三倍体丹参杂交种的花粉形态研究
2021-01-07庞俊秀薛惠芬刘婉秋龙鸿
庞俊秀 薛惠芬 刘婉秋 龙鸿


摘 要: 三倍體丹参是以二倍体丹参为父本、人工染色体加倍的四倍体白花丹参为母本杂交选育的杂交种。为深入了解三倍体丹参花粉的特性,以及为三倍体种质利用提供孢粉学依据,该文以二倍体丹参为对照,研究了三倍体丹参杂交种花粉的形态变异规律。利用光学显微镜和扫描电镜对二倍体和三倍体丹参的花粉萌发沟、外壁纹饰、花粉粒形状等特征进行了显微和超微形态观察,综合进行了花粉形态差异比较,并对花粉大小和形状数据进行了差异显著性分析和正态检验。结果表明:(1)二倍体丹参为6沟花粉,三倍体花粉萌发沟有6沟和8沟两种类型,沟内疣状颗粒分布不匀,出现畸形萌发沟。(2)二倍体和三倍体花粉外壁均为网状雕纹。二倍体花粉网眼内具多个多边形穿孔,穿孔大;6沟和8沟两种类型的三倍体花粉网眼无穿孔或仅有几个小穿孔,6沟和8沟花粉的外壁雕纹相同。(3)三倍体花粉的极轴长(P)和赤道宽(E)均值显著小于二倍体花粉,花粉大小呈偏正态分布,P*E的差异系数大于二倍体花粉,且有极值存在。三倍体和二倍体丹参的萌发沟和雕纹存在差异,而花粉形状差异不显著。综上结果表明三倍体丹参花粉在倍性效应和杂合性的双重影响下发生了形态变异,且有多种形态变化。
关键词: 丹参, 白花丹参(变型), 二倍体, 三倍体, 花粉形态
中图分类号: Q938; S567.5+3
文献标识码: A
文章编号: 1000-3142(2021)12-1996-08
收稿日期: 2020-07-27
基金项目: 国家级大学生创新创业训练计划项目(201810061004) [Supported by the National Innovation and Entrepreneurship Training Program for College Students (201810061004)]。
作者简介: 庞俊秀(1999-),主要从事植物资源鉴定评价研究,(E-mail)2690390206@qq.com。
通信作者: 龙鸿,博士,教授,主要从事果树遗传、发育生物学研究,(E-mail)longhong@tjau.edu.cn。
Pollen morphology of triploid Salvia miltiorrhiza hybrid
PANG Junxiu, XUE Huifen, LIU Wanqiu, LONG Hong*
( College of Horticulture and Landscape, Tianjin Agricultural University, Tianjin 300384 )
Abstract: To study pollen characteristics of triploid Salvia miltiorrhiza (tetraploid S. miltiorrhiza f. alba × diploid S. miltiorrhiza) hybrid, morphological variation of triploid pollen during the triploid process was examined. Triploid and diploid pollen were observed at microscopic and ultra-microscopic level using light and scanning electron microscopes. Pollen size and shape data were analyzed to determine statistical significance. The results were as follows: (1) Compared to typical 6-colpate diploid pollen grains, triploid S. miltiorrhiza are 6- or 8-colpate, with increased number of colpi and heterogenous granules on colpus membranes, and abnormal colpate. (2) Exine sculpturing of triploid pollen was reticulate like the diploid. Both 6- and 8-colpate triploid pollen had no perforation or a few small perforations, whereas diploid pollen had many large polygonal perforations in each lumen. (3) In terms of size, mean of the polar axis (P) and equatorial axis (E) in triploid pollen were significantly smaller than that of diploid pollen. The pollen size of triploid hybrid followed a skewed normal distribution, and the coefficient of variation (CV) in P*E value of triploid was greater than that of diploid and existed extreme values. Comparison data revealed variance in colpi and sculpture between triploid and diploid, however no significant differences in pollen shape is found. Overall, the triploid S. miltiorrhiza pollen has more morphological variations due to genetic effect of both ploidy level and heterozygosity.
