Morpho-anatomical characterization of Rhododendron subsessile Rendle, an endangered species of the Cordillera Central Range,Philippines
2021-01-11MabelGraceBitayanStarleeCervantesJonesNapaldet
Mabel Grace V. Bitayan · Starleeñe S. Cervantes ·Jones T. Napaldet
Abstract Conservation of endangered plant species requires a prerequisite basic knowledge of their taxonomy and ecology. Unfortunately in the Cordillera Central Range,several species have not been studied, even though this area harbors a unique type of vegetation not found in other parts of the Philippines. This study documents the morpho-anatomical characteristics of Rhododendron subsessile Rendle, one of the unique and endangered flora in the area.The species exhibits a morphology typical of plants in high elevations, being a hardy shrub with small tough leaves, and showy pink flowers that rival those of commercial rhododendron species. New findings on the species morphology include a variable number of corolla lobes, stamens and a description of its fruits and seeds. The anatomical features of the species are reported here for the first time. The plant exhibits a typical dicot anatomy with dorsiventral leaves,woody stem and root system with the usual arrangement of periderm followed by secondary phloem, vascular cambium, secondary xylem with vessel elements and pith (in the case of the stem). A unique characteristic was identified as the biseriate upper epidermis of the leaf. Documenting the morphological and anatomical features of the species should prove valuable as input baseline information in formulating measures for its conservation.
Keywords Taxonomy · Biodiversity conservation ·Biseriate epidermis · Variable stamens
Introduction
Rhododendron, the largest genus of the family Ericaceae of about 850 species (Zhang 2003), are important species widely distributed in temperate regions of Asia, North America, and Europe, as well as in tropical regions of southeast Asia and northern Australia. The largest number of species is native to Asia where they occur in a variety of habitats and exhibit considerable diversity in size and shape(Wang et al. 2006). Species of this genus usually grow in areas of high rainfall and high humidity on acidic soils, conditions under which few plants would survive (Gibbs et al.2011). Plant life-forms range from small, prostrate, alpine shrubs 5-cm tall to trees with large leaves that reach heights of 24 m (Leach 1961).
Rhododendron taxa are under threat of extinction in the wild as a result of habitat clearance and the increasing impact of climate change. Action clearly must be taken to conserve these threatened species as a matter of urgency(Gibbs et al. 2011). In southeast Asia, Rhododendron spp.occur primarily in tropical coniferous forests, a biome identified by Murphy and Romanuk (2016) as one of the least represented in local-scale species richness and distribution studies, while at the same time, one of most susceptible to land-use changes. These identify not only the need to implement conservation measures but also for basic studies on these species to serve as a foundation for conservation measures. This is supported by the findings of the Secretariat of the Convention on Biological Diversity (2018) which identified an insufficient knowledge base as one of the main barriers for biodiversity conservation, together with i nsuffi cient mainstreaming of biodiversity considerations in economic sectors and in cross-sectoral policies such as development planning and budgets. This highlights the need for more biodiversity studies and more efficient dissemination of biodiversity data to several sectors, particularly to those who need them.
Since Rhododendron was categorized by the Swedish botanist, Carl Linnaeus, its infrageneric relationships of morphological characteristics and molecular data have been well- studied by numerous taxonomists (Wang et al. 2006).There also have been studies on wood anatomy (Rivett 1920;Merev and Yavuz 2000), leaf epidermis (Wang et al. 2006),and as well as descriptions of new species (Kores 1984).However, as such a diverse and widely distributed genus,most studies are focused on specific regions or countries,and many species are not thoroughly studied, aside from their initial discovery and description. Such is the case of Rhododendron subsessile Rendle, an endemic shrub of the Cordillera Central Range (CCR) in northern Philippines. It was listed by Fernando et al. (2008) as one of the endangered plant species in the country but without any research data on this plant. Volis (2015) argued the need for thoroughly documenting endangered species, their description, distribution and population dynamics, to precede any conservation measures. To address this, the morphological and anatomical features of R. subsessile were characterized in this study. By this documentation, it is a contribution to baseline information about the species, which in turn, will help craft measures for its conservation.
Materials and methods
Study site
Samples of R. subsessile were collected from the village of Barangay Natubleng in the municipality of Buguias,Benguet (Fig. 1). Three sites were established. The area is an emerging tourist spot in Benguet for agrotourism, and is one of the many upland communities where the original mossy forest was heavily transformed into conventional vegetable farms. The only remnants of the original flora are found in steep ravines, unsuitable as farm land, where R. subsessile is found. Pelser et al. (2011) documented the occurrence of this species in different parts of the Cordillera Central Range stretching from Mountain Province to Ifugao to Benguet.
