇兰属(Bromheadia)28种。②多穗兰亚族(Polystachyinae):仅含多穗兰属(Polystachya)1种。③指甲兰亚族(Aeridinae):包括带叶兰属(Taeniophyllum)、白点兰属(Thrixspermum)、隔距兰属(Cleisostoma)、蝴蝶兰属(Phalaenopsis)、毛舌兰属(Trichoglottis)、寄树兰属(Robiquetia)、万代兰属(Vanda)、钗子股属(Luisia)等共67属1 319种。
2 东南亚兰科植物的生活习性
在陆地生态系统中,被子植物生活习性的多样性决定着植物类群的物种多样性、种群扩张程度及其在种群中的优势度(Ricklefs & Renner,1994; Tiffney & Mazer,1995)。兰科植物的生活习性多样,大约2/3的兰科植物都属于附生类,其次是地生类,少数属于腐生类,极少数种类具有攀援藤本的习性(Sosa et al.,2016)。大部分分布于温带的兰科植物都是地生习性,而热带地区具有最高的物种多样性,而且80%以上的种类都是附生习性(Sosa et al.,2016)。
附生习性兰科植物在属水平上的平均物种数要多于陆生习性的兰科植物(Gravendeel et al.,2004),并且相对陆生兰科植物的物种形成和分化速率也更高(Givnish et al.,2016)。附生習性是兰科植物极为重要的进化特征,影响着兰科植物物种的生存、形成、扩散和分化。主要体现在几大方面:①附生习性促进了兰科植物的“开疆拓土”和物种的保持。因为在树干和树枝上很少有维管束植物定植,附生兰科植物具有极小的竞争压力,保证了自己的生态位。②附生习性能够保持物种较高的遗传多样性。一是因为在森林内部树木表面积要远远大于地表,附生植物相对陆生植物具有更大的生长和扩散空间;二是不同树冠内部温湿度均不相同,能够促使物种在小尺度上出现不同的变异和分化。③附生习性往往与降雨和湿度有关,森林中雾气的沉积和蒸发率随海拔梯度的上升和地形变化创造出不同的生境条件,从而在更大尺度上导致隔离并加速物种的分化。④兰科植物种子可以扩散非常长的距离,但能够定植下来的却很少,促进了物种在不同生境中的分化(Givnish et al.,2015)。附生习性在树兰亚科新近类群(如树兰族、兰族、龙嘴兰族等)中至少进化过一次,发生时间不晚于35 Mya,而这一习性随后在部分类群中出现过至少三次的返祖事件,包括拟白芨亚族、布袋兰亚族和龙嘴兰亚族;其他亚科的部分类群,如杓兰亚科兜兰属和Phragmipedium,以及兰亚科双袋兰属和Eurystyles的一些物种也进化出了附生习性(Givnish et al.,2016)。
景天酸代谢途径(CAM,Crassulacean Acid Metabolism Pathway)与附生习性密切相关。附生兰科植物中一半以上的物种是景天酸代谢途径(Lüttge,2004; Gravendeel et al.,2004)。C3光合代谢途径(C3 Photosynthetic Pathway)是兰科的祖先特质,伴随着几个亚科的平行进化,CAM光合代谢途径在兰科植物中至少独立进化过10 次,并发生过几次返祖事件(Silvera et al.,2009)。现有研究表明,C3光合代谢途径包含几乎所有参与CAM光合代谢途径的基因,后者很可能是由前者经过调控和表达重组进化而来(Westeberhard et al.,2011)。现有证据表明,树兰亚科发生了大量CAM光合代谢途径的辐射分化事件,可能与65 Mya时的第三纪物种分化有关(Silvera et al.,2009)。而树兰亚科与兰亚科在距今64 Mya的古新世早期分化(Givnish et al.,2016),这一时期土壤干旱、二氧化碳浓度下降等剧烈气候变化频繁发生,这些因素也促进了具有CAM光合代谢途径的附生植物的生存和进化(Silvera et al.,2009)。
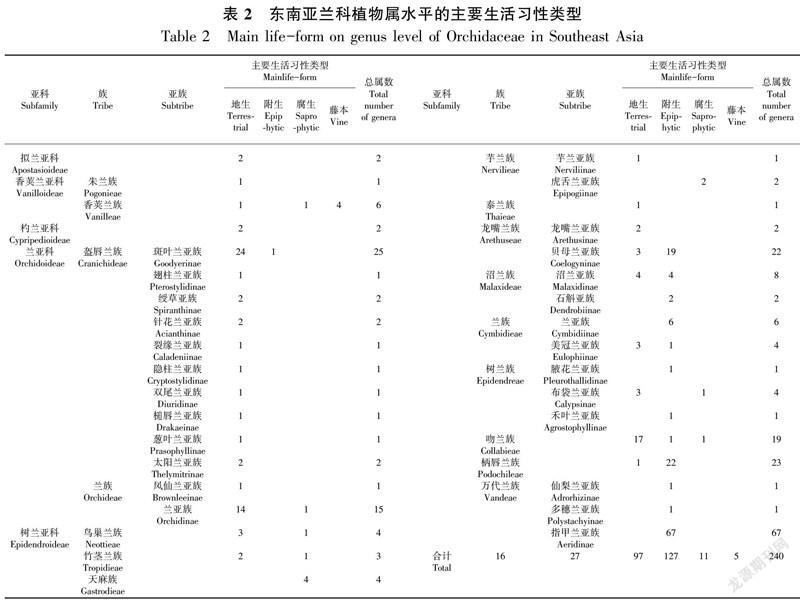
东南亚地区的兰科植物包含了附生、地生、腐生和藤本四种生活习性(表2)。
2.1 附生为主的种类
主要生活型为附生的种类有6 000种以上,约占东南亚兰科植物总数的2/3。它们分属于127属,占总属数的52.92%。附生习性为主的属主要集中在树兰亚科,种类最为丰富的类群为龙嘴兰族、柄唇兰族和万代兰族的大部分属;兰亚科仅有斑叶兰亚族Kipandiorchis 2种为附生;杓兰亚科兜兰属的部分种类具有附生习性;拟兰亚科和香荚兰亚科没有附生习性为主的属(表2)。
2.2 地生为主的种类
主要生活型为地生的种类有2 000种以上,约占东南亚兰科植物总数的1/4。它们分属于97属,占东南亚兰科植物总属数的40.42%。拟兰亚科植物均为地生;香荚兰亚科仅有朱兰族朱兰属(Pogonia)和香荚兰族芋兰属(Nervilia)为地生;兰亚科是地生习性最为主要的类群,大部分属的主要生活习性均为地生;树兰亚科在除天麻族和万代兰族的族中均具有地生为主的属,其中最为丰富的类群是吻兰族,有17个属的主要生活习性为地生(表2)。
2.3 腐生为主的种类
主要生活型为腐生的种类约有不足100种。它们分属于13属,占总属数的5.42%。主要集中在树兰亚科基部类群,如天麻族和芋兰族的虎舌兰亚族;兰亚科仅有林荫兰属(Silvorchis)、鳔唇兰属(Cystorchis)部分种类、齿唇兰属(Odontochilus)极少数种类为腐生;其余3个亚科没有腐生习性为主的属(表2)。
2.4 藤本为主的种类
主要生活型为藤本的种类有40多种,仅见于香荚兰亚科香荚兰族的倒吊兰属(Erythrorchis)、山珊瑚属(Galeola)、Pseudovanilla和香荚兰属(Vanilla)共4个属(表1),占东南亚兰科植物总属数的1.66%(表2)。
3 东南亚兰科植物的传粉系统
传粉者对兰科植物的多样性形成和分化起着重要作用(Inda et al.,2012),所以,大部分兰科植物保护中必须首要关注传粉系统及传粉者,特别是对于专一化较强的类群,如欺骗性传粉、长舌花蜂作为主要传粉者的类群(Mant et al.,2002; Swarts & Dixon,2009)。除无融合生殖,如南方玉凤花(Habenaria malintana)(Zhang & Gao,2018),自动自花授粉,如大根槽舌兰(Holcoglossum amesianum)(Liu et al.,2006)和非生物媒介辅助的自动自花授粉,如多花脆兰(Acampe rigida)(Fan et al.,2012)外,几乎所有兰科植物都是生物媒介传粉。除部分鸟类传粉类群外(Micheneau et al.,2006; Van der Niet et al.,2015),兰科植物大部分都是以昆虫作为传粉者,其中,以膜翅目(Hymenoptera)蜂类和双翅目(Diptera)蝇类传粉最为常见,占整个兰科的60%左右(Dressler,1993)。兰科植物传粉系统的特化在种间存在着极大的变异,大约60%的兰科植物拥有唯一特定的传粉者,形成了一一对应的特化传粉关系(Tremblay,1992)。
