Musashi-1和Wnt/β-catenin信号通路与卵巢子宫内膜异位症的相关性研究
2021-01-05于聪祥李跃飞
于聪祥 ,李跃飞
(1.内蒙古医科大学附属医院妇产科,内蒙古 呼和浩特;2.内蒙古自治区妇幼保健院妇产科,内蒙古 呼和浩特)
0 引言
卵巢子宫内膜异位症( ovarian endometriosis) 主要发生于女性生殖系统,发病率呈逐年上升趋势[1]。虽为良性疾病,但具有无限生长、侵袭及远处转移等恶性肿瘤的特性,故有“良性癌”之称。目前对于卵巢子宫内膜异位症发病机制尚未清晰。细胞凋亡(apoptosis)又称为细胞程序性死亡,机体通过调节细胞数量,进而维持组织细胞的动态平衡,是维持细胞稳态的重要方式之一。细胞凋亡异常与多种肿瘤的发生密切相关[2-3]。Mushasi-1及Wnt/β-catenin信号转导通路通过抑制细胞凋亡促进癌细胞增殖是近年来的研究热点[4]。因此,本研究通过检测Mushasi-1、β-catenin在卵巢子宫内膜异位症患者月经第一天脱落子宫内膜及未脱落子宫内膜的表达情况,探讨细胞凋亡与卵巢子宫内膜异位症的相关性。
1 对象与方法
1.1 对象
采集2012年5月至2015年9月内蒙古医科大学附属医院患者标本,实验组为:病理证实卵巢子宫内膜异位症志愿者子宫内膜标本36份,收集经血及使用内膜取样器各获得18份。对照组为:选择因输卵管因素不孕或宫颈病变住院手术志愿者的子宫内膜标本40份,收集经血及使用内膜取样器各获得20份。标本采集得到了内蒙古医科大学附属医院医学伦理委员会同意,并取得了患者本人授权。
入选标准:①年龄20-30岁;②无子宫肌瘤,无生殖器结核及恶性肿瘤病史,无剖宫产及子宫肌瘤剔除术史;③术前3个月未用激素类药物,未用抗生素及GnRHa;④未放置宫内节育器;⑤无高血压、糖尿病、肾病等病史;⑥病理证实无子宫内膜病变。
1.2 方法
1.2.1 标本采集及处理
术前一个月经周期收集标本,于月经第一天经血量较多时收集阴道内的经血获取脱落子宫内膜,常规消毒阴道及宫颈,使用内膜取样器轻轻刮取,获得未脱落子宫内膜,并分别用4℃生理盐水迅速冲洗,获得内膜组织标本,迅速置于无菌无酶的冻存管、液氮罐中保存,用于Real-Time PCR实验。
1.2.2 Real-Time PCR实验
Trizol、逆转录和Real-Time PCR试剂盒购于北京天根公司,引物购于上海生工。Trizol法提取组织Total RNA,通过琼脂糖电泳检测符合Real-Time PCR实验的要求。引物序列:Musashi-1上游5’-ATTTCCCCTCCTTTACCCTTTC-3’,下游 5’-GGGCTTCACATTCACAAACCTA-3’;β-catenin 上游 5’-TGACCTGAGACTGGATGTAGAAA-3’,下游 5’-GCTGGAATGACAACTGGA TAAAC-3’;β-actin上游 5’-TGACGTGGACATCCGCAAAG-3’,下游5’-CTGGAAGGTGGACAGCGAGG-3’,逆转录合成cDNA:42℃ 50min,95℃ 5min,PCR 扩增 cDNA:95℃ 15 min预变性,PCR 反应 95℃ 10sec,60℃ 20sec,72℃ 31sec,45Cyele。融解曲线分析 95℃ 10 sec,65℃ 15 sec,95℃ 10sec。ABI7300荧光定量PCR仪测定反应终产物,采取相对定量法,参照β-actin对各组目的基因表达的强度进行校正(△Ct = Ct目的基因- Ctβ-actin基因)。相对表达量差值,即2-△△Ct表示各组Musashi-1、β-catenin的表达差异。
表1 实验组、对照组Musashi-1/β-actin和β-catenin /β-actin的相对表达量比较()

表1 实验组、对照组Musashi-1/β-actin和β-catenin /β-actin的相对表达量比较()
组别 例数(n) 2-△△Ct Musashi-1β-catenin对照组(40例) 脱落内膜 20未脱落内膜 20 3.10 33.36实验组(36例) 脱落内膜 18 9.51 3.07 107.63 3.23未脱落内膜 18 11.31 3.66 1.19 164.28 4.92 1.53 F值 59.450 433.975 P值 0.0000 0.0000