Key words: Salvia miltiorrhiza, S. miltiorrhiza f. alba, diploid, triploid, pollen morphology
丹参(Salvia miltiorrhiza)是唇形科(Labiatae)鼠尾草属(Salvia Linn.)多年生草本植物,其干燥根和根茎为传统中药,具有活血祛瘀、通经止痛、清新除烦等功效(国家药典委员会,2015)。丹参为大宗药材,自20世纪70年代中期野生丹参已不能满足需求,市售丹参以栽培品为主,栽培种源多来自当地野生居群,未经过人工选择。丹参种植主要采用种子繁殖和根段繁殖,长期以来,品种只种不选,从而导致品种混杂、种性退化、药材产量和质量下降(高山林等,1995)。
种间杂交(hybridization)和多倍体化(polyploidization)是引入遗传变异的重要手段,多倍体品种通常在生长活力、抗逆性等方面优于二倍体品种,药用植物多倍化显著增加次级代谢产物的含量(Salma et al., 2017)。野生丹参植株为二倍体,多倍体育种始于1987年,四倍体植株在田间性状、产量、药材外观、化学成分含量、药理活性等方面均优于二倍体(高山林等,1995)。有研究以紫色花为标记性状,将白花丹参(变型)(S. miltiorrhiza f. alba)染色体加倍,以四倍体白花丹参植株为母本,与来自不同地区的二倍体紫花丹参杂交,选育出性状优良的丹参三倍体杂交种(陈力和李秀兰,2008;李秀兰和陈力,2012)。白花丹参(变型)于2018年由李晓娟等鉴定为中华白花丹参(S. sinealba)(李晓娟等,2018)。三倍体丹参在植株形态上保留了多倍化优势,与二倍体紫花丹参相比叶更宽,叶色更深,长势更好,根条数、根部鲜质量增加;在主要药物成分上,三倍体丹参表现出杂交种优势,隐丹参酮和丹参酮ⅡA含量均高于二倍体和四倍体亲本,丹酚酸B和丹参酮Ⅰ含量介于亲本之间(李秀兰和陈力,2012)。三倍体丹参经组培扩繁后已应用于大田生产。
生产实践中,三倍体还可作为载体将携带的亲本基因型转移到其他物种中(Ehlenfeldt & Hanneman, 1984)。以三倍体郁金香作为花粉供体的杂交组合显示,三倍体亲本产生了非整倍体和整倍体(x,2x,3x)配子,并且回交后代(BC1)中存在大量基因组重组(Marasek-Ciolakowska et al., 2014)。杨树(Populus spp.)育种中,三倍体也多次作为桥梁亲本,育成了三倍体、四倍体和非整倍体后代(李代丽等,2019)。
花粉形态由基因控制,受外界环境影响较小,具较强的遗传保守性,是开展三倍体种质利用的基础数据。以花粉大小、形状、穿孔直径和网脊宽度等参数对不同倍性的芍药花粉形态聚类分析发现,有共同亲本Paeonia lactiflora的四倍体品种和二倍体品种聚为一组,而同样以P. lactiflora为共同亲本的三倍体品种,因有另一个共同亲本P. officinalis而聚在另外一组,表明花粉形态可用于雜交种亲缘关系的鉴定并为杂交育种提供指导(Hao et al., 2016)。鼠尾草属植物通常为6沟花粉(Henderson et al., 1968)。