植物与传粉者间存在互惠互利的关系,传粉者为植物提供传粉服务,而植物以各种各样的报酬物回馈给传粉者,如食物、筑巢材料,甚至是提供庇护所或产卵地。在兰科植物中,报酬物大多是花蜜、花粉或脂类等食源性物质,也有树蜡、树脂类(昆虫筑巢之用)(Tremblay et al.,2005)以及芳香类物质(长舌花蜂所特有,用于交配吸引異性)(Eltz et al.,1999)。但一些植物却进化出了不为传粉者提供报酬的特质,已发现有8 000~10 000种被子植物的传粉方式是欺骗性传粉系统(Schiestl,2005; Jersáková et al.,2009),其中,兰科植物是数量最多(6 000~8 000种,占到兰科植物总数的三分之一)、欺骗形式最为多样和欺骗方式最为特别的一个类群(Jersáková et al.,2006,2009)。绝大多数的欺骗性兰科植物是通过泛化食源性欺骗(generalized food deception)方式来达到传粉的目的,而另一些则通过贝氏花拟态(batesian floral mimicry)系统,根据拟态对象的不同可分为食源性拟态(food-source mimicry)、性拟态(sexual mimicry)、栖息地拟态(shelter mimicry)、繁殖地拟态(oviposition-site mimicry)、信息素拟态(pheromone mimicry)(Jersáková et al.,2009)。
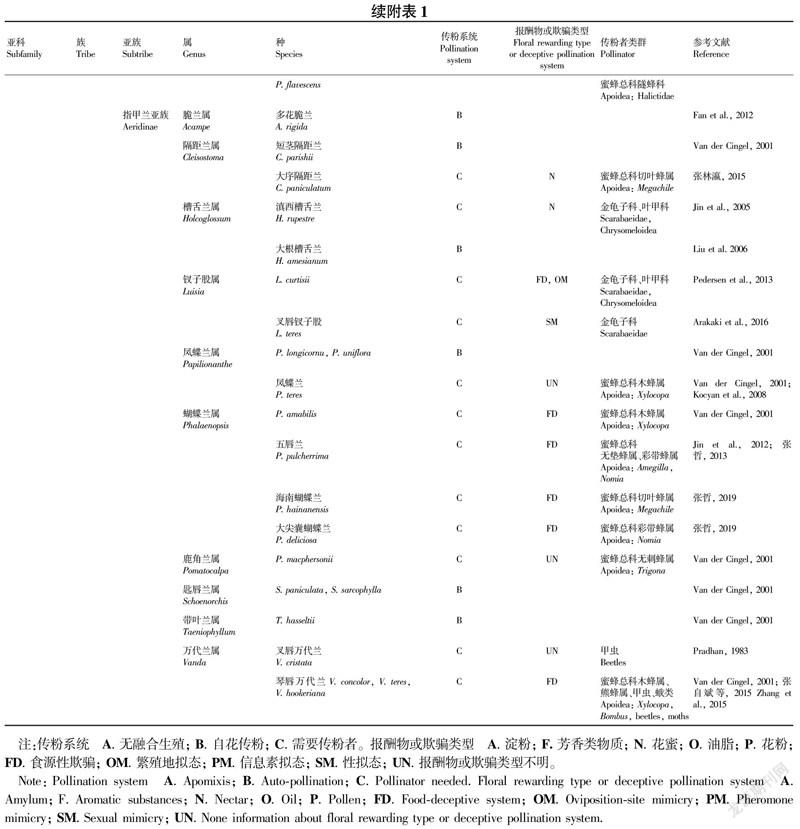
目前对东南亚兰科植物传粉生物学方面的研究还相对不足,本文仅从属水平或由其他地区报道的近缘种或近缘属来探索不同类群兰科植物的主要传粉者及其传粉系统,共整理了分布于东南亚的79个属的兰科植物传粉系统(附表1)。
3.1 自花授粉及无融合生殖系统
兰科植物的雌雄蕊集中于合蕊柱上,雌雄蕊间的隔离往往通过蕊喙来保证,进而防止自交的发生(Kurzweil et al.,2005; Efimov,2011)。一般而言,蕊喙后方凹陷形成药窝,前方形成“着粉盘”或其包囊;花粉粘合成团块,与蕊喙相连接。但在大部分自动自交的类群中,常有蕊喙退化、发育不完全或裂解的现象,进而使花粉团和柱头能够突破隔离,促使自交授粉的发生(Catling,1990)。另外,还有一些较为特别的类群,其柱头可通过分泌大量的水分促使对花粉团的包合,如Cyrtopodium polyphyllum (Catling,1990; Pansarin et al.,2008);花萼、雄蕊或花粉团的运动促使自交(Catling,1990; Liu et al.,2006);花粉团易碎或液化导致沉降于柱头表面(Hagerup,1952)等。自动自花授粉机制由祖先类群(异花授粉类群)在多种因素的影响下独立进化而来(Hapeman & Inoue,1997),现广泛出现于兰科植物多个类群中(Gamisch et al.,2014)。甚至在同属、同种内,也会出现兼性的传粉方式(既有自动自花授粉机制,又依赖传粉者)。
东南亚地区分布类群有44个属都出现了可自动自交的物种,如Thelymitra、石豆兰属(Bulbophyllum)、绶草属(Spiranthes)、头蕊兰属(Cephalanthera)、火烧兰属(Epipactis)、风兰属(Angraecum)和美冠兰属(Eulophia)(Gamisch et al.,2014)。有些物种还出现了闭花受精这种完全的自花授粉机制,如香荚兰属Vanilla bicolor(Van der Dam et al.,2010),天麻属Gastrodia flexistyloide(Suetsugu,2014)。部分自交类群还依赖非生物媒介协助,如脆兰属多花脆兰(Acampe rigida)授粉完全依赖雨水(Fan et al.,2012),通过雨滴坠落击打花药,进而反弹进入柱头完成授粉。羊耳蒜属Liparis loeselii、L. kumokiri和Oeceoclades maculata借助雨滴驱动授粉(González-Díaz & Ackerman,1988);带叶兰属Taeniophyllum hasseltii,隔距兰属Cleisostoma parishii 和凤蝶兰属Papilionanthe uniflora借助风力驱动授粉(Van der Cingel,2001)。
无融合生殖类群出现于少数物种中,如玉凤花属南方玉凤花(Habenaria malintana)(Zhang & Gao,2018),翻唇兰属白肋翻唇兰(Hetaeria cristata)(丁浩,2016)和线柱兰属线柱兰(Zeuxine strateumatica)(Sun,1997)。
3.2 具报酬物的传粉系统
尽管兰科植物以欺骗性传粉类群而闻名,但绝大多数的兰科植物还是具有报酬物。在兰科植物中,报酬物大多是花蜜、花粉或脂类等食物,也有树蜡或树脂类(昆虫筑巢之用)(Tremblay et al.,2005)以及芳香类物质(长舌花蜂所特有,用于交配吸引异性)(Eltz et al.,1999)。
由于兰科植物的花粉往往集合形成花粉团,因此,以花粉为报酬物的类群在兰科植物中并不多见,仅见于基部类群拟兰亚科,包括三蕊兰属(Neuwiedia)和拟兰属(Apostasia),其唇瓣特征进化并不明显(Kocyan & Endress,2001),花朵均没有花蜜,但以花粉作为传粉者报酬。三蕊兰属的主要传粉者为无刺蜂(Trigona bees),传粉者通过高频率的翅膀震动使花粉释放出来(Jersáková et al.,2006)以取得报酬。拟兰亚科代表着兰科植物以花粉作为报酬物的这一祖先特质(Bateman et al.,2003)。
以花蜜为报酬物的类群可能最早分化形成于香荚兰亚科,这一类群具有无蜜和花蜜两个类群,有蜜类群进一步分化形成了杓兰亚科、兰亚科及树兰亚科类群(Rudall & Bateman,2002; Kocyan et al.,2004)。以花蜜为报酬物是兰科植物中的主要报酬类型,广泛分布于兰亚科和树兰亚科,但这一特征在不同类群中出现了不断丢失或返祖事件(Cozzolino et al.,2001; Bateman et al.,2003),如Disa和Anacamptis(Johnson et al.,1998; Cozzolino et al.,2001)。以树蜡或树脂类为报酬的类群常见于Maxillaria及其近缘类群,也见于Rhetinantha notylioglossa和Heterotaxis brasiliensis等个别物种 (Whitten et al.,2007; Davies & Stpiczynska,2012),采集此种报酬的昆虫主要为蜂类,用于筑巢之用,包括胡蜂、长舌花蜂、无刺蜂和切叶蜂(Armbruster,2012,2017)。芳香类报酬仅见于长舌花蜂为传粉者的类群中,约有600种,主要分布于新热带区,包括Gongora、Notylia、Catasetum及香荚兰属等(Pansarin & Pansarin,2014; Hetheringtonrauth & Ramírez,2016)。
