新生杜藏和杜滇仔猪抗逆能力比较研究
2021-01-07刘韶娜赵智勇赵素梅赵彦光
郭 飞 刘韶娜 张 斌 方 晨 赵智勇 沙 茜 赵素梅 赵彦光*
(1.云南农业大学动物营养与饲料科学重点实验室,云南昆明650201;2.云南省畜牧兽医科学院,云南昆明650224)
我国地方猪种具有抗逆性强、肉质优良等特点,不同地方猪种抗逆能力也有差异,主要表现在耐寒、耐热、耐粗饲、对饥饿的耐受力、抗病力等方面[1]。目前对于地方猪的抗逆能力描述多是概念性的,未见相应量化研究报道。迪庆藏猪和滇南小耳猪均为云南特有的地方猪种资源,且具有较强抗逆能力,可作为抗逆性研究理想模型。在猪的生长阶段中,哺乳期因消化器官不发达、消化机能不完善、缺乏先天免疫力等因素,导致其体温调节能力差,仔猪腹泻率、死亡率明显高于其他生理阶段,抵抗疾病能力较差[2],此阶段可作为验证抗逆特性最好的生理阶段。为探讨猪的抗逆能力的品种差异,以及通过营养手段进行干预的可行性,本试验人为设置了一些应激条件,通过饲养试验,测定哺乳仔猪的平均日采食量、体增重、腹泻频率和死亡率,初步对猪的抗逆能力进行量化比较研究,试图挖掘地方猪种的抗逆潜力,并探究营养等手段干预猪抗逆能力的可行性,从而为猪抗逆性的相关研究提供一定理论基础。
1 材料与方法
1.1 试验动物
以杜洛克为父本,滇南小耳猪和迪庆藏猪分别为母本进行杂交,根据血缘、体重相近(0.9±0.23)kg、公母各半原则,从产期相近的12窝杂交F1代(滇南小耳猪和迪庆藏猪血缘各占50%,杜洛克血缘占50%,分别简称杜滇和杜藏)发育正常且健康的初生仔猪中,每窝选取4头,共48头为试验动物。
1.2 高能低蛋白代乳粉日粮设计
参照肉脂型地方猪5~8 kg 饲养标准(NY/T 65—2004)设计高能低蛋白代乳粉日粮配方,配方组成及营养水平见表1,采购优质原料,按相关标准配制代乳粉。所需添加的复合益生菌由云南畜牧兽医科学院养猪与动物营养研究所提供,详见表2。
1.3 试验设计
试验采用双因素交叉试验设计,将仔猪分为3组,设置母乳饲喂(母乳组)和人工饲喂(代乳粉组)、益生菌组,每个处理的杜藏、杜滇仔猪各8 头,公母各半,具体分组见表3。人工饲喂的仔猪采取群养单饲。试验于仔猪5 日龄断奶后进行分组预饲,7 日龄开始正式试验,28日龄结束,试验期22 d。

表1 代乳粉组成和营养水平(风干基础)

表2 益生菌组所用益生菌的添加量
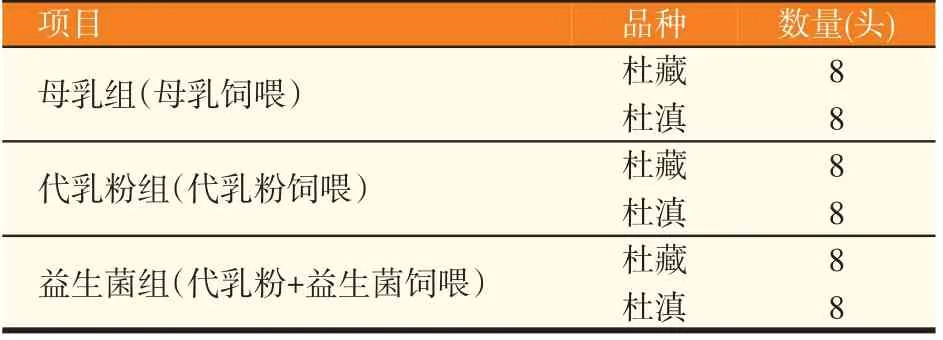
表3 试验动物分组情况
1.4 饲养管理
试验仔猪饲养于同一猪舍,舍内设有保温灯,平均温度保持在27~32 ℃之间。母乳组通过寄养的方式由母猪哺育(母猪哺育能力为8~10头)。人工饲喂需要对代乳粉进行调制。代乳粉的调制方法:将代乳粉与50~65 ℃的温开水按1∶6 的比例冲调配制成代乳粉液,冷却至40 ℃左右饲喂,益生菌组在每天饲喂前添加益生菌。代乳粉组和益生菌组的仔猪5 日龄断奶后进行群养单饲,配好代乳粉,利用猪用奶瓶单头进行人工饲喂。5~7日龄进行预饲和教食,少量多喂并观察每头仔猪的采食情况,7 日龄时试验正式开始。试验的第1~2 d每日每头次饲喂6次,第3~4 d饲喂5 次,第5 d 开始每天每头次饲喂4 次至21 日龄,21 日龄后减少至每天3 次,每次饲喂仔猪自由采食,记录每头仔猪代乳粉的日采食量。从21 日龄开始,对3个组的仔猪定量补伺少许乳猪料。试验期间,除饲喂方式不同外,其余饲养管理方式均相同。
1.5 指标测定与方法
1.5.1 采食量
对照组因母猪哺育,无法收集母乳采食量数据。试验组每头仔猪代乳粉采食量依据公式(1)计算,并按照代乳粉∶水为1∶6的配制比例折算为代乳粉采食量。