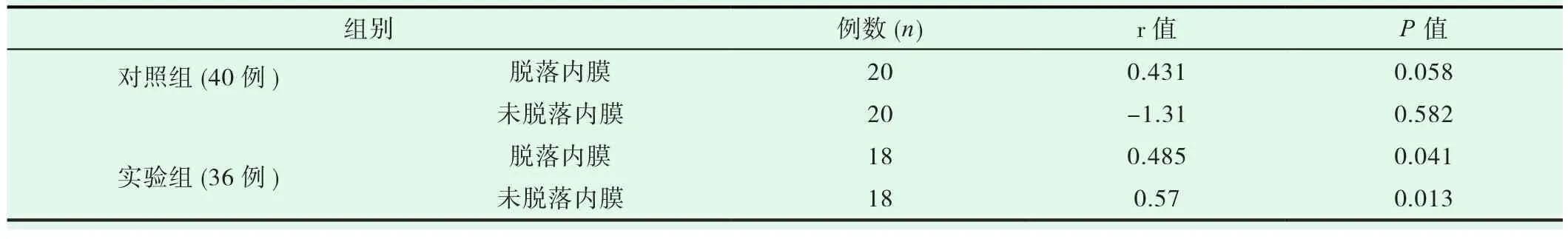
表2 实验组、对照组Musashi-1/β-actin和β-catenin /β-actin相对表达量的相关性
1.3 统计学处理
2 结果
2.1 Musashi-1相对表达量的方差分析
Musashi-1在实验组未脱落内膜的相对表达量较脱落内膜高1.19倍,而且比对照组脱落内膜及对照组未脱落内膜分别高11.31倍及3.66倍。Musashi-1在实验组脱落内膜的相对表达量比对照组脱落内膜及对照组未脱落内膜分别高9.51倍和3.07倍。Musashi-1在对照组未脱落内膜的相对表达量较脱落内膜高3.10倍。单因素方差分析结果显示,Musashi-1在实验组未脱落内膜的相对表达量高于对照组脱落内膜及对照组未脱落内膜,差异有统计学意义(Mean Diference=-3.4901,P=0.0000;Mean Diference=-1.8561,P=0.0000);并且Musashi-1在实验组脱落内膜的相对表达量也较对照组脱落内膜及对照组未脱落内膜高,差异均有 统 计 学 意 义 (Mean Diference=-3.2479,P=0.0000;Mean Diference=-1.6139,P=0.0000)。Musashi-1在实验组脱落内膜仍高表达,与实验组未脱落内膜组相比,差异无统计学意义 (Mean Diference=0.2422,P=0.4322)。 而 Musashi-1 在 对照组脱落内膜的相对表达量低于对照组未脱落内膜,差异具有统计学意义(Mean Diference=1.6340,P=0.0000)(见表1)。
2.2 β-catenin相对表达量的方差分析
β-catenin在实验组未脱落内膜的相对表达量比脱落内膜高1.53倍,比对照组脱落内膜及对照组未脱落内膜分别高164.28倍和4.92倍。β-catenin在实验组脱落内膜的相对表达量比对照组脱落内膜及对照组未脱落内膜分别高107.63倍和3.23倍。β-catenin在对照组未脱落内膜的相对表达量比脱落内膜高33.36倍。单因素方差分析结果显示,β-catenin在实验组未脱落内膜的相对表达量较其脱落内膜和对照组脱落内膜、对照组未脱落内膜均高,差异均具有 统 计 学 意 义 (Mean Diference=-0.6111,P=0.0114;Mean Diference=-7.3625,P=0.0000;Mean Diference=-2.3050,P=0.0000)。β-catenin在实验组脱落内膜的相对表达量较对照组脱落内膜及对照组未脱落内膜高,差异具有统 计 学 意 义 (Mean Diference=-6.7514,P=0.0000;Mean Diference=-1.6939,P=0.0000)。β-catenin在对照组未脱落内膜的相对表达量较脱落内膜高,差异有统计学意义(Mean Diference=-5.058,P=0.0113)(见表 1)。
2.3 Musashi-1与β-catenin相对表达量的Pearson直线相关分析
Musashi-1与β-catenin在实验组未脱落内膜及脱落内膜组中呈正相关,但对照组未脱落内膜及脱落内膜中二者无相关 (见表 2)。
3 讨论
卵巢子宫内膜异位症目前有“在位内膜决定论”、“经血逆流学说”、免疫等学说,却尚无一理论可解释其发病机制。随着分子生物学的发展,卵巢子宫内膜异位症发病机制的研究也已深入至分子水平。细胞凋亡是机体发育、动态平衡及免疫调节的重要过程,受多种基因调控的细胞自主且有序地程序性死亡[5]。月经增殖期时,增殖的新生细胞可以弥补损失的衰老细胞;然而月经期时,子宫内膜功能层却通过细胞凋亡机制清除衰老细胞。然而,研究发现子宫内膜异位症患者细胞凋亡比例明显降低,但上皮细胞活性增强[6]。
