Tropical dendrochronology applied to invasive tree species in the Brazilian Atlantic Forest
2021-01-11CarolinaNogueiraXavierDanielaGranatoSouzaAnaCarolinaBarbosaJosReinaldoMoreiradaSilva
Carolina Nogueira Xavier · Daniela Granato-Souza ·Ana Carolina M. C. Barbosa ·José Reinaldo Moreira da Silva
Abstract The Atlantic Forest is one of the largest and most biodiverse rainforests of South America but anthropogenic activities are drastically changing these landscapes.The invasion of alien or exotic species is one of the greatest threats to biodiversity. There are few studies of invasive species in tropical Brazilian ecosystems. This research examines growth and ecological aspects of Syzygium cumini and Clitoria fairchildiana, two invasive tree species in the Pedra Branca State Park, an Atlantic Forest remnant in southeastern Brazil. Both species were successfully dated,indicating an average age of 58 and 31 years, respectively.A positive relationship between growth and precipitation of the previous growing season suggests an ecological adaptive strategy, which could be facilitating their invasion into the environment. Cumulative growth diameter curves indicate slow and fast growth rates for S. cumini and C. fairchildiana,respectively. Growth release episodes detected in the tree rings indicate increased anthropogenic disturbances over the last 50 years.
Keywords Atlantic Forest biome · Invasive species ·Protected areas · Tree rings
Introduction
The Atlantic Forest is one of the largest rainforests of South America, widespread over tropical to subtropical regions and ranging from sea level to ~ 3000 m with high biodiversity. It is considered one of the 25 world hotspots (Myers et al. 2000; Tabarelli et al. 2005; Forzza et al. 2012). In Brazil, most of the Atlantic Forest region also includes some densely populated cities and human disturbances have drastically changed natural landscapes. This has resulted in significant habitat losses over past decades (Myers et al. 2000;Tabarelli et al. 2005; Zenni and Ziller 2011).
The introduction of non-native species may be one of the greatest threats to native urban environments (Gaertner et al. 2017), affecting ecological balance and biodiversity.The first records of exotic species in Brazil date from the early eighteenth century in the southeast (Zenni and Ziller 2011). Intentional or accidental introductions characterize patterns of the invasions but cultural and economic reasons are also important in an urban context (Zenni and Ziller 2011). However, little is known of the ecological aspects of these exotics, and strategies to prevent or reduce invasiveness do not exist or are ineffective (Zenni and Ziller 2011;Gaertner et al. 2017).
Understanding the mechanisms of biological introductions is challenging and a priority for the restoration of disturbed environments (Tabarelli et al. 2005). Dendrochronology is based on the study of tree ring formation (Fritts 1976; Stokes and Smiley 1996) and has provided reliable age-related information on tropical tree species (Stahle et al.1999; Brienen and Zuidema 2005; Schöngart et al. 2015).Based on dendrochronological research, non-native or exotic tree species with distinct growth rings can be analyzed(Closset-Kopp et al. 2011; Nuñez and Paritsis 2018; Brandes et al. 2019; Nogueira et al. 2019). Recent research has shown that the analysis of tree rings may also be applied to understanding growth patterns of non-native species, providing useful information on the introduction, invasiveness, and spatial-temporal distribution, although mainly in temperate environments (Closset-Kopp et al. 2011; Kasson et al. 2013;Nuñez and Paritsis 2018). However, there are few studies with similar approaches in Brazilian tropical environments(Brandes et al. 2019; Nogueira et al. 2019), which mainly concentrated in the development of chronologies with native species (Callado et al. 2001; Fontana et al. 2018; Lisi et al.2008; Aragão et al. 2019). Hence, a study of the growth trajectories of invasive species could contribute to understanding what benef its their establishment in new environments.
Conservation strategies have been applied to Atlantic Forest remnants, with most efforts towards the creation of“protected areas”, that includes parks, reserves, ecological stations and private reserves, and are strictly regulated by Brazilian environmental agencies (Tabarelli et al. 2005).The Pedra Branca State Park is one of the largest urban forests in the world. It is rich in biodiversity and plays an important role in the hydrological cycle of Rio de Janeiro.A list of alien species has been described for Pedra Branca State Park and surrounding area and identified an unusually large presence of three non-native tree species: Artocarpus heterophyllus Lam (Moraceae),Syzygium cumini(L.)Skeels (Myrtaceae) andClitoria fairchildianaR.A.Howard(Fabaceae) (Zenni and Ziller 2011; Fiotec 2016).