东南亚地区分布的兰科植物具报酬物的传粉系统类群有以下特点:拟兰亚科类群主要以花粉作为报酬物,传粉者为蜂类,如三蕊兰属Neuwiedia veratrifolia、N. zollingeri var. javanica(Kocyan & Endress,2001)。花蜜作为报酬物的类群涉及广泛,如血叶兰属(Ludisia)、绶草属(Spiranthes)、葱叶兰属(Microtis)、舌唇兰属(Platanthera)、鸟巢兰属(Neottia)等,传粉者类群也比较多样,包括蜂类、蝶类、蛾类及鸟类等(附表1)。油脂作为报酬物的类群如双袋兰属Disperis capensis,传粉者为蜂类(Steiner,1989; Johnson,1994);石豆兰属Bulbophyllum macranthum,传粉者为蝇类(Van der Cingel,2001)。以芳香类物质作为报酬物的见于香荚兰科的个别物种,传粉者类群是蜜蜂总科长舌花蜂科(Apidae:Euglossini)昆蟲(Lubinsky et al.,2006; Pansarin & Pansarin,2014)。
3.3 欺骗性传粉系统
3.3.1食源性欺骗 食源性欺骗传粉系统包括泛化的食源性欺骗和食源性拟态两种。泛化食源性欺骗传粉系统广泛存在于兰科植物中,约占欺骗性兰科植物的2/3。在此系统中,植物没有特定地模拟报酬植物,而是本身具有食源植物的花信号,以此使传粉者将其与传粉者报酬联系起来,其传粉成功依赖于传粉者较差的学习和区分能力。传粉成功且不仅取决于兰科植物自身花信号的质量,其分布区域内的其他报酬植物的丰度也对传粉成功有所影响(Chittka & Raine,2006)。食源性拟态传粉系统是指兰科植物通过模拟模型植物的花色和花型来达到欺骗的目的。蜂类的行为学实验研究发现,当蜂类获得报酬后,更倾向于访问花色和花型相似的其他个体,这就使得拟态的兰花获得更多的访问(Johnson et al.,2003)。
东南亚分布的兰科植物具有食源性欺骗传粉系统的种类有:香荚兰亚科朱兰属(Pogonia)和香荚兰属(Vanilla)的部分种类,杓兰亚科杓兰属(Cypripedium)和兜兰属(Paphiopedilum)的部分种类,兰亚科斑叶兰属(Goodyera)、双尾兰属(Diuris)、盔花兰属(Galearis)、玉凤花属(Habenaria)、舌喙兰属(Hemipilia)等的部分种类,树兰亚科头蕊兰属(Cephalanthera)、天麻属(Gastrodia)、芋兰属(Nervilia)、竹叶兰属(Arundina)、石豆兰属(Bulbophyllum)、石斛属(Dendrobium)、兰属(Cymbidium)、虾脊兰属(Calanthe)、鹤顶兰属(Phaius)及万代兰属(Vanda)等的部分种类(附表1)。
3.3.2 性拟态 性拟态是目前研究发现的最不可思议的传粉机制(Spaethe et al.,2010),它是指植物通过模拟传粉者的雌性个体来吸引雄性,雄性传粉者与花进行拟交配(Pseudocopulation)的过程中完成传粉(Schiestl,2005)。因此,性欺骗的传粉者经常是特有的,并且兰科植物模拟雌性昆虫性信息的气味只对特定的雄性传粉者具有吸引作用(Schiestl,2005)。
东南亚分布的兰科植物性拟态欺骗类群仅见于兰亚科翅柱兰属(Pterostylis)、裂缘兰属(Caladenia)、隐柱兰属(Cryptostylis)、Arthrochilus和胡须兰属(Calochilus)的部分种类,传粉者类群涉及膜翅目土蜂总科(Scolioidea)、胡蜂总科(Vespoidea)、姬蜂总科(Ichneumonoidea)和双翅目蕈蚊科(Mycetophilidae)、蚊科(Culicidae)的昆虫(附表1)。
3.3.3 繁殖地拟态 繁殖地拟态是指兰科植物利用某些昆虫的产卵行为,拟态昆虫的繁殖地信息,吸引昆虫前来产卵进而实现传粉的目的。产卵地拟态欺骗方式较为复杂,通常拟态腐败的气味或者真菌的子实体(Urru et al.,2011)。
东南亚分布的兰科植物繁殖地拟态类群见于杓兰亚科兜兰属(Paphiopedilum),兰亚科铠兰属(Corybas)和鸟足兰属(Satyrium),树兰亚科火烧兰属(Epipactis)、天麻属(Gastrodia)、石豆兰属(Bulbophyllum)、山兰属(Oreorchis)、钗子股属(Luisia)的部分种类,传粉者类群主要是双翅目蕈蚊科(Mycetophilidae)、食蚜蝇科(Syrphidae)及喜腐蝇类(附表1)。
3.3.4 信息素拟态 信息素拟态是指植物通过拟态一些与化学信号有关的信息素吸引传粉者传粉(Brodmann et al.,2009)。
东南亚分布的兰科植物已报道的信息素拟態的种类较少,如杓兰属Cypripedium fargesii,传粉者为扁足蝇科(Platypezidae)昆虫;石斛属华石斛(Dendrobium sinense)通过模拟蜜蜂报警信息素欺骗黑盾胡蜂(Vespa bicolor)传粉(附表1)。
4 结语
尽管东南亚地区兰科植还保留着极高的物种多样性,并且不断有新物种被发现,但在历史上也经历了一段人为采集和破坏的高峰。从18世纪中期到第一次世界大战前,在缅甸、婆罗洲和新几内亚等热带雨林地区,以兜兰属(Paphiopedilum)、蝴蝶兰属(Phalaenopsis)、石斛属(Dendrobium)等为代表的观赏价值极高的兰科植物遭受到毁灭性的采集和破坏(Koopowitz et al.,2003)。这些因素造成很多野生兰科植物种群逐渐退化或消失,目前全球有超过一半(约56.5%),含易危(vulnerable)、濒危(endangered)或极危(critically endangered)三类的兰科植物正面临着灭绝的危险(Gale et al.,2018)。兰科植物的生态学特化(ecological specialization)导致其生存受到一系列生物与非生物因素的影响,在生活史的各个阶段受到不同因素的制约(Cribb et al.,2003; Swarts & Dixon,2009)。兰科植物多为珍稀濒危植物,尽管所有的兰科植物都被列入了《濒危野生动植物国际贸易公约》(CITES),占该公约应保护植物在90%以上,并得到世界各国的支持和立法(Lavarack & Dixon,2003),但这也仅仅停留在科的水平上,目前没有一种兰科植物被列为IUCN保育计划的特别物种(IUCN Conservation Planning Specialist Group)(Gale et al.,2018)。东南亚地区兰科植物预计超过10 000种,约占世界兰科植物的1/3,是兰科植物最为丰富的区域之一,也是值得关注和研究的生物多样性热点区域。本文系统概述了东南亚地区兰科植物的物种多样性、生活习性及其传粉系统,尽管在诸多方面不断有新物种被发现,本文不能更加细致地进行总结概述,但对于东南亚这一兰科植物热点区域的关注以及对于兰科植物保育具有一定的理论和实践意义。
參考文献:
ANGIOSPERM PHYLOGENY GROUP,CHASE MW,CHRISTENHUSZ MJM,et al.,2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants:APG IV [J]. Bot J Linn Soc,181(1):1-20.
