长白山岳桦林与暗针叶林交错区植被群落结构特征分析
2021-01-05刘丽杰
贾 翔,王 超,曹 淋,赵 莹,金 慧,刘丽杰,尹 航
(1.长白山科学研究院,吉林 二道白河 133613;2.长白山生物群落与生物多样性吉林省联合重点实验室,吉林 二道白河 133613;3.吉林省长白山自然保护管理中心,吉林 二道白河 133613)
群落交错区(ecotone)又称生态交错区或生态过渡带,是两个或多个群落间的过渡区域[1].通常认为,群落交错区的环境条件比较复杂,是一个交叉地带或种群竞争的紧张地带,所以交错带群落结构比较脆弱,容易受到外界干扰而发生变化.目前,群落交错区是生态学的研究热点[2],受到了广泛关注[3].
长白山是东北地区的第一高山,气候条件在山体垂直高度上的差异,导致长白山成为东北植物区系唯一一座有明显植被垂直分布带的名山[4],自山麓至山顶依次为针阔混交林带、暗针叶林带、岳桦林带、高山苔原带.长白山不仅保存了完整的垂直植被带类型,而且拥有森林、湿地等多种生态系统类型,所以该地区具有众多种类的群落交错带,是研究群落交错区的理想之地.21世纪初,于大炮等[5]对长白山垂直分布带交错区的位置及其宽度进行了定量判定,明确提出长白山北坡暗针叶林和岳桦交错区为急变类型,宽度为50 m;邹春静等[2]的研究表明,长白山北坡岳桦种群在海拔1 600~1 700 m附近与云冷杉林形成森林过渡带;后期逐渐开展了群落交错区植被变化与演替规律[6-7]等研究.然而,以往对长白山交错区植被群落结构的研究只侧重于一个坡向.因此,本研究分别调查长白山北坡、西坡岳桦林与暗针叶林过渡带的植被群落,旨在了解该过渡带在不同坡向的植被群落结构特征,为该区植被恢复研究提供依据.
1 研究区概况及研究方法
1.1 研究区自然概况
长白山岳桦林与暗针叶林交错区位于长白山国家级自然保护区内,宽度较为狭窄,分布于海拔约1 700 m处.该区域气候属湿润性亚高山气候,冬季寒冷多风,夏季湿润多雨,年平均气温较低,生长季较短.交错区以上为岳桦林带,乔木以岳桦(Betulaermanii)为主,土壤为山地生草森林土;交错区以下为暗针叶林带,乔木以鱼鳞云杉(Piceajezoensisvar.microsperma)、红皮云杉(Piceakoraiensis)、臭冷杉(Abiesnephrolepis)为主,土壤为棕色针叶林土[8].
1.2 研究方法
1.2.1 野外调查
分别在长白山西坡(41°57′N,127°59′E,海拔1 680 m)和北坡(42°04′N,128°04′E,海拔1 700 m)设置30 m×30 m的调查样地,各设置3块,共6块.将样方划分为9个10 m×10 m的小样方进行乔木层调查;按对角线方向设置3个样方进行灌木层和草本层调查,灌木样方为5 m×5 m,草本样方为2 m×2 m.调查项目包括乔木层所有胸径≥2 cm的树种名称、胸径,灌木层的植物名称、基径和草本层的植物名称、株数、盖度等植被信息.
1.2.2 数据分析
群落各层重要值计算公式:乔木层重要值=((相对多度+相对频度+相对优势度)×100)/3,灌木层重要值=((相对多度+相对频度+相对基盖度)×100)/3,草本层重要值=((相对多度+相对频度+相对盖度)×100)/3.
α多样性测定主要包括物种丰富度(R)、Shannon-Wiener指数(H)、Simpson指数(D)和Pielou均匀度指数(J).
1)物种丰富度指数:
R=S,
式中:S为样方中的总物种数.
2)Shannon-Wiener指数:
式中:Ni为种i的个体数;N为所在群落全部种的个体总数.
3)Simpson指数:
4)Pielou均匀度指数:
1.2.3 数据统计与分析
利用Excel 2007和SPSS 19.0处理数据及进行统计分析,采用t检验法检验两个变量之间的差异显著性(α=0.05),利用Origin 8.5绘图.