Therefore, the goals of this study are to determine if these three species show distinct annual tree rings that can be reliably cross-dated, and the potential of the resulting chronologies to assess tree ages, reconstruct their growth trajectories and detect growth releases that could be related to past disturbances in the forest fragment. The climate-growth relationship of these species is also determined.
Materials and methods
Study site
The Pedra Branca State Park is located in the Rio de Janeiro municipality, southeastern Brazil (22° 56′ 12′ S; 43° 24′ 12′W; Fig. 1), and is considered one of the most representative in area and biodiverse urban parks of the world. It is Brazil’s largest urban forest, covering approximately 12,400 ha(INEA 2015). It was declared a protected area by the State Government of Rio de Janeiro in 1974 and has been managed by the Fundação Oswaldo Cruz since 2007, which has its own campus of approximately 500 ha in the Park’s buffer zone (Fiocruz 2009). This remnant of the Atlantic Forest plays a key role in controlling the water balance of the city of Rio de Janeiro, covering 10% of its territory. The cityis responsible for maintaining 50% of the Atlantic Forest remnants within its area (INEA 2015). Historical occupation has resulted in anthropogenic disturbances of the natural environment over many years, including the introduction of invasive species (Drummond 1997; Costa 2002).

Fig. 1 a Map showing the remaining Atlantic Forest biome distribution (shaded area) and the study site (red dot); b Pedra Branca State Park and surrounding buffer zone delimited by red line, and the study area by white dot. c Distribution of sampled A. heterophyllus (white dots),C. fairchildiana (blue dots) and S. cumini (red dots) within the buffer zone; d Climate diagram of Pedra Branca State Park.Data source: CRU TS4.00 0.5°gridded data set from 1956 to 2018, 16°-24° S, 40°-50° W
According to the Köppen climate classification, the region is humid tropical, with humid summers extending from December to March (average rainfall 800 mm), and the driest season during the winter months of June-August(average rainfall 60 mm) (Fig. 1 d). The average annual precipitation is 1300 mm, and there is little variation over the average temperature of 22 °C. All the climate data were obtained from CRU TS4.01, 16°-24° S and 50°-40° W from 1956 to 2018.
Target species
Artocarpus heterophyllus(Jackfruit) is native to southeastern Asia (Chaves et al. 1967; Ferrão 1993). It is an evergreen, monoecious species, attaining heights of more than 10 m (Chaves et al. 1967; Crane et al. 2002; Fig. 2 a). Flowering usually occurs between October and January (Barbosa 2016) with continuous fruiting over the year, but preferably during the summer months of December-February in Brazil (Cunha et al. 2006). Records of the first introductions date to the end of the seventeenth century in northeastern Brazil (Dean 2002), and the species was widely used for ornamental purposes during the nineteenth century (Santos et al. 2008). The first records of the introduction ofA. heterophyllusin the Rio de Janeiro municipality date back to 1862 in an attempt to recover areas degraded by cane and coffee production (Atala et al. 1966; Bandeira 1993; Dean 2002; Conti et al. 2008).
Syzigium cumini(Java plum) is a native, evergreen species of India and Sri Lanka, reaching heights of 20 m,flowering between September and November (Lorenzi et al.2003), and fruiting January to May (Vizzotto and Fetter 2009; Fig. 2 b).Syzygium cuminihas medicinal properties for treating hypoglycemia (low blood sugars), and with antiinflammatory and antioxidant properties, and hence has been used as alternative medicine by local people (Siani et al.2000; Braga et al. 2007; Lima et al. 2007).Syzygium cuminiusually have dense crowns and have been introduced into Brazilian environments for ornamental purposes (Lorenzi et al. 2003). Additionally,S. cuminiare reported to grow in many soil types and produce polyembryonic fruit; such characteristics may have favored progressive invasion of different Brazilian environments (Gurgel and Soubihe Sobrinho 1951; Donadio 2007).