Morphological characterization and morphometrics
In the field, plant height was measured before collecting leaf and flower samples from 21 individuals. Samples were brought to the Taxonomy Laboratory of the Biology Department, Benguet State University and were characterized phytographically. This means coming up of a complete morphological description of the plant and its organs-such as roots, stems, leaves, flowers/inflorescence, fruits and seeds,using taxonomic references (Pancho and Gruezo 1983, 2006,2009, 2012; Singh 1999). Aside from these, the plant organs or parts were measured as part of the description. These measurements were subjected to descriptive statistics to determine mean, range and standard deviation.
Anatomical description
After the morphological characterization, plant sections of the leaf, stem and roots were prepared and subjected to light microscopy to document anatomical features. References such as Haupt (1953), Fahn (1982), Bell (1991, 2008) and Shipunov (2018) were followed to describe the anatomy of the plant.
Results
Morphological description and morphometrics
Plant habit
Rhododendron subsessile is a suffrutescent shrub with a woody base that does not die down each year, perennial and evergreen growing up to 2.2 m but the average height of the sampled population in this study was 1.5 m (Table 1;Fig. 2 a). Shorter individuals generally occurred on exposed ridges where all the vegetation was short, but taller ones were found in shaded areas on deep soils interspersed with other Rhododendron spp., Vaccinium spp., Gaultheria spp.,Syzygium spp., and Tasmannia piperita.
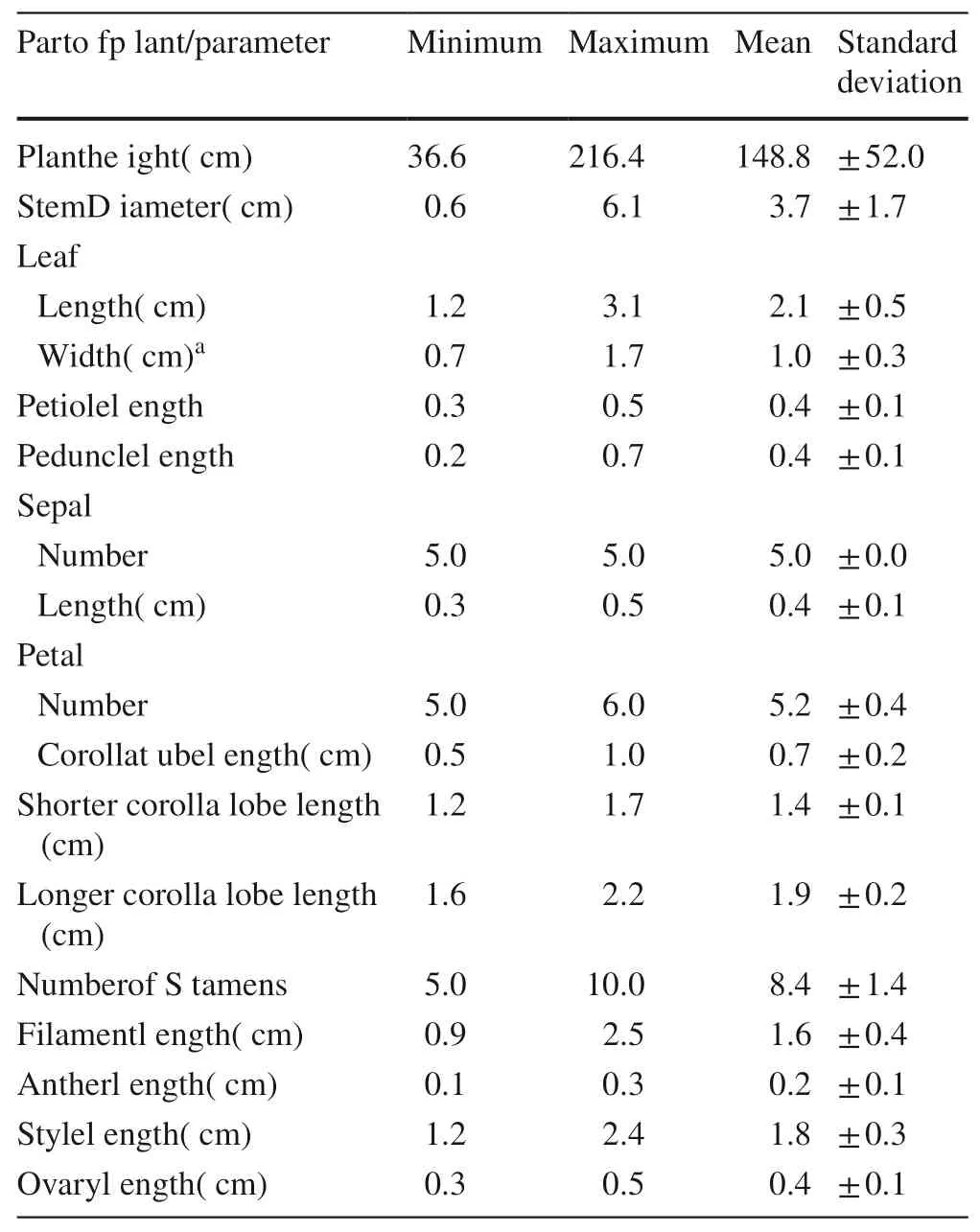
Table 1 Biometric characters of Rhododendron subsessile

Fig. 2 Morphological features of Rhododendron subsessile (a habit;b close up of the leaf; c the flowering shoots; d close-up of the flower showing 5 and 6 petal lobes)
Stem
The stems are light brown and often associated with lichens or bryophytes. More than two stems or trunks (usually 3-6)rise from the base with one or two being significantly larger.These major stems were measured 10 cm above the ground where there was profuse branching.