Maurizio(1956)报道草原鼠尾草(Salvia pratensis)和一串红(S. splendens)的2n品系花粉均为6沟,3n、4n品系花粉中则产生6~12沟多种萌发沟数量的花粉(Henderson et al., 1968; 埃尔特曼,1978),说明鼠尾草属的多倍体品系花粉随倍性增加萌发沟数量增加,且萌发沟数量出现分化。花粉大小与核内 DNA 含量呈正相关,在四倍体、六倍体等多倍体的种类中,花粉比双倍体大(埃尔特曼,1978)。青黑杨杂种三倍体饱满花粉直径的变化区间远大于相同杂交组合二倍体雄株的花粉变化,推测是由于三倍体复杂的染色体分离行为,使得花粉中含有不同的染色体数目,从而使三倍体产生大小不一的花粉(李代丽等,2019)。这些研究都表明,三倍体花粉的形态既受亲本影响,又因杂交和倍性的影响而产生变化。
三倍体丹参兼具多倍体和杂交种双重优势,可作为丹参远缘杂交和多倍体育种的亲本试材(张铌璇等,2019)。二倍体丹参花粉形态已有报道(徐任生,1990;杨德奎等,2003),李晓娟(2013)研究了丹参和白花丹参(变型)花粉的形态差异,三倍体丹参是以二倍体丹参为父本、以四倍体白花丹参(变型)为母本的杂交种,但其花粉形态尚无孢粉学特征分析。本文以二倍体丹参为对照,研究三倍体丹参杂交种花粉的外壁纹饰、萌发沟、花粉粒形状等花粉的形态特征,为三倍体丹参种质的利用、丹参杂交育种提供孢粉学依据。
1 材料与方法
1.1 材料
丹参为市场销售的普通丹参种苗,产地为河南伏牛山。四倍体白花丹参×二倍体丹参杂交选育的三倍体材料由南开大学生命科学学院陈力教授惠赠,为三倍体组培苗,经炼苗后移栽至天津农学院实验室。室温条件下盆栽种植,营养土∶蛭石=1∶1,高温灭菌冷却后埋根,每个长形塑料盆中放2~3个根茎,株距10~15 cm。于丹参盛花期(2019年4月)选择丹参二倍体、三倍体花序相同部位、刚展开的新鲜花朵,用镊子剥取花药,分别置于干净的1.5 mL的离心管内保存。一部分花药用于光学显微镜(light microscope, LM)观察,另一部分花药用于扫描电镜(scanning electron microscope, SEM)观察。
1.2 方法
1.2.1 花粉形态观察 光学显微镜观察: 将花粉抖落在载玻片上,滴蒸馏水,盖上盖玻片。用LEICA DM4000B正置显微镜在(1 000×)的视野下选择分散、独立的花粉粒,先用LAICA DFC 450数字相机拍照,再用LEICA LAS软件计数及测量花粉的长和宽。
扫描电镜观察: 参照顾欣等(2013)、张永芳等(2016)处理花粉的方法,先用签字笔做好标记,用pH=7.0的磷酸缓冲液漂洗,然后用3%戊二醛固定,置于4 ℃冰箱过夜,固定后用pH=7.0的磷酸缓冲液漂洗3次(每次间隔15 min),再进行乙醇梯度脱水(30%、50%、70%、80%、90%各漂洗15 min,无水乙醇漂洗3次,每次间隔10 min),脱水完毕后进行干燥,最后粘样品台,记录好台号对应的花粉倍性,上机测试。在FEI Quanta 200扫描电镜下观察花粉粒赤道面和极面,加速电压15 kV,挑选饱满、具有代表性、完整的花粉粒观察萌发孔类型和外壁纹饰以及进行拍照,使用Adobe Photoshop CS6软件制作图版。