2 结 果
2.1 物种组成
2.1.1 乔木层
长白山岳桦林与暗针叶林交错区乔木层物种组成见表1.结果显示:在长白山西坡共调查到3种乔木,总计101株,主要由岳桦和鱼鳞云杉等组成.其中,岳桦的重要值最大,为69.24,株密度为256株/hm2,胸高断面积为12.78 m2/hm2;鱼鳞云杉的重要值为29.77,株密度为115株/hm2,胸高断面积为4.45 m2/hm2.在长白山北坡共调查到6种乔木,总计500株,主要由鱼鳞云杉、臭冷杉、岳桦等组成.其中,鱼鳞云杉的重要值最大,为41.47,株密度为719株/hm2,胸高断面积为35.84 m2/hm2;臭冷杉的重要值为26.34,株密度为633株/hm2,胸高断面积为12.09 m2/hm2,二者均为长白山暗针叶林的典型优势树种,重要值之和接近70;岳桦重要值仅为15.24,株密度为226株/hm2,胸高断面积为8.25 m2/hm2.
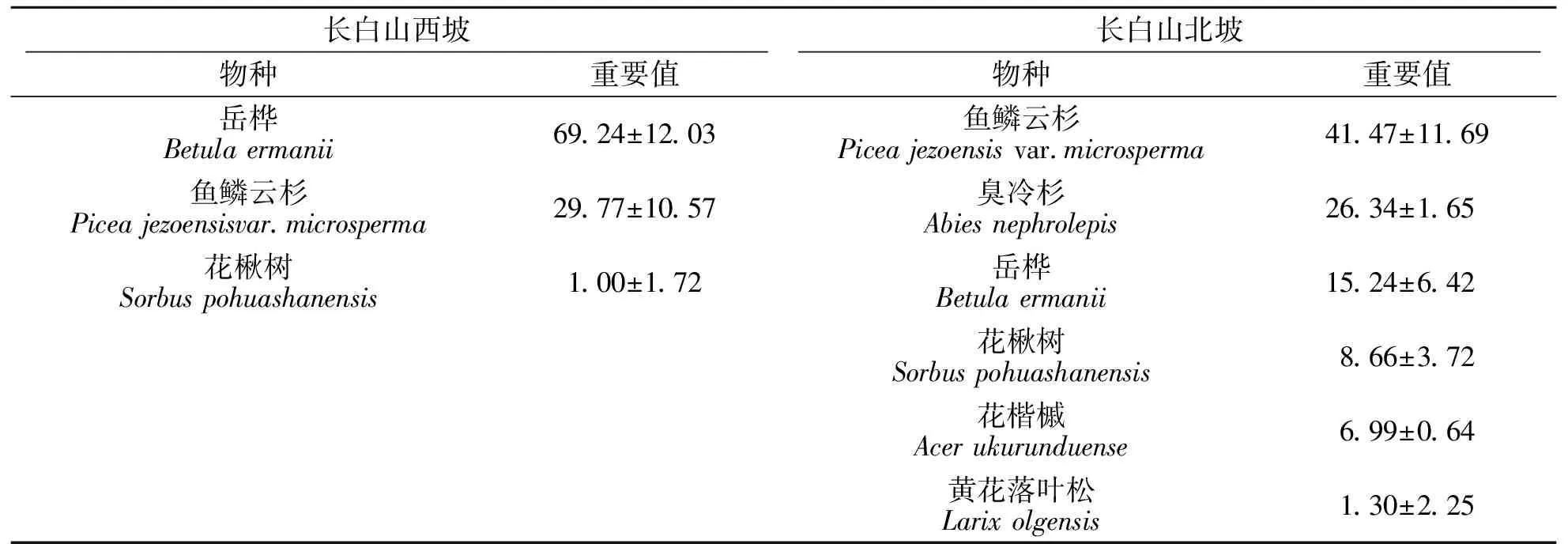
表1 岳桦林与暗针叶林交错区乔木层物种组成Tab.1 Species composition in the arbor layer of Betula ermanii and dark coniferous forest ecotone
2.1.2 灌木层
长白山岳桦林与暗针叶林交错区灌木层物种组成见表2.结果显示:在长白山西坡共调查到3种灌木,分别为蓝靛果忍冬(Loniceracaeruleavar.enulis)、库页悬钩子(Rubussachalinensis)和深山蔷薇(Rosamarretii),共计64株.其中,蓝靛果忍冬的重要值最大,为51.43,个体数为21株;库页悬钩子的重要值为31.98,个体数为37株.在长白山北坡共调查到7种灌木,主要由刺腺茶藨子(Ribeshorridum)、长白忍冬(Loniceraruprechtiana)、蓝靛果忍冬等组成,共计1 693株.其中,刺腺茶藨子的重要值最大,为36.24,个体数为938株;长白忍冬的重要值为27.73,个体数为500株.