Leaves
The leaves are small, 1.2-3.1 cm long × 0.7-1.7 cm wide,simple, alternate to spiral with no particular pattern, ovate to elliptic to oblong, with short petioles (0.3-0.5 cm), almost sessile-hence the epithet, subsessile (Fig. 2 b).
Young leaves and shoots are heavily covered with erect white to brownish hairs but mature leaves are glabrous.
Flowers
The inflorescences consist of 2-4 regular, complete flowers with short peduncles (0.2-0.7 cm); five green, ovate, hairy sepals fused at the base; a pink bilabiate corolla with 5-6 distinct lobes fused near the base; a variable number (5-10)of filamentous stamens with dithecal and dorsifixed anthers;and, a superior ovary of five syncarpous carpels covered with white erect hairs up to the middle of the style and with a discoid stigma (Fig. 2 c, d).
Fruit and seeds
The fruit is silique with persistent sepals, axile placentation, brown, dehiscing into five equal portions giving a starlike appearance with a central dark brown pointed replum.The seeds are minute achene, dark brown to black, and cone-shaped.
Anatomical characterization of R. subsessile
The leaf
On cross section, the leaf shows both upper and lower epidermises enclosing a dorsiventral mesophyll. The upper epidermis is distinctly biseriate composed oflarge, transparent,cuboidal cells covered by thick cuticle (Fig. 3 c). The lower epidermis is uniseriate. The mesophyll is distinctly divided into palisade and spongy layers. The palisade layer consists of elongated, tightly packed cells and is the major photosynthetic site in the leaf. In contrast, the spongy layer is more spread out with air pockets and is composed of cuboidal cells. Clusters of single-celled, elongated trichomes were also observed in the lower epidermis (Fig. 3 d). The midrib is conspicuous with distinct xylem in the upper portion and phloem under it. The vascular bundle of the midrib is covered by large parenchyma cells with a thicker layer at the lower portion, making the lower midrib more rounded.Collenchyma layers were observed near the lower epidermis.The cross-section of the petiole (Fig. 3 e) shows a unilacunar vascular bundle covered by several layers of parenchyma cells and a few collenchyma layers on the underside. The upper side of the petiole is relatively flat while the lower side is rounded. The stomata are anomocytic or irregular type,whereas the subsidiary cells are similar to the remaining epidermal cells (Metcalf and Chalk 1950).
The stem
The stem is woody up to the young shoots and we were unable to obtain a cross section of the primary tissues.Nevertheless, a cross section of the secondary tissues was obtained. The periderm forms the outermost covering of the stem and is visibly multi-layered but the phellem, phellogen and phelloderm are indistinct (Fig. 4 a). Under the periderm,the secondary vascular tissues are in this order: secondary phloem, vascular cambium, and secondary xylem. The primary xylem is distinct and is located below the secondary xylem, enclosing the central parenchymatous pith. In the secondary xylem, xylem rays are prominent, appearing as black lines, while the vessel elements are distinctly larger.
The root
As with the stem, we were unable to obtain a cross section of young roots to show its primary tissues. However, we obtained cross sections of older roots. The periderm is the outermost covering, visibly multi-layered but not evenly distributed (Fig. 4 b). Some areas of the periderm have thicker layers compared to other portions. Below the periderm is a thin layer of secondary phloem followed by vascular cambium and a thick layer of secondary xylem up to the central core. The older woody roots have distinct xylem rays and large vessel elements but no remnants of a pith.
Discussion
Morphological description and morphometrics
The description and measurements conform with the original description by Rendle (1896) with some variations. The leaves in this study are relatively shorter (1.2-3.1 vs 2-4 cm)but wider (0.7-1.7 vs 0.3-0.6) than the original description.These differences could be attributed to the leaf’s plasticity which could be significantly affected by environmental factors (Pancho and Gruezo 1983; Buot and Okitsu 1999).However, in terms of floral characters, the measurements in this study are similar to the original description. Thissupport previous findings that floral characters are generally not plastic (Pancho and Gruezo 1983).