Clitoria fairchildiana(sombreiro) is a native of the Amazon forests and classified as a light-demanding, pioneer species, most common in secondary forests. It is deciduous,reaching heights of 6-12 m and usually colonizes open,disturbed forests (Lorenzi 1992; Fig. 2 c). Flowering occurs during the boreal summer of April-May, with fruiting in May-July when it starts to shed leaves. It produces approximately 1800 seeds per year (Lorenzi 1992). A large canopy and fast growth rates have madeC. fairchildianaa species widely applied to the recovery of degraded areas (Lorenzi 1992). However,C. fairchildianais allelopathic and not ideal for introduction into its non-native environments(Soares et al. 2002; Sampaio and Schmidt 2014).

Fig. 2 a Artocarpus heterophyllus, b Syzigium cumini and c Clitoria fairchildiana in the buffer zone
Sampling
Cross sections were obtained from legally felled trees ofS.cumini,C. fairchildinanaandA. heterophyllusin the Park’s buffer zone (License no 002920 granted by the Secretaria Municipal de Ambiente e Cultura do Rio de Janeiro-SMAC).Thirty trees > 20 cm diameter at breast height (DBH) weresampled, 10 trees per species. Cross sections were obtained from the base (0.30 cm above ground) for tree-ring analyses.DBH and total height were measured for individual trees(Table 1).
Tree-ring analyses
Cross sections were dried and polished to obtain a clear surface and crossdating carried out using skeleton plots and visual dating under a microscope (Douglass 1941; Stokes and Smiley 1996). Growth ring boundaries of each species were identified and marked according to the recommendations of the International Association of Wood Anatomists(IAWA Committee 1989). At least one radius per specimen was marked and the dated ring widths measured to a precision of 0.001 mm (LINTABTSAPTM; Rinntech 2017). The resulting time series were submitted to dating and measurement quality control with the computer program COFECHA(Holmes 1983).
The time series were detrended and standardized by fitting cubic smoothing splines to remove non-climatic, low frequency growth trends using the program ARSTAN (Cook 1985; Cook and Krusic 2005). The detrending and standardization procedure removed low frequency variance from the relatively short ring width series, where lower frequency variance is more likely to result from age trends and stand dynamics. Tree ring indices were computed by dividing the measured ring width value for each year by the value of the fitted curve for the same year (ratios). The resulting indices of each radius from all trees were averaged on a year-by-year basis to construct the standard tree-ring chronology (Cook 1985; Cook and Krusic 2005).
A series of statistics were computed with the ARSTAN program to describe the internal consistency of the individual ring width series and the resulting standard chronology, [e.g., mean correlation among all series (RBAR; Cook and Pederson 2011), and the expressed population signal(EPS; Wigley et al. 1984)]. The quality of the chronology was evaluated by the value of RBAR, which is the average correlation coefficient resulting from comparing all possible segments of a predetermined length among all the series in the chronology (Briffa 1995). The EPS values provide evidence of the existence of common signals among trees in an inf initely replicated chronology (Wigley et al. 1984;Mérian et al. 2013).

Table 1 Description of the growth features of the tree species in the buffer zone of Pedra Branca State Park.Mean ± SD
Climate-growth relationship
The derived chronologies were used to investigate the relationship between tree growth and climate. Precipitation and temperature data were obtained from the Climatic Research Unit(CRU) TS4.00 0.5° gridded data set (Harris et al. 2014), and a Pearson correlation analysis used with a 95% significance level(Fritts 1976). The analysis was performed with the treeclim package (Zang and Biondi 2015) of R Core Team (2018), and total monthly precipitation from December of the previous growing season (t − 2) to June (t) at the end of the current growing season were computed over 19 consecutive months to identify the strongest monthly or seasonal precipitation signal.
Growth trajectories
The dated ring widths were measured on at least two radii per tree and used to model DBH as a function of tree age.The resulting increments per year were obtained through the average value of the analyzed radii per tree to correct growth variations within the trunk, (e.g., pith eccentricity, anomalies related to distorted trunks). Averaged annual increment rates for each tree were then accumulated to form individual growth curves related to the measured DBH (Brienen and Zuidema 2007; Schöngart 2008).