ARAKAKI N,YASUDA K,KANAYAMA S,et al.,2016. Attraction of males of the cupreous polished chafer Protaetia pryeri pryeri (Coleoptera:Scarabaeidae) for pollination by an epiphytic orchid Luisia teres (Asparagales:Orchidaceae) [J]. Appl Entomol Zool,51:241-246.
ARMBRUSTER WS,2012. Evolution and ecological implications of “specialized” pollinator rewards[M]// PATINY S. Evolution of plant-pollinator relationships. Cambridge,UK:Cambridge University Press:44-67.
ARMBRUSTER WS,2017. The specialization continuum in pollination systems:diversity of concepts and implications for ecology,evolution and conservation [J]. Funct Ecol,31(1):88-100.
BNZIGER H,1994. Studies on the natural pollination of three species of wild lady-slipper orchids (Paphiopedilum) in Southeast Asia [M]// PRIDGEON A. Proceedings of the 14th World Orchid Conference. Edinburgh:HMSO:201-202.
BNZIGER H,1996. The mesmerizing wart:the pollination strategy of epiphytic lady slipper orchid Paphiopedilum villosum (Lindl.) Stein (Orchidaceae) [J]. Bot J Linn Soc,121(1):59-90.
BNZIGER H,2002. Smart alecks and dumb flies:natural pollination of some wild lady slipper orchids (Paphiopedilum spp.,Orchidaceae) [M]// CLARK J,ELLIOTT WM,TINGLEY G,et al. Proceedings of the 16th World Orchid Conference. Vancouver:Vancouver Orchid Society:165-169,plates 45-57.
BNZIGER H,SUN HQ,LUO YB,2005. The pollination of a slippery lady slipper orchid in SW China:Cypripedium guttatum Swarzt (Orchidaceae) [J]. Bot J Linn Soc,148(3):251-264.
BNZIGER H,SUN HQ,LUO YB,2008. Pollination of wild lady slipper orchids Cypripedium yunnanense and C. flavum (Orchidaceae) in southwest China:why are there no hybrids? [J]. Bot J Linn Soc,156(1):51-64.
BATEMAN RM,HOLLINGSWORTH PM,PRESTON J,et al.,2003. Molecular phylogenetics and evolution of Orchidinae and selected Habenariinae (Orchidaceae) [J]. Bot J Linn Soc,142(1):1-40.
BERNHARDT P,BURNS-BALOGH P,1986. Floral mimesis in Thelymitra nuda (Orchidaceae) [J].Plant Syst Evol,151(3):187-202.
BRODMANN J,TWELE R,FRANCKE W,et al.,2009. Orchid mimics honey bee alarm pheromone in order to attract hornets for pollination [J]. Curr Biol,19(16):1368-1372.
BROOKS TM,MITTERMEIER RA,DA FONSECA GAB,et al.,2006. Global biodiversity conservation priorities [J]. Science,313(5783):58-61.
BUERKI S,FOREST F,ALVAREZ N,2014. Proto-South-East Asia as a trigger of early angiosperm diversification [J]. Bot J Linn Soc,174(3):326-333.
CATLING PM,1990. Auto-pollination in the Orchidaceae [M]// ARDITTI J. Orchid biology,reviews and perspectives,Vol. V. Portland,OR:Timber Press:121-158.
CHASE MW,CAMERON KM,FREUDENSTEIN JV,et al.,2015. An updated classification of Orchidaceae [J]. Bot J Linn Soc,177(2):151-174.
CHEN JL,ZHOU YZ,WU SS,et al.,2019 Pollination mechanism and breeding system of Pleione formosana [J]. J For Envrion,39(5):460-466. [陈进燎,周育真,吴沙沙,等,2019. 台湾独蒜兰传粉机制和繁育系统研究 [J]. 森林与环境学报,39(5):460-466. ]
CHEN XQ,LIU ZJ,ZHU GH,et al.,2009. Orchidaceae [M] WU ZY,RAVEN PH,HONG DY. Flora of China. Vol. 25 [M]. Beijing:Science Press & St. Louis:Missori Botanical Gorder Press.
CHENG J,SHI J,SHANGGUAN FZ,et al.,2009. The pollination of a self-incompatible,food-mimic orchid,Coelogyne fimbriata (Orchidaceae),by female Vespula wasps [J]. Ann Bot,104(3):565-571.
CHITTKA L,RAINE NE,2006. Recognition of flowers by pollinators [J]. Curr Opin Plant Biol,9(4):428-435.
CHRISTENSEN DE,1994. Fly pollination in the Orchidaceae [M]// ARDITTI J. Orchid biology:review and perspectives. VI. John Wiley and Sons Ltd:413-454.
CLEMENTS M,MACKENZIE A,COPSON G,et al.,2007. Biology and molecular phylogenetics of Nematoceras sulcatum,a second endemic orchid species from subantarctic Macquarie Island [J]. Polar Biol,30(7):859-869.
COZZOLINO S,ACETO S,CAPUTO P,et al.,2001. Speciation processes in Eastern Mediterranean Orchis s.l. species:molecular evidence and the role of pollination biology [J]. Isr J Plant Sci,49(2):91-103.
COZZOLINO S,WIDMER A,2005. Orchid diversity:an evolutionary consequence of deception? [J]. Trends in Ecol Evol,20(9):487-494.
CRIBB PJ,KELL SP,DIXON KW,et al. 2003. Orchid conservation:a global perspective [M]// DIXON KW,KELL SP,BARRETT RL,et al. Orchid conservation. Kota Kinabalu,Sabah:Natural History Publications:1-24.
CUN ZY,2005. A study on the reproductive ecology of Satyrium ciliatum Ldl. Orchidaceae) [D]. Beijing:University of Chinese Academy of Sciences. [寸智宇,2005. 緣毛鸟足兰的生殖生态学研究 [D]. 北京:中国科学院大学.]
DAFNI A,CALDER DM,1987. Pollination by deceit and floral mimesis in Thelymitra antennifera (Orchidaceae) [J]. Plant Syst Evol,158(1):11-22.
DAVIES KL,STPICZYNSKA M,2012. Comparative labellar anatomy of resin-secreting and putative resin-mimic species of Maxillaria s.l. (Orchidaceae:Maxillariinae) [J]. Bot J Linn Soc,170(3):405-435.
