红棕象甲触角感器的超微结构
2020-12-28段云博朱晓珍叶家桐
段云博 朱晓珍 叶家桐
摘要 为明确红棕象甲成虫触角感器的超微结构、种类、数量和分布情况,利用扫描电子显微镜观察雌雄成虫触角感器的外部形态特征、类型和分布。结果表明,红棕象甲触角共有6种感器,分别为刷形感器、簇状感器、毛形感器、刺形感器、芽孢形感器和叉形感器,其中刷形感器和簇状感器分别有2种亚型,毛形感器有3种亚型,刺形感器有4种亚型。雌雄之间部分感器的着生位置和数量存在明显的性二型现象,刺形感器Ⅲ型、刺形感器Ⅳ型以及叉形感器可能与雌虫寻找产卵场所时识别寄主挥发物有关。
关键词 红棕象甲; 触角感器; 扫描电镜; 超微结构
中图分类号: S 433.5
文献标识码: A
DOI: 10.16688/j.zwbh.2019516
Abstract In order to determine the ultrastructure, types, quantity and distribution of antenna sensilla in Rhynchophorus ferrugineus (Olivier) adults, the external morphological characteristics, types and distribution of the antenna sensilla of R.ferrugineus were observed by scanning electronic microscopy. The results showed that there were six types of sensilla on the antennae of R.ferrugineus, namely sensilla brush, sensilla cluster, sensilla trichodea, sensilla chaetica, sensilla gemmiformium and sensilla furcatea, including two kinds of sensilla brush, two kinds of sensilla cluster, three kinds of sensilla trichodea, and four kinds of sensilla