1.2.2 数据分析 对照扫描电镜下花粉形态,将光学显微镜下拍摄的花粉照片分类,用花粉赤道面观的长和宽测量值作为花粉粒极轴(P)长度和赤道轴(E)宽度,按孢粉学研究方法计算P*E、P/E值,其中P*E代表花粉大小,P/E代表花粉形状类型。花粉极轴长度10~20 μm属于小花粉,25~50 μm属于中等大小花粉,50~100 μm属于大花粉(王开发和王宪曾,1983)。利用SPSS软件(IBM SPSS Statistics 25版本)分析花粉测量数据。
2 结果与分析
2.1 丹参二倍体、三倍体花粉形态对比
2.1.1萌发沟 丹参二倍体、三倍体花粉外观相近,均为黄色、单粒花粉(图版Ⅰ)。二倍体花粉光学显微镜(图版I:A)和扫描电镜下(图版I:D)赤道面观都可观察到3条等距离萌发沟,沟长近两极,沟深,不形成合沟。花粉侧面观呈扁球形,沟形态与赤道面观相同(图版I:E)。极面观可见6条萌发沟(图版I:B,C,F,G):在光镜下,二倍体花粉极面随拍摄景深不同,6条萌发沟呈米字形排列(图版I:B),或在花粉边缘呈6个凹槽(图版I:C); 电镜下二倍体花粉极面观呈扁长的串珠形,萌发沟凹陷(图版I:F),或呈椭圆形,萌发沟不凹陷(图版I:G)。二倍体花粉萌发沟数量和形状与前人(徐任生,1990;杨德奎等,2003;李晓娟,2013)报道一致。
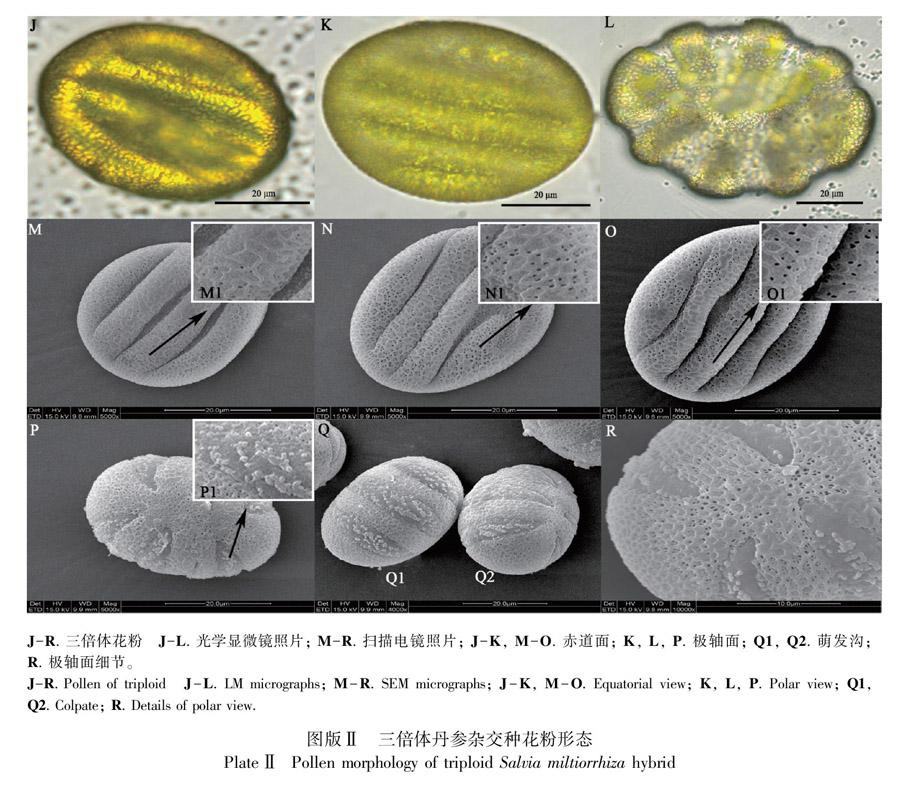
三倍体花粉在光学显微镜和扫描电镜均观察到赤道面观萌发沟数量有变异,呈6条(图版Ⅱ:J,M,N,P)或8条(图版Ⅱ:K,L,O)萌发沟两种类型。与二倍体花粉萌发沟相比,同一个花粉上的萌发沟宽度和深度更不规则,或出现畸形萌发沟(图版Ⅱ:N)。二倍体和三倍体花粉粒的萌发沟内均匀密布疣状颗粒,二倍体萌发沟内的颗粒物分布密度较均匀(图版I:G1,H1,H2),三倍体萌发沟内的颗粒成簇分布,簇内颗粒紧密排列,簇间空隙较二倍体的宽(图版Ⅱ:P1,Q1,Q2)。三倍体萌发沟数量有随倍性增加而增加的多倍化特征;三倍体萌发沟畸形和沟内颗粒分布变化表明三倍体花粉壁形成过程中某些代谢活动异常。
2.1.2 外壁纹饰 丹参二倍体、三倍体花粉外壁均为网状雕纹,网脊粗、平滑,网脊呈多角形。扫描电镜下二倍体花粉赤道面和侧面外壁雕纹网眼内具多个大小不一的多边形穿孔,穿孔之间的次生网脊薄,穿孔布满网眼,看上去较“透亮”(图版I:D1)。三倍体花粉的赤道面外壁多数网眼内无穿孔,或穿孔数少,孔小。3条萌发沟和4条萌发沟的三倍体花粉赤道面外壁特征相同(图版Ⅱ:M1,N1,O1)。二倍体和三倍体花粉的极面观外壁纹饰相近,在极顶处网眼变小,穿孔也变小(图版I:I;图版Ⅱ:R)。
二倍体花粉雕纹与前人(徐任生,1990;杨德奎等,2003;李晓娟,2013)报道一致,三倍体花粉雕纹与李晓娟(2013)报道的白花丹参(变型)接近。徐任生(1990)描述丹参的花粉外壁在光学显微镜下有不均匀细网状纹饰,本试验中,二倍体和三倍体花粉在光学显微镜下均可分辨出外壁的网状纹饰,二倍体花粉比三倍体花粉外壁网状纹饰更清晰,推测与两者外壁穿孔数量不同、反光程度不同有关。
2.2 花粉大小和形状
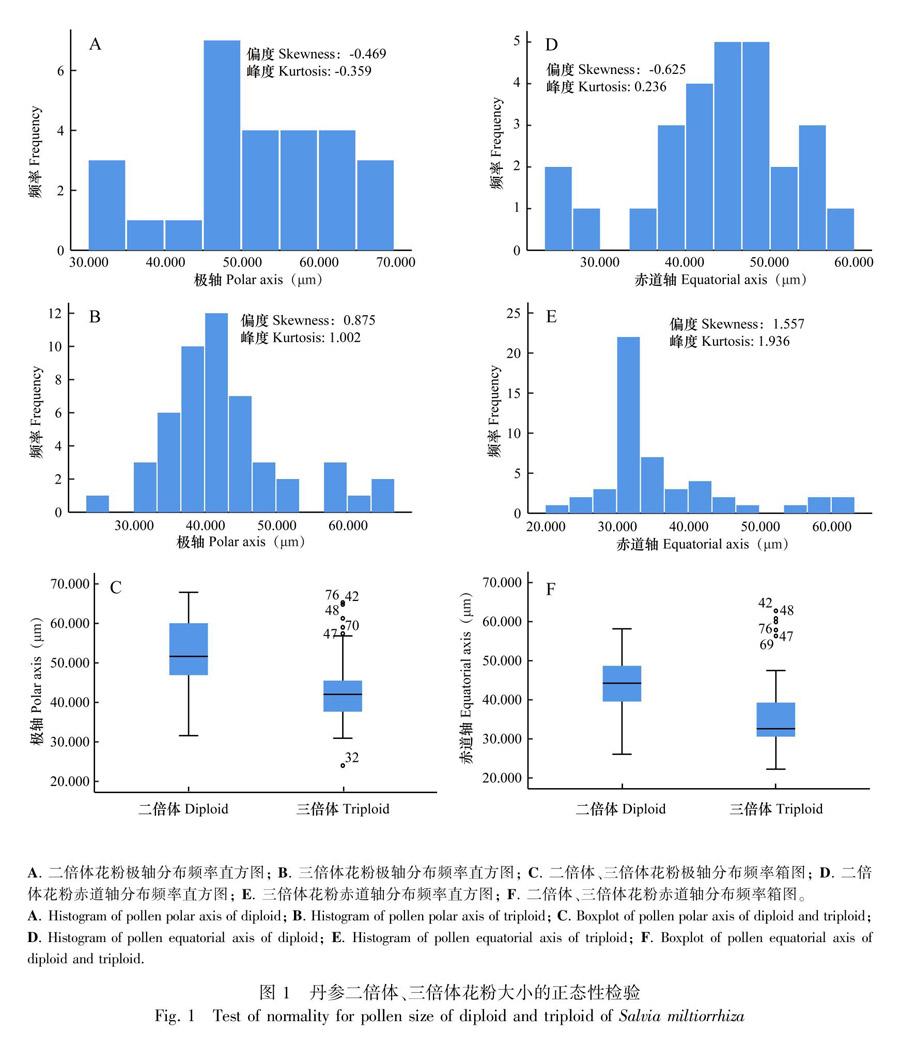