DING H,2016.Reproductive biology of Hetaeria cristata (Orchidaceae) [D]. Nanchang:Nanchang University. [丁浩,2016. 白肋翻唇蘭生殖生物学研究 [D]. 南昌:南昌大学.]
DRESSLER RL,1981. The orchids—natural history and classification [M]. Cambridge,MA:Harvard University Press.
DRESSLER RL,1993. Phylogeny and classification of the orchid family [M]. Portland:Timber Press.
EFIMOV PG,2011. An intriguing morphological variability of Platanthera s.l. [J]. Eur J Environ Sci,1(2):125-136.
ELTZ T,WHITTEN WM,ROUBIK DW,et al.,1999. Fragrance collection,storage,and accumulation by individual male orchid bees [J]. J Chem Ecol,25(1):157-176.
FAN XL,BARRETT SCH,LIN H,et al.,2012. Rain pollination provides reproductive assurance in a deceptive orchid [J]. Ann Bot,110(5):953-958.
GALE S,2007. Autogamous seed set in a critically endangered orchid in Japan:pollination studies for the conservation of Nervilia nipponica [J]. Plant Syst Evol,268(1/4):59-73.
GALE SW,FISCHER GA,CRIBB PJ,et al.,2018. Orchid conservation:Bridging the gap between science and practice[J]. Bot J Linn Soc,186(4):425-434.
GAMISCH A,FISCHER GA,COMES HP,2014. Recurrent polymorphic mating type variation in Madagascan Bulbophyllum species (Orchidaceae) exemplifies a high incidence of auto-pollination in tropical orchids [J]. Bot J Linn Soc,175(2):242-258.
GASKETT AG,2011. Orchid pollination by sexual deception:pollinator perspectives [J]. Biol Rev,86(1):33-75.
GIVNISH TJ,SPALINK D,AMES M,et al.,2015. Orchid phylogenomics and multiple drivers of their extraordinary diversification [J]. Proc R Soc B,282(1814):171-180.
GIVNISH TJ,SPALINK D,AMES M,et al.,2016. Orchid historical biogeography,diversification,Antarctica and the paradox of orchid dispersal [J]. J Biogeogr,43(10):1905-1916.
GONZLEZ-DAZ N,ACKERMAN JD,1988. Pollination,fruit set,and seed production in the orchid,Oeceoclades maculata [J]. Lindleyana,3(3):150-155.
GRANTHAM MA,FORD BA,WORLEY AC,2019. Pollination and fruit set in two rewardless slipper orchids and their hybrids (Cypripedium,Orchidaceae):large yellow flowers outperform small white flowers in the northern tall grass prairie [J]. Plant biol,21(6):997-1007.
GRAVENDEEL B,SMITHSON A,SLIK FJW,et al.,2004. Epiphytism and pollinator specialization:drivers for orchid diversity [J]. Philos Trans R Soc B,359(1450):1523-1535.
GUO YY,LUO YB,LIU ZJ,et al.,2012. Evolution and biogeography of the slipper orchids:Eocene vicariance of the conduplicate genera in the Old and New World Tropics [J]. PLoS ONE,7(6):e38788.
GUO YY,LUO YB,LIU ZJ,et al.,2015. Reticulate evolution and sea-level fluctuations together drove species diversification of slipper orchids (Paphiopedilum) in South-East Asia [J]. Mol Ecol,24(11):2838-2855.
HAGERUP O,1952. The morphology and biology of some primitive orchid flowers [J]. Phytomorphology 2:134-138.
HAPEMAN JR,INOUE K,1997. Plant-pollinator interaction and floral radiation in Platanthera (Orchidaceae) [M]// GIVNISH TJ,SYTSMA KJ. Molecular evolution and adaptive radiation. Cambridge,UK:Cambridge University Press:433-454.
HETHERINGTON-RAUTH MC,RAMREZ SR,2016. Evolution and diversity of floral scent chemistry in the euglossine bee-pollinated orchid genus Gongora [J]. Ann Bot,118(1):135-148.
HSU TC,CHUNG SW,KUO CM,2012. Supplements to the orchid flora of Taiwan (VI) [J]. Taiwania,57(3):271-277.
HSU TC,KUO CM,2010. Supplements to the orchid flora of Taiwan (IV):four additions to the genus Gastrodia [J]. Taiwania,55(3):243-248.
HSU TC,KUO CM,2011. Gastrodia albida (Orchidaceae),a new species from Taiwan [J]. Ann Bot Fenn,48(3):272-275.
HU AQ,HSU TC,LIU Y,2014. Gastrodia damingshanensis (Orchidaceae:Epidendroideae):a new myco-heterotrophic orchid from China [J]. Phytotaxa,175(5):256-262.
IKEUCHI Y,SUETSUGU K,SUMIKAWA H,2015. Diurnal skipper Pelopidas mathias (Lepidoptera:Hesperiidae) pollinates Habenaria radiata (Orchidaceae) [J]. Entomol News,125(1):7-11.
INDA LA,PIMENTEL M ,CHASE MW,2012. Phylogenetics of tribe Orchideae (Orchidaceae:Orchidoideae) based on combined DNA matrices:inferences regarding timing of diversification and evolution of pollination syndromes [J]. Ann Bot,110(1):71-90.
INOUE K,KATO M,INOUE T,1995. Pollination ecology of Dendrobium setifolium,Neuwiedia borneensis,and Lecanorchis multiflora (Orchidaceae) in Sarawak [J]. Tropics,5(1/2):95-100.
JAKUBSKA-BUSSE A,JASICKA-MISIAK I,POLIWODA A,et al.,2014. The chemical composition of the floral extract of Epipogium aphyllum Sw. (Orchidaceae):a clue for their pollination biology [J]. Arch Biol Sci,66(3):989-998.
JERSKOV J,JOHNSON SD,JRGENS A,2009. Deceptive behaviour in pants. Ⅱ. Food deception by plants:from generalized systems to specialized floral mimicry [M]// BALUKA F. Plant-environment interactions,signaling and communication in plants,from sensory plant biology to active plant behavior. Berlin Heidelberg:Springer-Verlag:223-246.
JERSKOV J,JOHNSON SD,KINDLMANN P,2006. Mechanisms and evolution of deceptive pollination in orchids [J]. Biol Rev Camb Philos Soc,81(2):219-235.
JIN XH,CHEN SC,QIN HN,2005. Pollination system of Holcoglossum rupestre (Orchidaceae):A special and unstable system [J]. Plant Syst Evol,254(1/2):31-38.
JIN XH,LI DZ,REN ZX,et al.,2012. A generalized deceptive pollination system of Doritis pulcherrima (Aeridinae:Orchidaceae) with non-reconfigured pollinaria [J]. BMC Plant Biol,12(1):67.
JIN XH,REN ZX,XU SZ,et al.,2014. The evolution of floral deception in Epipactis veratrifolia (Orchidaceae):from indirect defense to pollination [J]. BMC Plant Biol,14(1):63.
JOHNSON SD,1994. Preliminary observations on the pollination of Disperis capensis (Orchidaceae) [J]. S Afr Orchid J,25:22-23.
JOHNSON SD,1996. Bird pollination in South African species of Satyrium (Orchidaceae) [J]. Plant Syst Evol,203(1):91-98.
JOHNSON SD,ALEXANDERSSON R,LINDER HP,2003. Experimental and phylogenetic evidence for floral mimicry in a guild of fly-pollinated plants [J]. Bot J Linn Soc,80(2):289-304.
JOHNSON SD,LINDER HP,STEINER KE,1998. Phylogeny and radiation of pollination systems in Disa (Orchidaceae) [J]. Am J Bot,85(3):402-411.
JOHNSON SD,PETER CI,NILSSON LA,et al.,2003. Pollination success in a deceptive orchid is enhanced by co-occurring rewarding magnet plants[J]. Ecology,84(11):2919-2927.
JOPPA LN,ROBERTS DL,PIMM SL,2011. How many species of flowering plants are there? [J]. Proc R Soc London Ser B,278(1705):554-559.