chaetica. There was sexual dimorphism in the position and quantity of sensilla between male and female. Sensilla chaetica Ⅲ, sensilla chaetica Ⅳ and sensilla furcatea may be involved in the identification of host volatiles when the female looks for oviposition sites.
Key words Rhynchophorus ferrugineus; antennal sensilla; scanning electronic microscopy; ultrastructure
红棕象甲Rhynchophorus ferrugineus (Olivier)属鞘翅目Coleoptera象甲科Curculionidae,俗称锈色棕榈象、椰子甲虫和亚洲棕榈象甲等,是世界著名的毁灭性入侵害虫[1-2]。该虫原产于印度[3],现在除南极洲以外世界各地均有该虫为害[4-5],我国也早已将其列为林业检疫性害虫之一[6]。红棕象甲主要为害棕榈科植物,如加拿利海枣Phoenix canariensis、西谷椰子Metroxylon sagu、桄榔Arenga pinnata、肯氏鱼尾葵Caryota cumingii、槟榔Areca catechu、王棕Roystonea regia和大叶蒲葵Livistona saribus等[3],以幼虫蛀食树干或树冠心叶,食量巨大,一旦成功定殖,通常6~8个月即可令树木整株枯死,造成严重的经济损失[7]。近年来我国多地已陆续有该虫发生为害的报道[8-10]。
昆虫的感受器是帮助自身感知外界环境变化和刺激的重要结构,且大多数感受器集中分布于昆虫的触角上,它们在昆虫寻找寄主、配偶和产卵等方面发挥着重要作用[11]。因此,明确昆虫触角上感受器的种类、分布和功能是研究其两性交流机制的关键。同時,也是研究利用昆虫性信息素进行害虫防治工作的前提[12]。随着扫描电子显微镜技术在昆虫学领域内的广泛应用,国内外越来越多的学者利用该项技术对昆虫的触角感受器进行了研究[13]。