1.5.2 增重
分别在7、21、28 日龄早晨8 点进行称重,记录每头仔猪体重,按阶段计算仔猪体重变化。
1.5.3 腹泻指标
每天观察试验仔猪的健康情况,统计各试验阶段的腹泻仔猪头次数,作为仔猪腹泻频率。同时记录仔猪死亡情况,计算死淘率,具体见公式(2)。

1.5.4 数据处理
试验数据用Exel 2010 统计,结果以”平均数±标准误“表示。所有数据利用SPSS 22.0 进行正态性检验,符合正态分布的数据采用General Linear Model进行分析,并以LSD 进行多重比较,非正态分布数据采用Kruskal-Wallis 多样本秩和检验进行显著性分析,P<0.05视为差异显著。腹泻频率为次数资料,利用卡方检验进行显著性检验。
2 试验结果与分析
2.1 品种与饲喂方式对新生仔猪平均日采食量的影响(见表4)
表4所示,品种对7~20日龄仔猪平均日采食量无显著影响(P>0.05),对21~28、7~28 日龄仔猪平均日采食量具有显著影响(P<0.05);饲喂方式对7~20、21~28 日龄和7~28 日龄仔猪平均日采食量影响显著(P<0.05),益生菌组与代乳粉组相比,7~20、21~28日龄及7~28 日龄的仔猪平均日采食量分别提高了7.64、14.10、10.36 g;品种与饲喂方式互作对不同日龄仔猪平均日采食量均有显著影响(P<0.05)。

表4 品种与饲喂方式对新生仔猪平均日采食量的影响(g)
2.2 品种与饲喂方式对新生仔猪体增重的影响(见表5)
如表5 所示,在7~20、21~28、7~28 日龄各阶段,杜藏仔猪的体增重均高于杜滇仔猪,但差异不显著(P>0.05);饲喂方式显著影响仔猪体增重(P<0.05),母乳组仔猪体增重显著高于代乳粉组和益生菌组(P<0.05),代乳粉组与益生菌组7~20 日龄仔猪的体增重呈负增长,但益生菌组的负增长值小于代乳粉组。
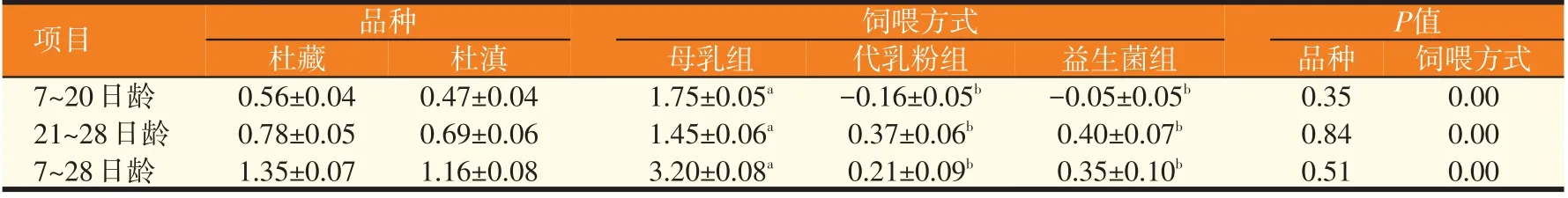
表5 品种与饲喂方式对新生仔猪体增重的影响(kg)
2.3 品种与饲喂方式对新生仔猪腹泻及死淘率的影响(见表6)
如表6 所示,仔猪的腹泻频率与品种显著相关(P<0.05),在7~20、21~28、7~28 日龄阶段的杜滇仔猪腹泻频率均高于杜藏仔猪,7~20 日龄时杜滇仔猪和杜藏仔猪腹泻最严重,杜滇仔猪较杜藏仔猪高13.7 个百分点;杜滇仔猪的死淘率为25%,杜藏仔猪无死亡。仔猪的腹泻频率与饲养方式显著相关(P<0.05),7~20、21~28、7~28 日龄仔猪的腹泻频率表现为代乳粉组>益生菌组>母乳组;代乳粉组与益生菌组的死淘率均为12.5%,母乳组无仔猪死亡。