KATO M,TSUJI K,KAWAKITA A,2006. Pollinator and stem- and corm-boring insects associated with mycoheterotrophic orchid Gastrodia elata [J]. Ann Entomol Soc Am,99(5):851-858.
KELLY MM,TOFT RJ,GASKETT AC,2013. Pollination and insect visitors to the putatively brood-site deceptive endemic spurred helmet orchid,Corybas cheesemanii [J]. New Zealand J Bot,51(3):155-167.
KOCYAN A,ENDRESS PK,2001. Floral structure and development of Apostasia and Neuwiedia (Apostastioideae) and their relationships with other Orchidaceae [J]. Int J Plant Sci,162(4):847-867.
KOCYAN A,QIU YL,ENDRESS PK,et al.,2004. A phylogenetic analysis of Apostasioideae (Orchidaceae) based on ITS,trnL-F and matK sequences [J]. Plant Syst Evol,247(3/4):203-213.
KOCYAN A,VOGEL EF,CONTI E,et al.,2008. Molecular phylogeny of Aerides (Orchidaceae) based on one nuclear and two plastid markers:a step forward in understanding the evolution of the Aeridinae [J]. Mol Phylogenet Evol,48(2):422-443.
KOOPOWITZ H,LAVARACK PS,DIXON KW,2003. The nature of threats to orchid conservation [M]// DIXON KW,KELL SP,BARRETT RL,et al. Orchid conservation. Kota Kinabalu,Sabah:Natural History Publications:25-42.
KURZWEIL H,WESTON PH,PERKINS AJ,2005. Morphological and ontogenetic studies on the gynostemium of some Australian members of Diurideae and Cranichideae (Orchidaceae) [J]. Telopea,11(1):11-33.
LI P,HUANG BQ,PEMBERTON RW,et al.,2011. Floral display influences male and female reproductive success of the deceptive orchid Phaius delavayi [J].Plant Syst Evol,296(1-2):21-27.
LI P,LUO YB,BERNHARDT P,et al.,2008a. Pollination of Cypripedium plectrochilum (Orchidaceae) by Lasioglossum spp. (Halictidae):the roles of generalist attractants versus restrictive floral architecture [J]. Plant Biol,10(2):220-230.
LI P,LUO YB,2009. Reproductive biology of an endemic orchid Cypripedium smithii in China and reproductive isolation between C. smithii and C. tibeticum [J]. Biodivers Sci,17(4):406-413. [李鵬,罗毅波,2009. 中国特有兰科植物褐花杓兰的繁殖生物学特征及其与西藏杓兰的生殖隔离研究 [J]. 生物多样性,17(4):406-413.]
LI P,LUO YB,BERNHARDT P,et al.,2006. Deceptive pollination of the Lady’s Slipper Cypripedium tibeticum (Orchidaceae) [J]. Plant Syst Evol,262(1-2):53-63.
LI P,LUO YB,DENG YX,KOU Y,2008b. Pollination of the lady’s slipper Cypripedium henryi Rolfe (Orchidaceae) [J]. Bot J Linn Soc,156(4):491-499.
LIU KW,LIU ZJ,HUANG L,et al.,2006. Pollination:self-fertilization strategy in an orchid [J]. Nature,441(7096):945-946.
LIU ZJ,LIU KW,CHEN LJ,et al.,2006. Conservation ecology of endangered species Paphiopedilum armeniacum (Orchidaceae) [J]. Acta Ecol Sin,26(9):2791-2800. [刘仲健,刘可为,陈利君,等,2006. 濒危物种杏黄兜兰的保育生态学 [J]. 生态学报,26(9):2791-2800.]
LUBINSKY P,VAN DER DAM M,VAN DER DAM A,2006. Pollination of Vanilla and evolution in Orchidaceae [J]. Lindleyana,75 (12):926-929.
LUO YB,CHEN SC,2010. Observations of putative pollinators of Hemipilia flabellata Bur. et Franch. (Orchidaceae) in north-west Yunnan Province,China [J]. Bot J Linn Soc,131(1):45-64.
LTTGE U,2004. Ecophysiology of crassulacean acid metabolism (CAM) [J]. Ann Bot,93(6):629-652
MA XK,SHI J,BNZIGER H,et al.,2016. The functional significance of complex floral colour pattern in a food-deceptive orchid [J]. Funct Ecol,30(5):721-732.
MANT JG,SCHIESTL FP,PEAKALL R,et al.,2002. A phylogenetic study of pollinator conservatism among sexually deceptive orchids [J]. Evolution,56(5):888-898
MARTOS F,CARIOU ML,PAILLER T,et al.,2015. Chemical and morphological filters in a specialized floral mimicry system [J]. New Phytol,207(1):225-234.
MATSUI K,USHIMARU T,FUJITA N,2001. Pollinator limitation in a deceptive orchid,Pogonia japonica,on a floating peat mat [J]. Plant Species Biol,16(3):231-235.
MICHENEAU C,FOURNEL J,PAILLER T,2006. Bird pollination in an angraecoid orchid on Reunion Island (Mascarene Archipelago,Indian Ocean) [J]. Ann Bot,97(6):965-974.
OKADA H,KUBO S,MORI Y,1996. Pollination system of Neuwiedia veratrifolia Blume (Orchidaceae,Apostasioideae) in the Malesian wet tropics [J]. APG,47(2):173- 181.
PANSARIN ER,AGUIAR JMRVB,PANSARIN LM,2014. Floral biology and histochemical analysis of Vanilla edwallii Hoehne (Orchidaceae:Vanilloideae):an orchid pollinated by Epicharis (Apidae:Centridini) [J]. Plant Species Biol,29(3):242-252.
PANSARIN ER,PANSARIN LM,2014. Floral biology of two Vanilloideae (Orchidaceae) primarily adapted to pollination by euglossine bees [J]. Plant Biol,16(6):1104-1113.
PANSARIN LM,PANSARIN ER,SAZIMA M,2008. Reproductive biology of Cyrtopodium polyphyllum (Orchidaceae):a Cyrtopodiinae pollinated by deceit [J]. Plant Biol,10(5):650-659.
PATT JM,MERCHANT MW,WILLIAMS DRE,1989. Pollination biology of Platanthera stricta (Orchidaceae) in Olympic National Park,Washington [J]. Am J Bot,76:1097-1106.
PEAKALL R,BEATTIE AJ,1989. Pollination of the orchid Microtis parviflora R. Br. by flightless worker Ants [J]. Funct Ecol,3(5):515-522.
PEDERSEN H,WATTHANA S,SRIMUANG K,2013. Orchids in the torrent:on the circumscription,conservation and rheophytic habit of Epipactis flava [J]. Bot J Linn Soc,172(3):358-370.
PEDRON M,BUZATTO CR,SINGER RB,et al.,2012. Pollination biology of four sympatric species of Habenaria (Orchidaceae:Orchidinae) from southern Brazil [J]. Bot J Linn Soc,170(2):141-156.
PETER CI,JOHNSON SD,2009. Autonomous self-pollination and pseudo-fruit set in South African species of Eulophia (Orchidaceae)[J]. S Afr J Bot,75(4):791-797.
PETTERSSON B,1989. Pollination in the African species of Nervilia (Orchidaceae) [J]. Lindleyana,4:33-41.
PRADHAN GM,1983. Vanda cristata[J]. America Orchid Soc Bull,52(5):464-468
PRIDGEON AM,BATEMAN RM,COX AV,et al.,1997. Phylogenetics of subtribe Orchidinae (Orchidoideae,Orchidaceae) based on nuclear ITS sequenes,1. intergeneric relationships and polyphyly of Orchis sensu lato [J]. Lindleyana,12(2):89-109.
PRIDGEON AM,CRIBB PJ,CHASE MW,et al.,1999. Genera Orchidacearum Vol. 1,General Introduction,Apostasioideae & Cypripedioideae [M]. Oxford:Oxford University Press.