表2 岳桦林与暗针叶林交错区灌木层物种组成Tab.2 Species composition in the shrub layer of Betula ermanii and dark coniferous forest ecotone
2.1.3 草本层
长白山岳桦林与暗针叶林交错区的草本层重要值排名前5位的物种见表3.结果显示:在长白山西坡共调查到22种草本植物,主要为小叶章(Calamagrostisangustifolia),其个体数占总个体数的85.97%,重要值达到61.07,为典型的小叶章群落.在长白山北坡共调查到36种草本植物,主要由二叶舞鹤草(Maianthemumbifolium)、杉蔓石松(Lycopodiumannotinnm)、广布鳞毛蕨(Dryopterisexpansa)、高山露珠草(Circaeaalpina)等组成.其中,二叶舞鹤草的重要值最大,为22.99;其次是杉蔓石松,重要值为11.71;广布鳞毛蕨的重要值为10.15.
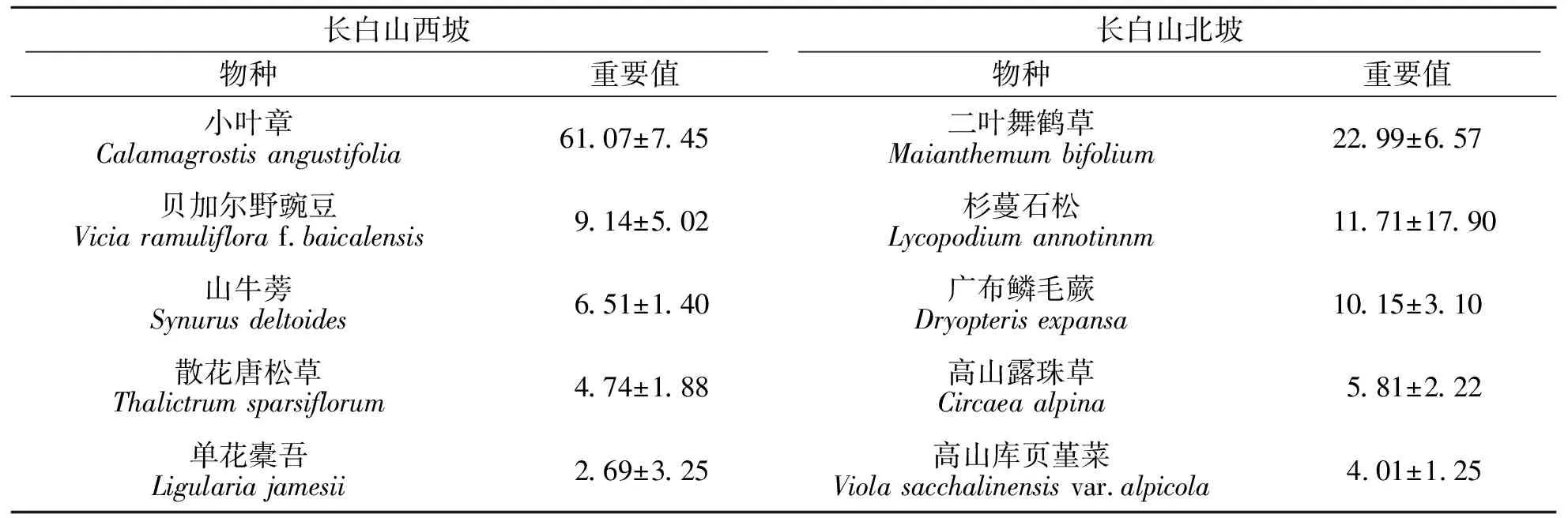
表3 岳桦林与暗针叶林交错区草本层重要值排名前5位的物种Tab.3 Important values of species ranked in the top 5 in the herbaceous layer of Betula ermanii and
2.2 多样性指数
长白山岳桦林与暗针叶林交错区的α多样性指数见图1(不同字母表示差异显著).