其中,与红棕象甲同科的昆虫有枣食芽象甲Scythropus yasumatsui Kono et Morimoto[14]、褐纹甘蔗象Rhabdoscelus lineaticollis (Heller)[15]、米象Sitophilus oryzae (Linnaeus)[16-17]、香樟齿喙象Pagiophloeus tsushimanus Morimoto[18]、香蕉假茎象Odoiporus longicollis Olivier[19]、杨干象Cryptorrhynchus lapathi L.[20]、茶丽纹象甲Myllocerinus aurolineatus Voss[21]、樟子松球果象甲Pissodes validirostris Gyll.[22]、稻水象甲Lissorhoptrus oryzophilus Kuschel[23-24]和松梢象Pissodes nitidus Roel.[25]等象甲科昆虫的触角感器已被报道,但未见针对红棕象甲触角感器的研究报道。本文以红棕象甲雌雄成虫触角为研究对象,利用扫描电子显微镜研究该虫触角感受器的种类、形态和分布,以期为进一步探索红棕象甲的两性交流机制、寄主定位以及利用信息素开展该虫的防治工作奠定基础。
1 材料与方法
1.1 供试昆虫
红棕象甲成虫于2019年5月采自广西植物研究所(110.30°E, 25.08°N)内受害的董棕Caryota urens L.,在温度(25±1)℃、相对湿度(70±5)%和光周期L∥D=14 h∥10 h条件下用塑料盒(110 mm×55 mm×35 mm)单头饲养,每日用洁净、无病虫害的董棕茎干切块饲养待用。
1.2 试验方法
随机选取触角完整且健康的红棕象甲雌、雄成虫各10头,在体视显微镜下用镊子和解剖刀小心剪取整个触角。之后在2.5%戊二醛磷酸缓冲液(pH 7.4)中固定5 h,再将样品浸入70%乙醇溶液中,用超声波振荡仪(JP-020S,斯凯清洁设备有限公司,深圳,中国)振荡清洗10 min,除去表面的黏附物杂质,此步骤重复3次。经上述处理后,再将样品依次放入浓度梯度为70%、80%、90%和100%的乙醇溶液中逐级脱水,每个浓度梯度脱水30 min。将完成脱水后的样品自然晾干,最终将处理好的触角样品按照背面、腹面、内侧面和外侧面四个角度贴于粘有导电胶的样品台上;最后经108 auto离子溅射仪喷金后置于EV018型扫描电子显微镜(Carl Zeiss,Baden-Wurttemberg,Germany)下进行观察和拍照,扫描电子显微镜的工作电压为150 kV。