PRIDGEON AM,CRIBB P J,CHASE MW,et al.,2001. Genera Orchidacearum Vol. 2,Orchidoideae,Part 1 [M]. Oxford:Oxford University Press.
PRIDGEON AM,CRIBB P J,CHASE MW,et al.,2003. Genera Orchidacearum Vol. 3,Orchidoideae,Part 2 [M]. Oxford:Oxford University Press.
PRIDGEON AM,CRIBB PJ,CHASE MW,et al.,2005. Genera Orchidacearum Vol. 4,Epidendroideae (Part 1) [M]. Oxford:Oxford University Press.
PRIDGEON AM,CRIBB PJ,CHASE MW,et al.,2009. Genera Orchidacearum Vol. 5,Epidendroideae (Part 2) [M]. Oxford:Oxford University Press.
PRIDGEON AM,CRIBB PJ,CHASE MW,et al.,2014. Genera Orchidacearum. Vol. 6,Epidendroideae (Part 3) [M]. Oxford:Oxford University Press.
RAMREZ SR,ELTZ T,FUJIWARA MK,et al.,2011. Asynchronous diversification in a specialized plant-pollinator mutualism [J]. Science,333(6050):1742-1746.
REN ZX ,LI DZ ,BERNHARDT P,et al.,2011. Flowers of Cypripedium fargesii (Orchidaceae) fool flat-footed flies (Platypezidae) by faking fungus-infected foliage [J]. Proc Natl Acad Sci USA,108(18):7478-7480.
REN ZX,WANG H,BERNHARDT P,et al.,2014. Which food-mimic floral traits and environmental factors influence fecundity in a rare orchid,Calanthe yaoshanensis? [J]. Bot J Linn Soc,176(3):421-433.
RICKLEFS RE,RENNER SS,1994. Species richness within families of flowering plants [J]. Evolution,48(5):1619-1636.
RUDALL PJ,BATEMAN RM,2002. Roles of synorganisation,zygomorphy and heterotopy in floral evolution:the gynostemium and labellum of orchids and other lilioid monocots [J]. Biol Rev Camb Philos Soc,77(3):403-441.
SCHIESTL FP,2005. On the success of a swindle:pollination by deception in orchids [J]. Naturwissenschaften,92(6):255-264.
SCHIESTL FP,COZZOLINO S,2008. Evolution of sexual mimicry in the orchid subtribe orchidinae:the role of preadaptations in the attraction of male bees as pollinators [J]. BMC Evol Biol,8(1):27
SCHIESTL FP,SCHLTER PM,2009. Floral isolation,specialized pollination,and pollinator behavior in orchids [J]. Ann Rev Entomol,54(1):425-446.
SHI J,LUO YB,BERNHARDT P,et al.,2009. Pollination by deceit in Paphiopedilum barbigerum (Orchidaceae):a staminode exploits the innate colour preferences of hoverflies (Syrphidae) [J]. Plant Biol,11(1):17-28.
SILVERA K,SANTIAGO LS,CUSHMAN JC,et al.,2009. Crassulacean acid metabolism and epiphytism linked to adaptive radiations in Orchidaceae [J]. Plant Physiol,149(4):1838-1847.
SINGER RB,SAZIMA M,1999. The pollination mechanism in the ‘Pelexia alliance’ (Orchidaceae:Spiranthinae) [J]. Bot J Linn Soc,1999,131(3):249-262.
SOSA V,CAMERON KM,ANGULO DF,et al.,2016. Life form evolution in epidendroid orchids:Ecological consequences of the shift from epiphytism to terrestrial habit in Hexalectris [J]. Taxon,65(2):235-248.
SPAETHE J,STREINZER M,PAULUS HF,2010. Why sexually deceptive orchids have colored flowers [J]. Commun Integr Biol,3(2):139-141.
SRIMUANG K,WATTHANA S,PEDERSEN H,et al.,2010. Aspects of biosubsistence in Sirindhornia (Orchidaceae):Are the narrow endemics more reproductively restricted than their widespread relative? [J]. Ann Bot Fenn,47(6):449-459.
ST GEORGE I,2007. The pollination of Nematoceras iridescens [J]. J N Z Nat Orchid Gro,102:10.
STEINER KE,1989. The pollination of Disperis (Orchidaceae) by oil-collecting bees in southern Africa [J]. Lindleyana,4:164-183.
SUETSUGU K,2013. Autogamous fruit set in a mycoheterotrophic orchid Cyrtosia septentrionalis [J]. Plant Syst Evol,299:481-486.
SUETSUGU K,2014. Gastrodia flexistyloides (Orchidaceae),a new mycoheterotrophic plant with complete cleistogamy from Japan [J]. Phytotaxa,175(5):270-274.
SUETSUGU K,2015. Autonomous self-pollination and insect visitors in partially and fully mycoheterotrophic species of Cymbidium (Orchidaceae) [J]. J Plant Res,128(1):115-125.
SUETSUGU K,FUKUSHIMA S,2014a. Pollination biology of the endangered orchid Cypripedium japonicum in a fragmented forest of Japan [J]. Plant Species Biol,29(3):294-299.
SUETSUGU K,FUKUSHIMA S,2014b. Bee pollination of the endangered orchid Calanthe discolor through a generalized food-deceptive system [J]. Plant Syst Evol,300(3):453-459.
SUETSUGU K,NAITO R,FUKUSHIMA S,2015. Pollination system and the effect of inflorescence size on fruit set in the deceptive orchid Cephalanthera falcata [J]. J Plant Res,128(4):585-594.
SUETSUGU K,TANAKA K,2014. Consumption of Habenaria sagittifera pollinia by juveniles of the katydid Ducetia japonica[J]. Entomol Sci,17(1):122-124.
SUGIURA N,1995. The pollination ecology of Bletilla striata (Orchidaceae) [J]. Ecol Res,10(2):171-177.
SUGIURA N,2013. Specialized pollination by carpenter bees in Calanthe striata (Orchidaceae),with a review of carpenter bee pollination in orchids [J]. Bot J Linn Soc,171(4):730-743.
SUGIURA N,2016. Mate-seeking and oviposition behavior of Chyliza vittata (Diptera:Psilidae) infesting the leafless orchid Gastrodia elata [J]. Entomol Sci,19(2):129-132.
SUGIURA N,FUJIE T,INOUE K,et al.,2001. Flowering phenology,pollination,and fruit set of Cypripedium macranthos var. rebunense,a threatened lady’s slipper (Orchidaceae) [J]. J Plant Res,114(2):171-178.
SUGIURA N,GOUBARA M,KITAMURA K,et al.,2002. Bumblebee pollination of Cypripedium macranthos var. rebunense (Orchidaceae):a possible case of floral mimicry of Pedicularis schistostegia (Orobanchiaceae) [J]. Plant Syst Evol,235(1-4):189-195.
SUGIURA N,OKAJIMA Y,MAETA Y,1997. A note on the pollination of Oreorchis patens (Orchidaceae) [J]. Ann Tsukuba Bot Gard,16:69-74.
SUN HQ,HUANG BQ,YU XH,et al.,2011. Reproductive isolation and pollination success of rewarding Galearis diantha and non-rewarding Ponerorchis chusua (Orchidaceae) [J]. Ann Bot,107(1):39-47.
SUN M,1997. Genetic Diversity in three colonizing orchids with contrasting mating systems[J]. Am J Bot,84(2):224-232.
SWARTS ND,BATTY A,HOPPER S,et al. 2007. Does integrated conservation of terrestrial orchids work? [J]. Lankesteriana,7(1-2):219-222.
SWARTS ND,DIXON KW,2009. Terrestrial orchid conservation in the age of extinction [J]. Ann Bot,104(3):543-556.
S'WICZKOWSKA E,KOWALKOWSKA AK,2015. Floral nectary anatomy and ultrastructure in mycoheterotrophic plant,Epipogium aphyllum Sw. (Orchidaceae) [J]. Sci World J,2015:1-11.