The study of growth rings provides access to a record of the response of trees growing under natural or disturbed environments. Abrupt changes in annual growth rates over the years reveal disturbance events or periods and can be precisely dated to the calendar year. The running mean method was used to identify growth release episodes on tree ring data performed by the TRADER package in R (Altman et al. 2014). The radial growth averaging method (Nowacki and Abrams 1997) was applied to calculate growth releases on tree ring data. Radial growth averaging identifies the percentage in growth change(% GC) by averaging radial growth over 5- and 10-year periods, forC. fairchildianaandS. cuminirespectively, which were the only two species successfully dated in this study,including the target year (M1), and the average radial growth over the subsequent 10-year period, excluding the target year(M2) (Eq. 1).

The minimum threshold is identified as moderate release with 25% of growth change, and major releases with 50% of growth change (Nowacki and Abrams 1997; Altman et al.2014).
Results
Average DBH and commercial heights of the ten sampled trees of each species,A. heterophyllus,C. fairchildianaandS. cuminiare shown in Table 1. The cross section discs of A.heterophyllus, C fairchildiana andS. cuminishow distinct growth rings, however crossdating was only possible for the last two species by standard dendrochronological methods(Fig. 3). Tree rings are characterized by alternating fiber and parenchyma bands in bothC. fairchildiana(Fig. 3 b) andS.cumini(Fig. 3 c), but are better identified at lower magnification inC. fairchildianasamples because of apparent discontinuity of true tree ring boundaries. Distinct growth rings were also detected inA. heterophyllus, characterized by thick-walled and radially flattened latewood fibers compared with thin-walled earlywood fibers (Fig. 3 a). However, the rings are not regularly distributed over the samples, preventing crossdating by standard dendrochronological methods.Cross sections of 10 trees ofS. cuminiand nine of C.fairchildiana showed satisfactory growth rings, allowing the samples to be cross synchronized and dated, and providing evidence of their yearly growth (Table 2).Syzygium cuminitrees are older, up to 76 years with the youngest at 52 years. There is little variation around the average age of individuals.Clitoria fairchildianatrees are younger, with a maximum age of 47 and half the trees less than 30 years old.
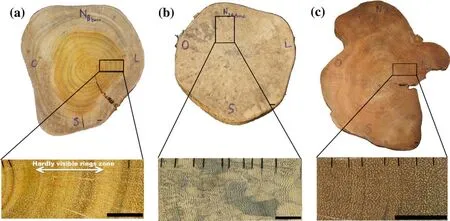
Fig. 3 Growth rings of a Artocarpus heterophyllus, b Clitoria fairchildiana and c Syzigium cumini. True growth rings (black bars) and the zone with indistinct growth ring boundaries (white arrows)(scale = 1 cm)
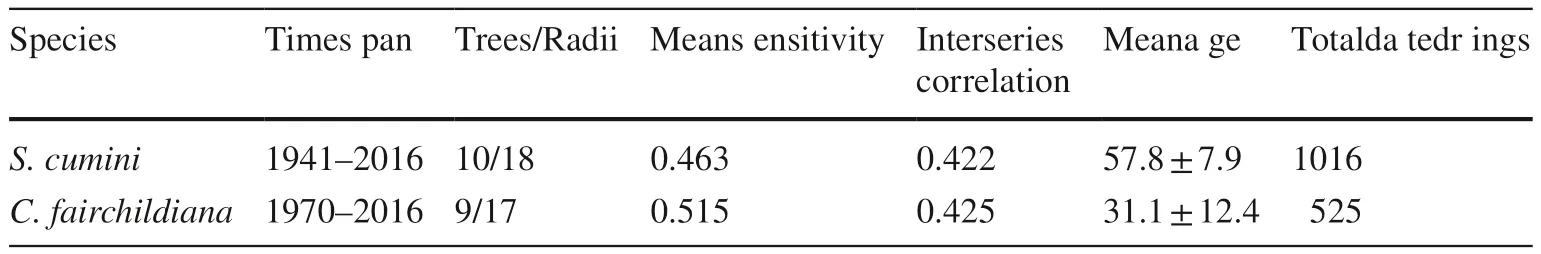
Table 2 Statistical parameters obtained with Cofecha software of S. cumini and C.fairchildiana chronologies
Preliminary standard chronologies ofS. cuminiand C.fairchildiana were developed with 6/12 and 7/14 trees/radii,respectively, showing good internal correlation between individual series (Fig. 4 a, b). Both standard chronologies are positively correlated during the common time interval(1970-2016, r = 0.19). However, the correlation is higher considering 1970-2010 (r = 0.34) (Fig. 4 c). The lagged(t − 1) total annual precipitation in the area is shown in Fig. 4 c.