Fig. 3 Anatomical features of a Rhododendron subsessile leaf showing a typical dicot anatomy but a unique biseriate upper epidermis (a upper epidermis at high power objective (HPO),b lower epidermis at HPO showing stomata, c lamina x.s. at low power objectives(LPO), d lower epidermis at HPO showing trichomes (T),e cross-section of the petiole at LPO; S-Stomata, UE-upper epidermis, LE-lower epidermis, PL-palisade, SP-spongy, Stom-Stomata, M-midrib, X-xylem, P-phloem,VB-Vascular bundle)
Rhododendron subsessile contrasts with other species of rhododendrons that have different spring and summer leaves,usually temperate species (Chamberlain and Rae 1990). The variable number of stamens in R. subsessile is first reported here, and this is not simply be attributed to stamens dropping since the flowers were just blooming or at the onset of anthesis. This study also includes the first description of the fruits and seeds of R. subsessile.
Anatomical description of R. subsessile
The anatomical characters show a typical dicot anatomy with dorsiventral leaves, woody stem and roots with the usual arrangement of periderm followed by secondary phloem,vascular cambium, secondary xylem with vessel elements,and pith (in the case of the stem). However, a unique feature was identified in the biseriate upper epidermis of the leaf.The epidermis ofleaves is usually uniseriate (Prateeksha n.d.; Ayomipo and Johnson 2015; Shipunov 2018), and only some species of Basella, Ficus, Piper and Nerium have multiseriate epidermis. Now, as a result of this study, R. subsessile appears to be part of this group. It is believed that a multiple-layered epidermis protects the inner mesophyll from desiccation. This is readily understood in the case of R.subsessile which grows on exposed ridges with a stony substrate, high precipitation and winds. However, we hypothesize that the desiccation protection of the biseriate epidermis in R. subsessile is functional only during summer drought months, but is more applicable in providing toughness of the leaf against windy conditions and heavy rainfalls.

The anomocytic or irregularly celled stomata of R.subsessile is similar to those of R. redowskianum Maxim.(Wang et al. 2006). Anomocytic stomata have also been reported in species of Lilium and for species in Boraginaceae, Ranunculaceae and Geraniaceae families (Napaldet 2017). On the other hand, the stem anatomy of R. subsessile shows distinct growth rings with evenly distributed pores without a specific pattern. This is consistent with the general wood description of rhododendrons by Merev and Yavuz (2000).
From a conservation perspective, it is important to understand the many aspects of plant biology, including the ecological and physiological. An appreciation of both morphology and anatomy is fundamental. Plant anatomy,along with “whole plant” physiology and comparative plant morphology, has long formed the core of general botany courses. This is the subject matter at the center of the study of plants at the organism level. Aside from being a source of characters for taxonomic classification and phylogeny reconstruction, plant morphology and anatomy elucidate the relationship of these structures to their functions or physiology (Pancho and Gruezo 2012).This is true in the case of R. subsessile. As an endangered species, it needs thorough characterization and documentation for baseline information for conservation measures(Volis 2015).
Conclusions
This study fully characterized the morphology and anatomy of R. subsessile, an endangered plant species in the Philippines. It exhibits the morphology typical of plants at high elevations, a hardy shrub with small, tough leaves but with showy pink flowers that rival those of commercial rhododendrons. In this study, new findings on the plant’s morphology include a variable number of stamens and a description of the fruits and seeds. The anatomical features of the species show a typical dicot anatomy with dorsiventral leaves, woody stem and roots with the usual arrangement of periderm followed by secondary phloem,vascular cambium, secondary xylem with vessel elements and pith (in the case of the stem). The biseriate upper epidermis was a unique character of this species. These morphological and anatomical features of the plant could prove valuable as input baseline information in crafting measures for its conservation.
AcknowledgementsSpecial thanks also to Eugene Logatoc for helping in mapping the study site. We would also like to acknowledge the support and generous encouragement from colleagues in the Biology Department.
杂志排行

Journal of Forestry Research的其它文章
- A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing
- Reconciliation of research on forest carbon sequestration and water conservation
- A theory to link relationships of stand volume, density, mean diameter and height in forestry data
- A new model for predicting the total tree height for stems cut-to-length by harvesters in Pinus radiata plantations
- Comparative performances of new and existing indices of crown asymmetry: an evaluation using tall trees of Eucalyptus pilularis(Smith)
- Tree mortality and biomass loss in drought-affected forests of East Texas, USA