利用Adobe Photoshop CS6(Adobe,San Jose,USA)软件处理图片以及测量成虫触角各节的长度和直径[26],重复测量5次后取平均值。所有毛形感器和芽孢形感器均在其着生部位(基部、中部和端部)各选取1 000 μm2面积内计数,其他感器均以其着生的整个小节计数,重复计数5次后取平均值。
1.3 特征描述
红棕象甲成虫触角感受器的分类以及命名参考Schneider[27]命名系统进行。
1.4 数据分析
文中所有数据均使用SPSS 21.0(IBM,Chicago,USA)软件进行数据计算与分析,采用独立样本t检验方法比较触角各节长度、直径和感受器数量在雌、雄虫间的差异,显著性水平P<0.05。
2 结果与分析
2.1 成虫触角的形态特征
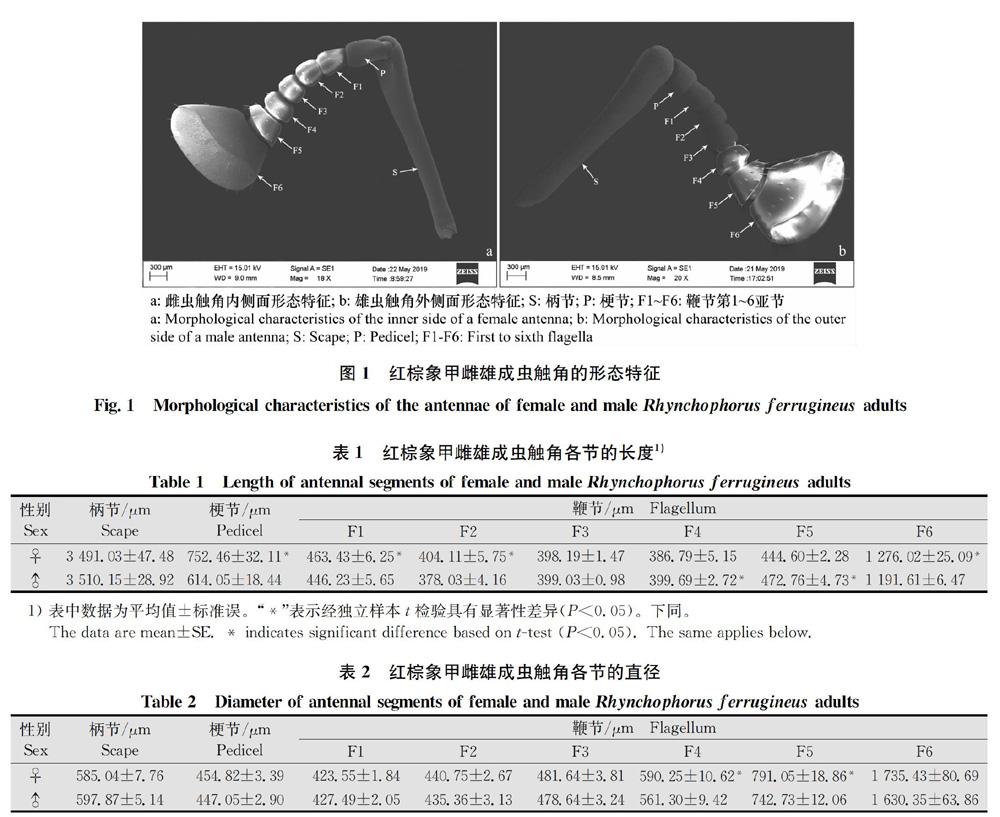
红棕象甲雌雄成虫触角着生于该虫喙的基部两侧,其外部形态特征一致,均为膝状,包含柄节(scape)、梗节(pedicel)和鞭节(flagellum)3个部分。其中柄节最长,约占触角总长的50%;梗节较短小;鞭节分为6个亚节,第1亚节(F1)基部较细长;第2~4亚节(F2-F4)形态基本一致,形似算盘珠;第5亚节(F5)马蹄形,第6亚节(F6)体积最大,末端有丰富的感器(图1)。
2.1.1 雌雄成虫触角各节的长度对比
由表1可知,红棕象甲雌雄成虫触角柄节以及鞭节第3亚节长度差异不显著(P>0.05),雌虫触角梗节、鞭节第1亚节、第2亚节以及第6亚节均显著(P<0.05)长于雄虫,而雄虫触角鞭节第4亚节和第5亚节均显著(P<0.05)长于雌虫。
2.1.2 雌雄成虫触角各节的直径对比
由表2可知,红棕象甲雌虫触角鞭节第4亚节和第5亚节直径均显著(P<0.05)大于雄虫,其余触角各节雌雄虫间差异不显著(P>0.05)。
2.2 成虫触角感器的种类、形态以及分布
通过扫描电子显微镜观察,发现红棕象甲成虫触角上共有6种类型感受器(图2和图3),分别为刷形感器、簇状感器、毛形感器、刺形感器、芽孢形感器和叉形感器,其中刷形感器和簇状感器分别有2种亚型,毛形感器有3种亚型,刺形感器有4种亚型。
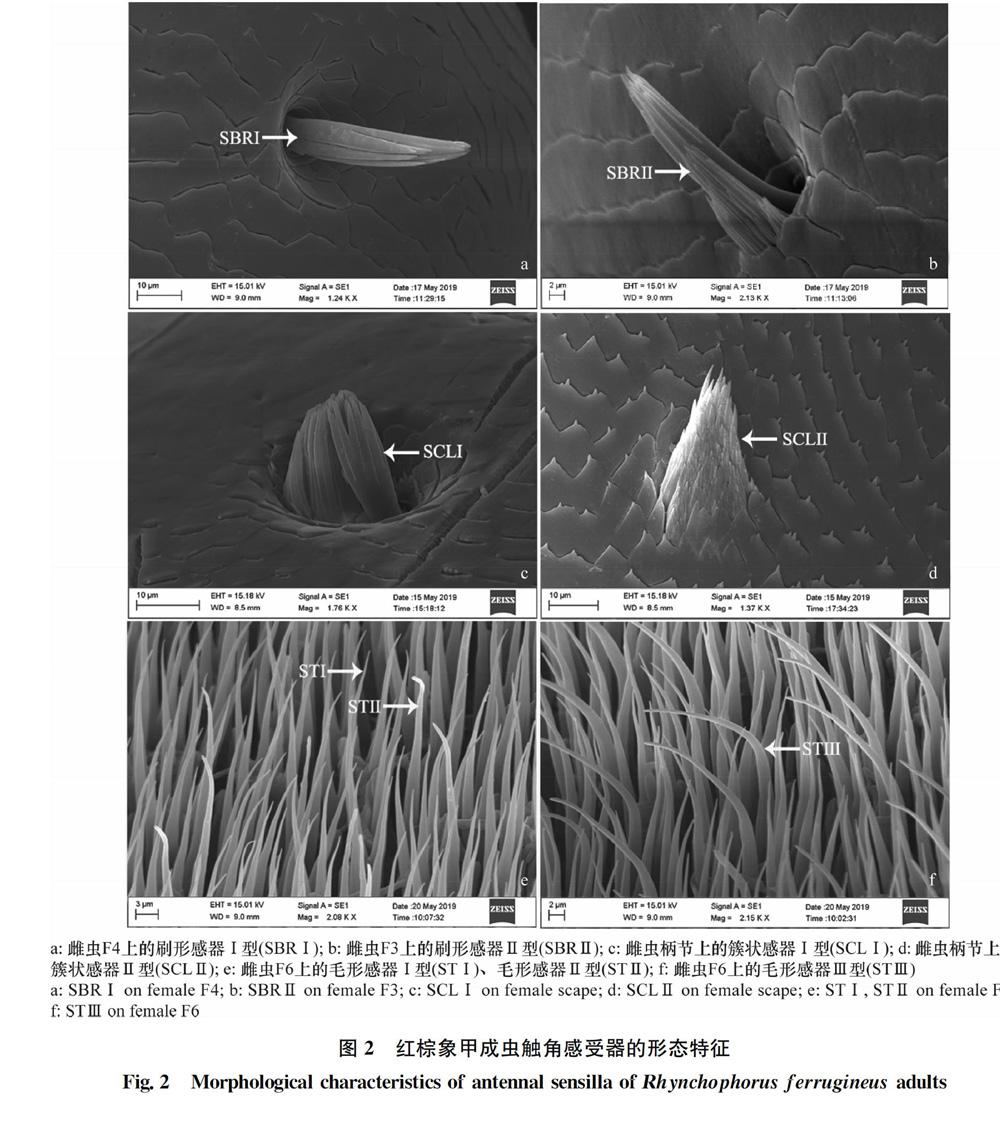