表6 品种与饲喂方式对新生仔猪腹泻及死淘率的影响
3 讨论
3.1 品种对仔猪抗逆能力的影响
滇南小耳猪与迪庆藏猪是云南特有的地方猪种,前者主要分布在热带、亚热带低海拔地区,具有耐高温潮湿、抗逆性强的种质特性[3];后者主要分布在寒冷、干燥、低氧的高原地区,长期以来形成了四肢结实、抗逆性强、抗病力强、耐粗饲等适应高原生态环境的种质特性[4]。本研究发现,含有50%迪庆藏猪血缘的杜藏仔猪对人为设置的应激条件的抵抗能力比具有50%滇南小耳猪血缘的杜滇仔猪强,不同日龄阶段杜藏仔猪平均日采食量、体增重、腹泻频率、死淘率均优于杜滇仔猪,表现出更强的抗逆性,这可能与作为母本的迪庆藏猪、滇南小耳猪的影响息息相关。线粒体遗传遵循严格的母系遗传,当前许多动物对恶劣环境的适应性研究也是从线粒体基因组角度入手[5],线粒体微卫星(mitochondrial microsatellite, mtMs)丰富的变异和特有结构与动物的长期适应性进化有关,有研究发现,与多个猪种相比,藏猪的mtMs复合结构多达8 个,且RARCRE为藏猪特有mtMs,藏猪在长期进化中可能拥有适应高海拔和高寒环境的mtDNA 遗传基础[6]。此外,有研究表明,母体乳汁、肠道、产道、粪便等母源微生物会通过垂直传递影响新生仔猪肠道黏膜微生物的定植[7],白浩男[8]通过藏猪盲肠内容物的16S rDNA高通量测序发现,相比于其他猪种,藏猪的拟杆菌门、螺旋体门细菌物种相对丰度较高而厚壁菌门、肠杆菌科细菌较低,这一独特的菌群结构可能与藏猪草食、抗病、抗逆的种质特性密切相关;李卓君[9]通过藏猪肠道菌群功能预测分析表明,藏猪群体特有的微生物或许与藏猪的抗逆、抗病和耐粗等种质特性相关。总之,作为母本的迪庆藏猪可能通过线粒体遗传、母源微生物的垂直传递等方式影响杜藏仔猪的抗逆能力,但具体机制还需进一步研究。
3.2 不同饲喂方式对仔猪抗逆能力的影响
有研究指出,在日粮中添加的益生菌可通过竞争抑制作用、刺激宿主免疫系统提高宿主免疫功能,通过有机酸、多种消化酶及生物活性物质的产生促进宿主的营养消化吸收,通过改善肠道微生态环境促进宿主的肠道健康[10]。本研究中的益生菌组仔猪日粮是在代乳粉组配方的基础上添加了益生菌,在不同日龄阶段,益生菌组平均日采食量显著高于代乳粉组,腹泻频率较代乳粉组低,表明仔猪饲粮中添加益生菌可提高仔猪采食量,降低腹泻频率,改善仔猪抗逆能力。曾秀玲等[11]、陈宝剑等[12]研究也指出益生菌对仔猪的生长具有显著的益生作用。此外,本研究中人为设置的应激因素包括两个方面,一方面是仔猪过早断奶产生的断奶应激,另一方面是代乳粉取代母乳的营养应激。无论是断奶应激和营养应激均会对仔猪造成诸多不利影响[13-14]。本研究发现,与母乳组相比,在仔猪早期阶段代乳粉组和益生菌组的仔猪体增重呈负值,但益生菌组的负增长值小于代乳粉组,体增重呈负值可能是由于仔猪需要适应试验中人为设置的应激因素,但益生菌的添加可减缓应激对仔猪的影响。总之,添加益生菌可以改善仔猪的抗逆能力,这也意味着通过营养手段可以干预猪抗逆能力。
4 结论
杜藏仔猪抗逆能力强于杜滇仔猪,这可能与母系的品种差异有关;添加益生菌提高了仔猪的采食量,降低了仔猪腹泻频率,改善了仔猪抗逆能力,表明通过营养手段干预仔猪抗逆能力的可行性。