2.2.1 花粉大小 二倍体和三倍体花粉分别测量和拍摄了70和75粒花粉。与扫描电镜照片相比,二倍体赤道面观花粉27粒,三倍体赤道面观花粉50粒。丹参二倍体花粉极轴(P)长为31.57~67.86 μm,均值为52.03 μm;三倍体花粉极轴长为24.00~65.28 μm,均值为43.07 μm。二倍体花粉赤道轴(E)宽为26.06~58.16 μm,均值為43.83 μm;三倍体花粉赤道轴宽为22.25~62.75 μm,均值为36.23 μm。三倍体花粉P值区间与二倍体相近、E值区间大于二倍体。但P、E以及P*E的均值均显著小于二倍体花粉(P<0.01)(表1)。
二倍体和三倍体花粉都产生两种大小类型的花粉, 即中等花粉和大花粉。二倍体和三倍体花粉极轴长的变异系数相同,均为20%,三倍体花粉的赤道轴宽变异系数(26%)大于二倍体花粉(18%)。二倍体花粉P*E值的变异系数为35%,三倍体花粉P*E值的变异系数为49%。由三倍体花粉的P、E极值区间和P*E值的变异系数增大可知,三倍体花粉粒大小的差异幅度比二倍体花粉大。对二倍体和三倍体花粉的P、E测量数据进行正态分布检验发现,虽然二倍体和三倍体花粉测量数据的变异系数相近,但三倍体花粉大小偏向左侧,P和E的偏度绝对值(分别为0.875和1.557,图1:B, E)约为二倍体花粉P、E偏度绝对值(分别为-0.469和-0.625,图1:A,D)的2倍,峰度也大于二倍体数据。二倍体花粉P和E没有极值,三倍体花粉P和E均有极值存在(图1:C,F),表明三倍体花粉中,P和E值小于均值的花粉数量较多,但又有个别花粉粒的P、E值偏大。
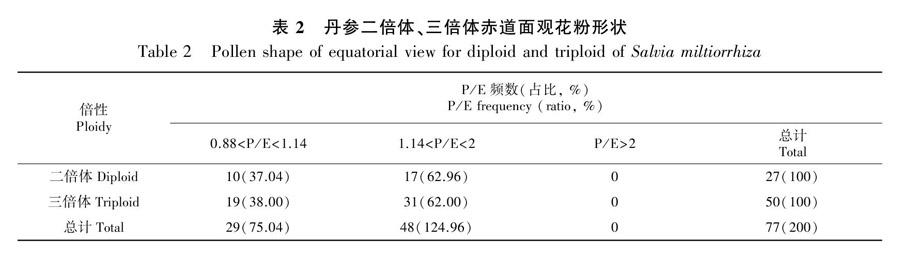
2.2.2 花粉形状 孢粉学中,花粉形状以P/E表示,其中P/E<0.88为扁球形,0.88
2为超长球形(王开发和王宪曾,1983)。二倍体和三倍体花粉P/E值均在0.88~1.14和1.14~2范围内,花粉形状为近球形和长球形两种类型,符合鼠尾草属花粉的一般特征(Henderson et al., 1968)。二倍体P/E的均值为1.19±0.11,三倍体的P/E均值为1.21±0.15,差异不显著(P>0.05)。对倍性与花粉形状两因素做独立性Χ2检验,P>0.05,表明花粉粒形状不受倍性影响(表2)。
3 讨论与结论
花粉形态在科、属间稳定,花粉外壁纹饰的网眼直径、网眼密度差异明显,是种的分类依据,也是鉴定栽培品种亲缘关系的依据(史艳财等,2020)。本研究结果中,三倍体花粉在萌发沟数量、外壁雕纹、花粉大小上均与二倍体不同,且花粉萌发沟有6沟和8沟两种类型,花粉大小的变异系数增大,表明三倍体花粉在倍性效应和杂合性的双重影响下发生了形态变异,且有多种形态变化。鼠尾草属植物花粉的一般特征是花粉形状为近球形和长球形,具有对称的6条萌发沟和多边形网状雕纹,不同物种的沟数相同、网状雕纹内的穿孔数量和大小不同,萌发沟数量随倍性增加而增加(Henderson et al., 1968)。在本研究中,二倍体丹参材料为普通市售丹参种苗,花粉形状、大小、萌发沟数量和外壁雕纹形状与不同时间的不同研究者(徐任生,1990;杨德奎等,2003;李晓娟,2013)的报道一致,表明丹参的花粉形态非常稳定。