2.2.1 刷形感器(sensilla brush,SBR)
红棕象甲触角共有2种刷形感器(图2a, b),分别为刷形感器Ⅰ型(SBRⅠ)和刷形感器Ⅱ型(SBRⅡ)。刷形感器的两种亚型均只分布于雌雄虫触角的柄节、梗节和鞭节第1~4亚节。刷形感器Ⅰ型形似一把刷子,几乎与触角表皮呈90°,有基窝,表面光滑,从中下部开始有分叉,各节数量不均等;刷形感器Ⅱ型与刷形感器Ⅰ型较相似,但基部比刷形感器Ⅰ型多一簇感器,类似附属物,与表皮呈45°角,各节数量不均等。
2.2.2 簇状感器(sensilla cluster,SCL)
红棕象甲触角共有2种簇状感器(图2c, d),分别为簇状感器Ⅰ型(SCLⅠ)和簇状感器Ⅱ型(SCLⅡ)。簇状感器Ⅰ型见于触角柄节、梗节和鞭节第2~4亚节,但簇状感器Ⅱ型仅见于柄节和梗节。簇状感器Ⅰ型形似毛笔头,有基窝,较短小,几乎垂直于触角表皮,各节数量不均等;簇狀感器Ⅱ型形似火山爆发,无基窝,几乎紧贴触角表皮着生,各节数量不均等。
2.2.3 毛形感器(sensilla trichodea,ST)
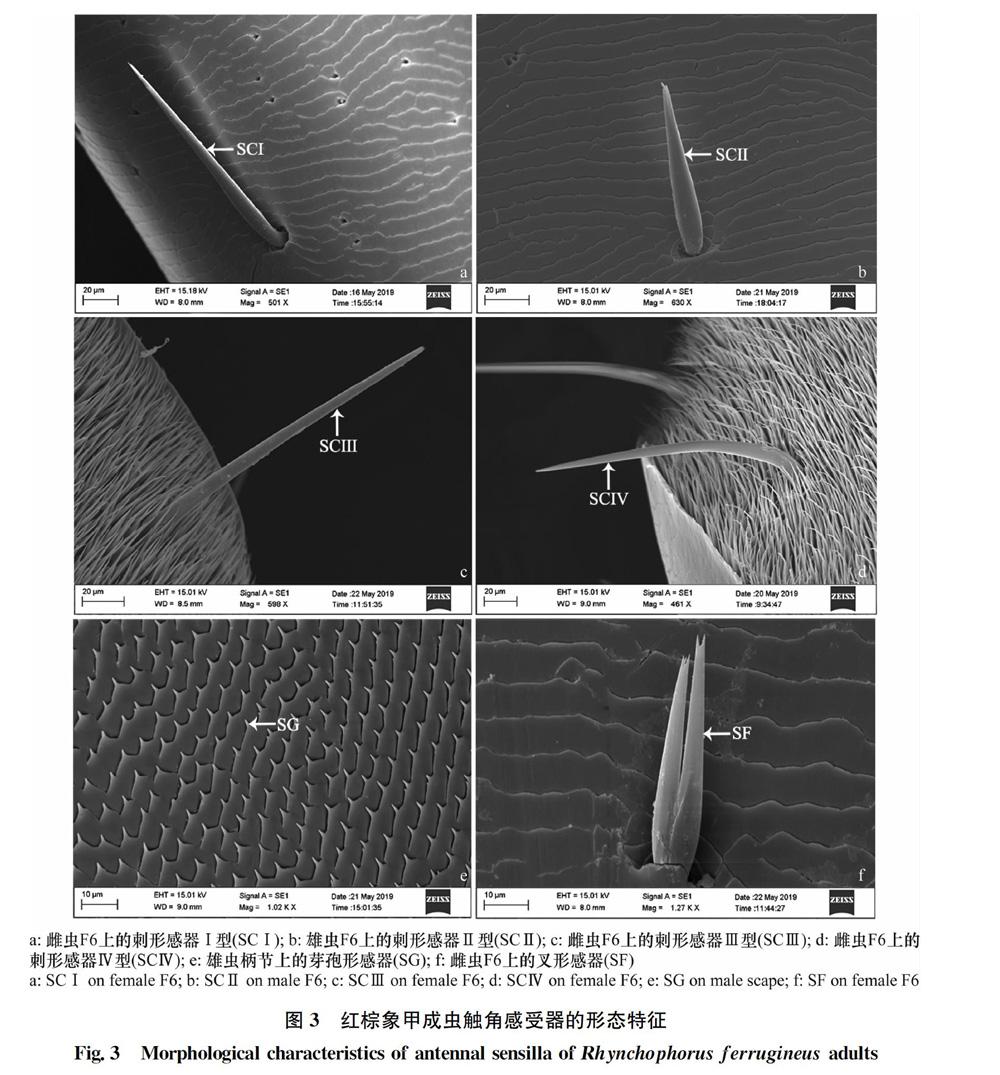
红棕象甲触角共有3种毛形感器(图2e, f),分别为毛形感器Ⅰ型(STⅠ)、毛形感器Ⅱ型(STⅡ)和毛形感器Ⅲ(STⅢ)型。这3种亚型均仅见于触角鞭节第6亚节。毛形感器Ⅰ型形似毛发,不弯曲,表面光滑,直立于触角表面,末端尖细,数量较多;毛形感器Ⅱ型端部弯曲,较钝,表面光滑,直立于触角表面,数量少于毛形感器Ⅰ型;毛形感器Ⅲ型从中部开始弯曲,指向触角基部,末端较钝,表面光滑,数量极少。
2.2.4 刺形感器(sensilla chaetica,SC)
红棕象甲触角共有4种刺形感器(图3a, b, c, d),分别为刺形感器Ⅰ型(SCⅠ)、刺形感器Ⅱ型(SCⅡ)、刺形感器Ⅲ型(SCⅢ)和刺形感器Ⅳ型(SCⅣ)。刺形感器Ⅰ型着生于触角鞭节第4~6亚节,刺形感器Ⅱ型着生于柄节以及触角鞭节第4~6亚节,刺形感器Ⅲ型以及刺形感器Ⅳ型仅见于触角鞭节第6亚节。刺形感器Ⅰ型针形,表面光滑,有基窝,末端尖,与表皮约呈30°角指向触角末端,各节数量不均等;刺形感器Ⅱ型与刺形感器Ⅰ型较相似,但长度短于Ⅰ型,表面光滑,有基窝,与表皮约呈30°角指向触角末端,各节数量不均等;刺形感器Ⅲ型刺状,几乎垂直于触角表皮,末端较钝,表面光滑;刺形感器Ⅳ型针形,从中下部开始向触角基部弯曲,与触角表皮大约呈80°指向触角基部,末端较尖,表面光滑。