The chronologies ofS. cuminiandC. fairchildianashow significant correlation with the CRU gridded precipitation data (Fig. 5).Syzygium cuminiis positively correlated with February precipitation (r = 0.46;p< 0.01) and May (r = 0.27;p< 0.05) from the previous growing season (Fig. 5 a).Clitoria fairchildianais positively correlated with January precipitation (r = 0.45;p< 0.01) from the previous rainy season and negatively correlated with precipitation of November(− 0.28;p< 0.05) and March (− 0.29;p< 0.05) of the current growing season (Fig. 5 b). There is no relationship between temperature and the growth of both species. Cumulative diameter growth curves of both species show different patterns, suggesting slower/faster growth rates forS. cuminiand C. fairchildian a, respectively (Fig. 6). Average annual increments of 3.5 mm/year (± 2.5 SD) were verified forS. cuminiand 5.5 mm/year (± 4.1 SD) forC. fairchildiana.
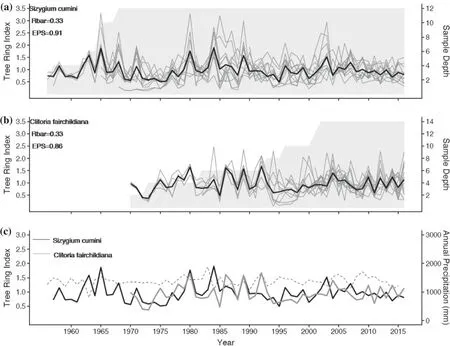
Fig. 4 Dated, detrended, and standardized radii (gray line) and mean index standard chronology (black line) of a Syzygium cumini (1956-2016) and b Clitoria fairchildiana (1970-2016); sample depths shown by shaded areas (a, b). Mean RBAR and EPS values obtained with the ARSTAN program are indicated; similarities of final index standard chronologies and c the total annual precipitation lagged 1 year (t − 1) shown at the bottom
The radial growth averaging method shows disturbance events in 68% of the total trees ofS. cuminiandC. fairchildiana(Fig. 7). EightS. cuminiand five out of nine C.fairchildiana showed at least one release episode during their complete growth trajectories. Major release episodes dominated the forest disturbances observed within the analyzedS. cuminitrees. Most release events occurred between 1970 and 2002 when trees experienced moderate to major disturbance events. More than one release episode was identified in the growth trajectories of fourS. cuminiand oneC. fairchildianatrees. The oldestS. cuminigerminated in 1941 while the majority of the individuals of this species germinated during the 1960s. Trees ofC. fairchildianashow a more uneven age distribution.
Discussion
Distinct annual growth rings are reported for the first time for two invasive species,Syzygium cuminiandClitoria fairchildiana. Growth rings were successfully crossdated by standard dendrochronological techniques, and chronology statistics and internal coherence within and between trees support the annual nature of the growth rings in both species(Stahle 1999). Previous studies have described poorly visible and erratic growth rings byS. cuminiin western Kenya and the Western Ghats of India (Nath et al. 2012; David et al.2014). However, tree-ring distinctiveness can be related to the environmental conditions where the trees are growing,(e.g., climate factors influencing local variability, stand and ecological parameters, and even different soil conditions).Working on larger cross-sectional samples allows a clearer view of the true growth ring boundaries, hence facilitating cross-dating between trees in tropical environments (Worbes 1985; Stahle et al. 1999; López et al. 2017; Barbosa et al.2018).Artocarpus heterophyllustrees showed distinct growth rings but discontinuous and barely visible in some portions of the cross-sections and could not be crossdated with standard dendrochronological techniques.