三倍体花粉形状为近球形和长球形,有对称的6条或8条萌发沟,多边形网状雕纹,符合鼠尾草属属内花粉的共同特征,兼具多倍体品系萌发沟数量增加的多倍化特征。萌发沟数量随倍性增加在其他多倍体植物花粉中亦有报道(刘文革等,2003;Lazarevic' et al., 2013;王尚德等,2015),但形成机制未知。4n品系主要产生 2n配子,从鼠尾草属4n品系中以8沟花粉常见,少数为6~12沟(Henderson et al., 1968)推测,三倍体丹参中的8沟花粉可能与花粉倍性增加有关。
三倍体花粉的外壁网状雕纹网眼内没有穿孔或只有少量的小穿孔的形态偏向于李晓娟(2013)描述的未加倍的二倍体白花丹参(变型),且雕纹形态与萌发沟数量无关。目前,四倍体白花丹参的花粉形态尚未有报道,加倍后的白花丹参四倍体基因组在花粉形态上是否也具有这种遗传偏好性值得进一步研究,以增进对三倍体丹参花粉外壁雕纹变异规律的理解。
花粉粒大小是孢粉学研究的重要参数。本研究中三倍體丹参花粉的P、E均值显著小于二倍体花粉,P、E的数据均呈偏正态分布,出现花粉极值,即三倍体产生了更多的偏小花粉和少量大花粉。青黑杨三倍体杂交种花粉均值大于同组合二倍体杂交种,花粉最小值和最大值的变化范围大于二倍体杂交种,数据呈偏正态分布(李代丽等,2019);桦属(Betula)三倍体种间杂交种产生两种大小的花粉,小花粉数量多,花粉大小分布呈现2个峰值(Karlsdottir et al., 2008);芍药的三倍体花粉小于二倍体花粉(Hao et al., 2016);西瓜三倍体的花粉大小介于四倍体和二倍体之间(刘文革等,2003)。这些研究中,与二倍体相比,三倍体花粉大小结果均不一致。考虑到三倍体复杂的减数分裂行为,三倍体花粉中含有的染色体数目变异丰富,花粉大小的统计数值范围增大,数值大小的频数呈偏正态分布,通过花粉形态鉴定不同倍性品种的亲缘关系研究,应对花粉大小数据进行检验和较正,避免鉴定结果产生偏差。
参考文献:
CHEN L, LI XL, 2008. Natural hybridization seed-production method of triploid radix Salvia miltiorrhizae using purple gene as label character: CN200810152847.6 [P]. 2008-11-06. [陈力, 李秀兰, 2008. 以紫色基因为标志性状的三倍体丹参的自然杂交制种方法:CN200810152847.6 [P]. 2008-11-06.]
Chinese Pharmacopoeia Commission, 2015. Pharmacopoeia of the People’s Republic of China (Volumes I) [M]. Beijing: China Medical Science Press: 76-77. [国家药典委员会, 2015. 中华人民共和国药典(一部) [M]. 北京: 中国医药科技出版社: 76-77.]