TAAAJ I,OSTROWIECKA B,WOSTOWSKA E,et al.,2017. The ability of spontaneous autogamy in four orchid species:Cephalanthera rubra,Neottia ovata,Gymnadenia conopsea,and Platanthera bifolia [J]. Acta Biol Crac Ser Bot,59(2):51-61.
TAN K,MALABRIGO PL,REN MX,2020. Origin and evolution of biodiversity hotspots in Southeast Asia [J]. Acta Ecol Sin,2020,40(11):3866-3877. [譚珂,Malabrigo PL,任明迅,2019. 东南亚生物多样性热点地区的形成与演化 [J]. 生态学报,40(11):3866-3877.]
THOMAS DC,HUGHES M,PHUTTHAI T,et al.,2012. West to east dispersal and subsequent rapid diversification of the mega-diverse Begonia (Begoniaceae) in the Malesian archipelago [J]. J Biogeogr,39(1):98-113.
TIFFNEY BH,MAZE SJ,1995. Angiosperm growth habit,dispersal and diversification reconsidered [J]. Ecol Evol,9(1):93-117.
TREMBLAY RL,1992. Trends in the pollination ecology of the Orchidaceae:evolution and systematics [J]. Can J Bot,70(3):642-650.
TREMBLAY RL,ACKERMAN JD,ZIMMERMAN JK,et al.,2005. Variation in sexual reproduction in orchids and its evolutionary consequences:a spasmodic journey to diversification [J]. Bot J Linn Soc,84(1):1-54.
URRU I,STENSMYR MC,HANSSON BS,2011. Pollination by brood-site deception[J]. Phytochemistry,72(13):1655-1666.
VAN DER CINGEL NA,2001. An atlas of orchid pollination:America,Africa,Asia and Australia [M]. Rotterdam:Balkema.
VAN DER DAM AR,HOUSEHOLDER JE,LUBINSKY P,2010. Vanilla bicolor Lindl. (Orchidaceae) from the Peruvian Amazon:auto-fertilization in Vanilla and notes on floral phenology [J]. Genet Resour Crop Evol,57(4):473-480.
VAN DER NIET T,COZIEN RJ,JOHNSON SD,2015. Experimental evidence for specialized bird pollination in the endangered South African orchid Satyrium rhodanthum and analysis of associated floral traits [J]. Bot J Linn Soc,177(1):141-150.
VAN DER NIET T,HANSEN DM,JOHNSON SD,2011. Carrion mimicry in a South African orchid:flowers attract a narrow subset of the fly assemblage on animal carcasses [J]. Ann Bot,107(6):981-992.
VAN DER PIJL,DODSON CH,1966. Orchid flowers:their pollination and evolution [M]. Coral Gables,USA:University of Miami Press.
WALLACE LE,2006. Spatial genetic structure and frequency of interspecific hybridization in Platanthera aquilonis and P. dilatata (Orchidaceae) occurring in sympatry [J]. Am J Bot,93(7):1001-1009.
WANG CQ,LUO YB,TAI YD,et al.,2008. Ants pollinate Neottia listeroides (Orchidaceae) in Sichuan,China [J]. J Syst Evol,46(6):836-846. [王淳秋,罗毅波,台永东,等,2008. 蚂蚁在高山鸟巢兰中的传粉作用 [J]. 植物分类学报,46(6):836-846.]
WANG W,2013. Pollination biology of Calanthe alismaefolia [D]. Nanchang:Nanchang University. [王武,2013. 泽泻虾脊兰的传粉生物学研究 [D]. 南昌:南昌大学.]
WESTEBERHARD K,NEUBIG KM,WHITTEN WM,et al.,2010. Evolution along the crassulacean acid metabolism continuum [J]. Funct Plant Biol,37(11):995-1010.
WESTEBERHARD MJ,SMITH JAC,WINTER K,2011. Photosynthesis,reorganized [J]. Science,332(6027):311-312.
WHITTEN WM,WILLIAMS NH,BLANCO MA,et al.,2007. Molecular phylogenetics of Maxillaria and related genera (Orchidaceae:Cymbidieae) based upon combined molecular data sets [J]. Am J Bot,94(11):1860-1889.
WILLEMS JH,LAHTINEN ML,1997. Impact of pollination and resource limitation on seed production in a border population of Spiranthes spiralis (Orchidaceae) [J]. Plant Biol,46(4):365-375.
XIONG YZ,LIU CQ,HUANG SQ,2015. Mast fruiting in a hawkmoth-pollinated orchid Habenaria glaucifolia:an 8-year survey [J]. J Plant Ecol,8(2):136-141.
YANG XQ,2007. Pollination biology of Ponerorchis chusua and two Galearis species[D]. Beijing:University of Chinese Academy of Sciences. [楊小琴,2007. 广布小蝶兰及两种根茎兰属植物的传粉生物学研究 [D]. 北京:中国科学院大学.]
ZHA ZB,TANG J,LIANG YL,et al.,2016. Breeding system and pollination biology of Goodyera foliosa (Orchidaceae) [J]. J Trop Subtrop Bot,24(3):333-341. [查兆兵,唐静,梁跃龙,等,2016. 多叶斑叶兰繁育系统与传粉生物学研究[J]. 热带亚热带植物学报,24(3):333-341.]
ZHANG HF,LI LQ,LIU ZJ,et al.,2010. The butterfly Pieris rapae resulting in the reproductive success of two transplanted orchids in a botanical garden [J]. Biodivers Sci,18(1):11-18.[张洪芳,李利强,刘仲健,等,2010. 菜粉蝶对两种迁地保护的兰科植物传粉和繁殖成功的作用 [J]. 生物多样性,18(1):11-18.]
ZHANG LY,2015. Pollination biology and germination characteristics of Cleisostoma paniculatum [D]. Fuzhou:Fujian Agriculture and Forestry University. [张林瀛,2015. 大序隔距兰(Cleisostoma paniculatum)传粉生物学及种子萌发特性研究 [D]. 福州:福建农林大学.]
ZHANG WL,GAO JY,2018. High fruit sets in a rewardless orchid:a case study of obligate agamospermy in Habenaria [J]. Aust J Bot,66(2):144-151.
ZHANG Z,2013. Reproductive ecology of Phalaenopsis pulcherrima(Orchidaceae),a species endemic to East Asia [D]. Haikou:Hainan University [張哲,2013. 东亚特有种五唇兰繁殖生态学研究 [D]. 海口:海南大学.]
ZHANG Z,2019.Conservation biology of three Phalaenopsis species in Hainan Island [D]. Haikou:Hainan University. [张哲,2019. 海南三种蝴蝶兰属植物的保育生物学研究 [D]. 海口:海南大学.]
ZHANG ZB,CHENG J,YANG M,et al.,2015. Food-deceptive pollination of Vanda concolor (Orchidaceae) [J]. J Beijing For Univ,37(6):100-106. [张自斌,程瑾,杨媚,等,2015. 琴唇万代兰食源性欺骗传粉研究 [J]. 北京林业大学学报,37(6):100-106.]
ZHOU T,JIN XH,2018. Molecular systematics and the evolution of mycoheterotrophy of tribe Neottieae (Orchidaceae,Epidendroideae) [J]. Phytokeys,94(94):39-49.
ZHOU X,LIN H,FAN XL,et al.,2012. Autonomous self-pollination and insect visitation in a saprophytic orchid,Epipogium roseum (D. Don) Lindl [J]. Aus J Bot,60(2):154.
ZHOU XX,2017.Taxonomy and phylogeography study of Sect. Goodyera (Orchidaceae) [D]. Shanghai:East China Normal University. [周晓旭,2017. 斑叶兰组Sect. Goodyera(兰科)分类学与谱系地理学研究 [D]. 上海:华东师范大学.]
(责任编辑 李 莉)