Fig. 5 Monthly values for CRU TS4.00 gridded precipitation correlated with standard chronologies of a Syzygium cumini from 1956 to 2016 and b Clitoria fairchildiana from 1970 to 2016 to identify monthly precipitation totals most important to radial growth of both species for a 19-month period, including previous rainy and dry sea-sons (lowercase letters) and the year concurrent with growth (uppercase letters). Filled black bars represent significant correlations. Gray area is seasonal precipitation (CRU TS4.00 0.5° gridded data set from 1956 to 2016)

Fig. 6 Cumulative ring-width increments for a S. cumini and b C. fairchildiana; Loessfunctions fitted to cumulative diameters (bold lines) represent smoothed conditional means(shaded areas are 95% conf idence intervals of the loessfunctions)

Fig. 7 Dates of release events per tree according to growth averaging method analysis of S. cumini and C. fairchildiana; moderate releases (black square) and major releases(black triangle)
The chronologies obtained ofS. cuminiandC. fairchildianashow that the growth of both species was positively influenced by rainfall of the previous rainy season. In fact,both species showed similar growth-climate responses,resulting in positive correlations between chronologies over their lifespan. It is interesting that most native species in the Atlantic Forest environment show positive correlations between growth of the trees and precipitation of the current growing season (Latorraca et al. 2015; Souza et al. 2016;Fontana et al. 2018; Aragão et al. 2019). Moisture-sensitive tree species tend to grow and accumulate nutrients during the rainy season (Fritts 1976; Dünisch et al. 2003; Kunert et al. 2010) between September and April in the study area(Fig. 1 d). However, a few native species have shown similar climate-growth relationships asS. cuminiandC. fairchildiana(Pagotto et al. 2015; Aragão et al. 2019). One hypothesis is that they may have been producing and storing nutrients during the previous growing season which are likely used for secondary growth during the current growing season (Fritts 1976; Oliveira et al. 2009). It also may suggest an adaptive strategy to produce energy for flowering and fruiting during the current growing season, a period of high energy costs for plants (Lorenzi 1992; Woodhouse 1997; Lorenzi et al.2003; Evert 2014), thus enhancing their successful establishment in a foreign environment. If they colonize a new area and become established over the native species, this growth strategy could be one of the reasons why they have successfully invaded the exotic environment.
Growth trajectory analyses show older and homogeneous age distribution among individuals ofSyzygium cumini.The lower growth rates reported in this study conf irm the behavior ofS. cuminigrowing in Indian environments where trees may attain more than 100 years of age, suggesting this species is long-lived (Nath et al. 2012). A young, heterogeneous age distribution and relatively fast growth rates characterize the growth ofC. fairchildiana. Their fast growth and high nitrogen fixation have been described in previous studies, suggesting it as a potential species for the recovery of degraded areas (Lorenzi 1992; Nobre et al. 2010; Costa et al. 2014). BothS. cuminiandC. fairchildianashow similar average annual increments to those found for native species in the Atlantic Forest environment (Barbosa et al. 2018;Fontana et al. 2018) and do not show stabilized growth rates,which adds to the knowledge of the lifespan of these invasive non-native species.
Tree ring analyses showed that the oldestS. cuminiindividual dates from 1941, pointing to a successful establishment of the species, with the rest of the trees dating back to the 1960s. The oldestC. fairchildianatree dates from 1970,showing most individuals are relatively young. It is possible that these dates do not represent the first introduction episodes of both species in the area and may be more likely to be natural recruitment. Historical records describe intense human activity such as cane sugar and cassava production and cattle breeding between the eighteenth and nineteenth century in the area where the Pedra Branca State Park is located (Drummond 1997). The area was expropriated by the federal government at the beginning of the twentieth century in order to recover the native vegetation and act as a protection of hydrological resources (Correa 1936; Costa 2002). Therefore, the trees in this study are regrowth of the first introductions and have successfully been colonizing the Atlantic Forest remnants for decades. A recent survey has recorded large populations for the alien species in this study,and raises concern by the administration of the Pedra Branca State Park (Fiotec 2016). This study shows how tree rings may be a reliable tool to access historical records and also provide key information about the status of ongoing species invasions in natural environments, one of the biggest threats to biodiversity conservation.