EHLENFELDT MK, HANNEMAN RE, 1984. The use of Endosperm Balance Number and 2n gametes to transfer exotic germplasm in potato [J]. Theor Appl Genet, 68(1-2): 155-161.
ERDTMAN G, 1978. Handbook of palynology [M]. Beijing: Science Press. [埃尔特曼, 1978. 孢粉学手册 [M]. 北京: 科学出版社.]
GAO SL, XU DR, CAI ZH, et al., 1995. Selection and identification of tetraploid elite line 61-2-22 of Salvia miltiorrhiza [J]. Chin J Chin Mat Med, 20(6): 335-337. [高山林, 徐德然, 蔡朝晖, 等, 1995. 丹参四倍体优良新品系61-2-22的选育与鉴定 [J]. 中国中药杂志, 20(6): 335-337.]
GU X, ZHANG YL, NIU LX, 2013. Pollen morphology observation of 15 wild lilies from four provinces in western China [J]. Acta Hortic Sin, 40(7): 1389-1398. [顾欣, 张延龙, 牛立新, 2013. 中国西部四省15种野生百合花粉形态研究 [J]. 园艺学报, 40(7): 1389-1398.]
HAO LH, MA H, TEIXEIRA DA SILVA JA, et al., 2016. Pollen morphology of herbaceous peonies (Paeonia L.) with different ploidy levels [J]. J Am Soc Hortic Sci, 141(3): 275-284.
HENDERSON DM, PRENTICE H, HEDGE IC, 1968. Pollen morphology of Salvia and some related genera [J]. Grana, 8(1): 70-85.
KARLSDOTTIR L, HALLSDOTTIR M, THORSSON AT, et al., 2008. Characteristics of pollen from natural triploid Betula hybrids [J]. Grana, 47(1): 52-59.
LAZAREVIC' M, SILJAK-YAKOVLEV S, LAZAREVIC' P, et al., 2013. Pollen and seed morphology of resurrection plants from the genus Ramonda (Gesneriaceae): Relationship with ploidy level and relevance to their ecology and identification [J]. Turk J Bot, 37(5): 872-885.