Musashi-1高表达于肿瘤组织,且具有促进肿瘤细胞生长及抑制肿瘤细胞凋亡的功能[7]。研究发现Musashi-1在恶性肿瘤组织的表达率明显高于良性病变组织,敲除Musashi-1 RNA结合蛋白后,发现肿瘤生长停滞,且细胞凋亡增加,增殖降低[8-9]。本研究结果显示,Musashi-1在实验组未脱落内膜的相对表达量高于对照组脱落内膜及对照组未脱落内膜,差异具有统计学意义(P=0.0000,P=0.0000),Musashi-1在实验组脱落内膜的相对表达量也较对照组脱落内膜、对照组未脱落内膜高,差异具有统计学意义(P=0.0000,P=0.0000)。提示卵巢子宫内膜异位症患者未脱落子宫内膜、脱落子宫内膜组织比正常对照组中细胞凋亡要明显减少,这意味着卵巢子宫内膜异位症患者未脱落子宫内膜、脱落子宫内膜组织很可能仍具有较强的增殖能力,与Zhao[10]提出的细胞凋亡异常参与了子宫内膜异位症的结论相符。张连锁[11]等研究发现下调程序性细胞死亡因子5(programmed cell death 5,PDCD5)的表达,可以引起细胞凋亡减少,生存能力增强,促使异位的内膜细胞种植生长,进而形成卵巢子宫内膜异位症。本研究实验结果提示,Musashi-1在实验组脱落内膜仍高表达,与实验组未脱落内膜相比差异无统计学意义(P=0.4322)。而Musashi-1在对照组脱落内膜的相对表达量低于对照组未脱落内膜,差异具有统计学意义(P=0.0000)。从而进一步论证了正常对照组子宫内膜组织发生了细胞凋亡,而卵巢子宫内膜异位症患者的未脱落子宫内膜、脱落子宫内膜组织细胞过度存活,可能仍然具有较强的增殖能力,如果异位于适当的肿瘤微环境便可能形成子宫内膜异位病灶。提示抑制Musashi-1的表达可能是卵巢子宫内膜异位症基因治疗的手段之一。
Wnt/β-catenin信号转导通路对细胞凋亡的调控作用与肿瘤细胞的增殖相关[12]。β-catenin作为 Wnt/β-catenin信号转导通路的关键信号因子,在肿瘤的发生发展过程中起重要调控作用,通过抑制细胞凋亡过程,激活下游促增殖蛋白转录从而促进肿瘤细胞的增殖、存活及转移[13]。本研究结果显示β-catenin在实验组未脱落内膜的相对表达量高于对照组脱落内膜及对照组未脱落内膜,差异具有统计学意义 (P=0.0114,P=0.0000,P=0.0000)。β-catenin 在实验组脱落内膜的相对表达量高于对照组脱落内膜及对照组未脱落内膜,差异具有统计学意义(P=0.0000,P=0.0000)。β-catenin在对照组未脱落内膜的相对表达量高于脱落内膜,差异有统计学意义。意味着正常对照组的子宫内膜组织发生了细胞凋亡,属于子宫内膜周期性变化,而卵巢子宫内膜异位症患者,脱落子宫内膜和未脱落子宫内膜组织凋亡异常,细胞凋亡明显减低,意味着卵巢子宫内膜异位症患者脱落子宫内膜和未脱落子宫内膜组织可能仍具有较强的增殖分化活性。提示Wnt/β-catenin信号转导通路的激活可能抑制了子宫内膜细胞凋亡的发生,并参与卵巢子宫内膜异位症的发生及发展。学者提出Wnt/β-catenin信号转导通路是药物干预治疗胃癌的重要靶点[13]。因此,推测抑制Wnt/β-catenin信号转导通路可能是治疗卵巢子宫内膜异位症的潜在靶点。
Musashi-1主要通过调节 Wnt/β-catenin、Notch等信号转导通路参与肿瘤的发生发展进程[12]。Wang[14]等研究证实Musashi-1可以通过激活Wnt/β-catenin信号转导通路促进乳腺癌的发生发展。赵明明[15]等提出Wnt/β-catenin信号转导通路一旦被激活后,便可抑制星形胶质细胞发生细胞凋亡。本研究发现Musashi-1和β-catenin在实验组未脱落内膜、脱落内膜组中呈正相关性,而对照组未脱落内膜及脱落内膜中二者无相关性。提示Musashi-1有可能通过激活Wnt/β-catenin信号转导通路及其下游靶基因,抑制细胞凋亡,从而增强组织细胞的增殖分化侵袭能力,参与卵巢子宫内膜异位症的形成。通过调节Wnt/β-catenin信号转导通路,可能是卵巢子宫内膜异位症基因治疗的手段,有望成为卵巢子宫内膜异位症新的治疗途径。
综上所述,Musashi-1可能通过激活Wnt/β-catenin信号转导通路参与抑制子宫内膜细胞发生凋亡,从而增强组织细胞的增殖分化侵袭能力,参与卵巢子宫内膜异位症的形成。但仍存在不足之处:β-catenin通过激活了哪些下游靶基因从而降低卵巢子宫内膜异位症患者脱落子宫内膜和未脱落子宫内膜组织凋亡的,还有待课题组进一步实验揭示。课题组将进一步扩大样本量,体外培养卵巢子宫内膜异位症细胞,通过调节Musashi-1及Wnt/β-catenin信号转导通路的相关上游及下游靶基因,进一步探讨其致病机制,为临床治疗提供理论依据。