Moderate to major release episodes were detected in the growth of almost all the study trees ofS. cuminiand C.fairchildiana, primarily at the end of the twentieth century.Under natural and undisturbed environments, tree- growth variation is due to a combination of age-related trends,regional climate scale patterns, and stand exogenous and endogenous disturbances, and thus the effects of climate usually create a common signal between trees which allow them to be synchronized and dated (Fritts 1976; Cook 1990;Stokes and Smiley 1996). Local endogenous disturbances,in turns, create individual signals in each tree, and thus suppressed growth usually represents strong competition under a dense canopy (Tilman 1988; Cook 1990). Growth release episodes reflect the response of the trees to canopy openings, illustrating an important indicator of forest dynamics(Nowacki and Abrams 1997; Tanner et al. 2014; Abiyu et al.2018).
Past records of forest disturbances can be accessed through the identification of the growth release events in each study tree and anthropogenic disturbances have been reported as the most important modifiers of forest dynamics and composition over the last century (McCann 1997;Bräuning et al. 2013; Abiyu et al. 2018). Despite the relatively longS. cuminichronology dating back to 1941, the first release episode was only detected in 1964, becoming more frequent after 1970, as well as an increased number of both non-native species in the area. This may represent an increasing effect of human-induced forest disturbance over the last 50 years, thus justifying the conservation status of the Pedra Branca State Park. Moreover, the detected growth release episodes show thatS. cuminiandC. fairchildianahave the potential to benef it from recent canopy openings and hence may be taking advantage to successfully replace native species.
This study presents information to help fill the gaps regarding the behavior of two non-native, invasive species in southeastern Brazil. Further research should also consider including tree-ring analyses with native species growing in the same forest fragment. This would provide interesting comparisons between climate-growth relationships and also improve analysis of growth trajectories, adding reliable contributions to the actual invasive status of the study area.
Conclusions
The Atlantic Forest of Brazil is one of the largest rainforests in South America, accounting for high biodiversity but is also a highly degraded environment due to the effects of human disturbances over many decades. The Brazilian government has implemented protected areas in an attempt to reduce habitat losses, and thus urban parks have been created to conserve the remaining vegetation.However, accidental or intentional introductions of alien species represents one of the greatest threats to the conservation of the native urban environments, and still,relatively little is known about the biological aspects of the invasions. Tree- ring analyses of Syzigium cumini andClitoria fairchildianaprovide interesting contributions on the ecological aspects of both species. Tree rings were successfully dated with standard dendrochronological techniques in both species. The climate- growth relationship shows a positive correlation between tree growth and precipitation of the previous growing season, which may represent a strategy to establish and invade the Atlantic Forest fragment. Growth trajectory analyses show slower growth rates forS. cuminiand faster ones forC. fairchildiana. Growth release episodes identified for both species, mainly after the 1970s, suggest an increasing effect of human disturbances over the last 50 years. Future studies should consider a broader approach with a larger number of samples to improve the outcomes that could help clarify the biological invasions in the Atlantic Forest remnant in southeastern Brazil.
AcknowledgementsThe authors thank Fiocruz Campus Mata Atlântica for donating material and field support.
Open AccessThis article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source,provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creat iveco mmons.org/licen ses/by/4.0/.
杂志排行

Journal of Forestry Research的其它文章
- A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing
- Reconciliation of research on forest carbon sequestration and water conservation
- A theory to link relationships of stand volume, density, mean diameter and height in forestry data
- A new model for predicting the total tree height for stems cut-to-length by harvesters in Pinus radiata plantations
- Comparative performances of new and existing indices of crown asymmetry: an evaluation using tall trees of Eucalyptus pilularis(Smith)
- Tree mortality and biomass loss in drought-affected forests of East Texas, USA