LI DL, SHANG J, TIAN J, et al., 2019. Meiotic chromosome behavior of pollen mother cells and pollen variation in triploid hybrid between section Tacamahaca and sect. Aigeiros of Populus [J]. J Beijing For Univ, 41(7): 75-82. [李代麗, 商静, 田菊, 等, 2019. 青黑杨杂种三倍体花粉母细胞减数分裂染色体行为及其花粉变异研究 [J]. 北京林业大学学报, 41(7): 75-82.]
LI XJ, 2013. The research on classification and identification of Salvia shandongensis [D]. Jinan: Shandong University of Traditional Chinese Medicine. [李晓娟, 2013. 山东丹参与其近缘种的分类鉴定研究 [D]. 济南: 山东中医药大学.]
LI XJ, LI JX, XU N, et al., 2018. Morphological classification and identification of Salvia sinealba and its relatives [J]. Guihaia, 38(5): 635-645. [李晓娟, 李建秀, 徐宁, 等, 2018. 中华白花丹参与其近缘种的形态分类鉴定 [J]. 广西植物, 38(5): 635-645.]
LI XL, CHEN L, 2012. Breeding for triploids of Salvia miltiorrhiza and its sustainable utilization [J]. Chin Trad Herb Drugs, 43(2): 375-379. [李秀兰, 陈力, 2012. 三倍体丹参的培育及其可持续利用研究 [J]. 中草药, 43(2): 375-379.]
LIU WG, WANG M, YAN ZH, 2003. Observation and comparison on pollen morphology of different ploidy watermelon [J]. Acta Hortic Sin, 30(3): 328-330. [刘文革, 王鸣, 阎志红, 2003.不同倍性西瓜花粉形态观察 [J]. 园艺学报, 30(3): 328-330.]
MARASEK-CIOLAKOWSKA A, XIE SL, ARENS P, et al., 2014.Ploidy manipulation and introgression breeding in Darwin hybrid tulips [J]. Euphytica, 198(3): 389-400.
SALMA U, KUNDU S, MANDAL N, 2017. Artificial polyploidy in medicinal plants: Advancement in the last two decades and impending prospects [J]. J Crop Sci Biotechnol, 20(1): 9-19.
SHI YC, WEI YJ, ZOU R, et al., 2020. Observation on pollen morphology of main varieties of Sophora japonica ‘Jinhuai’ and S. japonica ‘Mihuai’ [J]. Guihaia, 40(1): 24-30. [史艳财, 韦宇静, 邹蓉, 等, 2020. 金槐和米槐主流品种花粉形态研究 [J]. 广西植物, 40(1): 24-30.]
WANG KF, WANG XZ, 1983. Introduction to palynology [M]. Beijing: Peking University Press. [王开发, 王宪曾, 1983. 孢粉学概论 [M]. 北京: 北京大学出版社.]
WANG SD, WANG XC, LIU JS, et al., 2015. Pollen morphology and germination characteristics of peach triploid germplasms [J]. N Hortic, (12): 80-84. [王尚德, 王晓川, 刘佳棽, 等, 2015. 桃三倍体种质的花粉形态及萌发特性 [J]. 北方园艺, (12): 80-84.]
XU RS, 1990. Salvia miltiorrhiza: Biology and utilization [M]. Beijing: Science Press. [徐任生, 1990. 丹参: 生物学及其应用 [M]. 北京: 科学出版社.]
YANG DK, SUN JT, WANG BQ, 2003. Pollen morphology and taxnomic significance of Salvia from Shandong [J]. Shandong Sci, 16(1): 14-16. [楊德奎, 孙京田, 王丙全, 2003. 山东鼠尾草属花粉形态的研究及其在分类上的意义 [J]. 山东科学, 16(1): 14-16.]
ZHANG NX, ZHANG LY, CAO YN, et al., 2019. Cytological analysis on fertility of triploid Salvia miltiorrhiza [J]. Chin J Trop Crop, 40(8): 1546-1550. [张铌璇, 张璐瑶, 曹艳楠, 等, 2019. 丹参三倍体育性的细胞学分析 [J]. 热带作物学报, 40(8): 1546-1550.]
ZHANG YF, HU CQ, YANG Y, et al., 2016. Pollen morphology observation of eight resources in Diospyros [J]. Acta Hortic Sin, 43(6): 1167-1174. [张永芳, 胡超琼, 杨勇, 等, 2016. 柿属8种植物花粉形态观察 [J]. 园艺学报, 43(6): 1167-1174.]
(责任编辑 周翠鸣)