由图1可知:长白山北坡乔木层物种丰富度、Shannon-Wiener多样性指数和Simpson优势度指数均显著高于西坡,Pielou均匀度指数虽然高于西坡,但不显著;灌木层物种丰富度、Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数均高于西坡,但都不显著;草本层物种丰富度、Shannon-Wiener多样性指数和Simpson优势度指数、Pielou均匀度指数均显著高于西坡.
2.3 乔木径级结构
长白山岳桦林与暗针叶林交错区的乔木径级分布见图2.由图2可知:长白山西坡乔木径级结构分布无明显规律,但近年来乔木更新良好.调查发现,幼树(DBH<12 cm)个体数相对较多,占全部个体数的31.68%,群落呈现增加趋势;如果单独考虑DBH≥14 cm的乔木,则径级分布总体呈现正态分布;长白山北坡样地乔木径级结构总体呈现明显的倒“J”形分布.
为了更加清楚地了解各群落的径级结构特征,进一步对群落乔木层重要值较大的树种进行径级结构分析,结果见图3.由图3可见:西坡岳桦的小径级和中等径级个体在样地内分布较多,其中DBH<4 cm和18 cm≤DBH<32 cm的个体数占总个体数的15.87%和62.32%;鱼鳞云杉较大径级个体零星分布于样地内,小径级个体较多,其中,DBH<12 cm的个体数占总个体数的54.84%.北坡鱼鳞云杉径级呈现“L”形分布,DBH<14 cm的个体有123株,占总个体数的63.40%;臭冷杉径级呈现倒“J”形分布,主要为中、小径级个体,其中,DBH<8 cm和8 cm≤DBH<26 cm的个体数占总个体数的35.67%和57.89%;岳桦共计61株,各径级分布较为均匀.
3 讨 论
在海拔1 700 m左右,长白山西坡和北坡的植被类型都属于岳桦云冷杉林,为岳桦林与暗针叶林交错区.从物种组成看,相同海拔的长白山岳桦林与暗针叶林交错区北坡比西坡物种种类更加丰富.西坡乔木层以岳桦为主,零星伴生鱼鳞云杉,群落外貌与岳桦林带相似,接近岳桦林带分布的海拔高度下限;而北坡乔木层以鱼鳞云杉、臭冷杉等暗针叶树种为主,伴随少量岳桦均匀分布于内,群落外貌与暗针叶林带相似,接近暗针叶林带分布的海拔高度上限.长白山北坡陡峭、西坡平缓.有研究[9]表明,长白山西、北坡地形差异是导致该海拔段西、北坡群落结构异质性的重要原因.西坡草本层以小叶章为主,而小叶章为多年生、湿中生根茎禾草,本身常见于长白山西坡岳桦林下[10],尤其是在台风袭击西坡岳桦林后,形成大面积林窗,促进了小叶章等林下草本的大量繁殖,进一步巩固了岳桦林下小叶章种群的优势地位[10-11];北坡草本层则以暗针叶林中较为常见的耐阴性矮小植物二叶舞鹤草、杉蔓石松、广布鳞毛蕨等为主.
从径级结构看,西坡虽然有一部分岳桦更新苗(2 cm≤DBH<4 cm),但整体数量较少,仍然呈现出更新不良的状态,一方面可能与风干扰后小叶章大量繁殖,形成草根盘结,岳桦种子无法接触土壤,无法生根发芽有关[12];另一方面也可能与岳桦种子质量有关系,尽管岳桦为了适应环境恶劣的高海拔地带,其繁殖策略倾向于R策略,但是在种子的形成过程中,仍然容易受到诸如倒春寒等极端环境影响,导致出现种子瘪粒现象.西坡乔木个体总数相比北坡较少的原因可能与风干扰有关.大径级岳桦根系更加发达,深入土层,抓地力更强,比小径级岳桦更抗风干扰[13],当年风灾造成岳桦幼树死亡,保留了目前所见的大径级岳桦,造成小径级岳桦较少或缺失;而鱼鳞云杉因其个体较高且根系集中于地表,在严重的风干扰之下造成大部分大径级个体死亡,但是随着后期的不断更新,鱼鳞云杉小径级个体相对较多.此外,长白山北坡各层次群落的生物多样性指数均高于西坡,整体上北坡群落的生物多样性较高且群落稳定性高于西坡,预测西坡可能是北坡的演替前期,未来此处可能过渡为类似于北坡的以暗针叶林为主的群落交错区.