2.2.5 芽孢形感器(sensilla gemmiformium,SG)
红棕象甲触角只有一种芽孢形感器(图3e),触角各节均有分布,各节数量不均等。该感器外形较短小,似刚长出的小芽尖,密布于触角各节,数量较多。
2.2.6 叉形感器(sensilla furcatea,SF)
红棕象甲触角只有一种叉形感器(图3f),仅见于触角鞭节第4~6亚节,从基部开始分叉,末端甚至继续分叉,各节数量不均等,较少。
2.3 雌雄成虫触角各节感受器的数量对比
由表3可知,雌虫触角梗節和鞭节第2亚节上刷形感器Ⅰ型数量显著多于雄虫(P<0.05),而雄虫触角柄节和鞭节第3亚节上刷形感器Ⅰ型数量显著多于雌虫(P<0.05);雄虫触角梗节、鞭节第1亚节和第3亚节上刷形感器Ⅱ型数量显著多于雌虫(P<0.05),而雌虫触角鞭节第4亚节上刷形感器Ⅱ型数量显著多于雄虫(P<0.05);雌虫触角柄节以及鞭节第4亚节上的簇状感器Ⅰ型数量显著多于雄虫(P<0.05),但雄虫触角梗节以及鞭节第3亚节上的簇状感器Ⅰ型数量显著多于雌虫(P<0.05);雌虫触角梗节上簇状感器Ⅱ型数量显著多于雄虫(P<0.05),但雄虫触角柄节上簇状感器Ⅱ型数量显著多于雌虫(P<0.05);雌虫触角鞭节第6亚节上刺形感器Ⅲ型和刺形感器Ⅳ型的数量均显著多于雄虫(P<0.05),但雄虫触角鞭节第4~5亚节上的刺形感器Ⅰ型和第4亚节、第6亚节上的刺形感器Ⅱ型数量均显著多于雌虫(P<005);雌雄虫触角间毛形感器Ⅰ型、毛形感器Ⅱ型和毛形感器Ⅲ型的数量均无显著性差异(P>005);雌虫触角柄节、梗节以及鞭节第1~5亚节上的芽孢形感器数量均显著多于雄虫(P<0.05),但雄虫触角鞭节第6亚节上的芽孢形感器数量显著多于雌虫(P<0.05);雌虫触角鞭节第6亚节上叉形感器的数量显著多于雄虫(P<0.05)。
3 讨论
本文利用扫描电子显微镜对红棕象甲触角超微结构进行观察,结果显示,该虫触角共有6种感器类型,其中毛形感器、刺形感器和芽孢形感器为象甲科昆虫触角常见感器类型,且已在多种象甲科昆虫中有报道[14-15,19];叉形感器在已报道的象甲科昆虫中也多次出现[17-18,21];此外,该虫触角上分布有2种刷形感器以及2种簇状感器,此类型感器系象甲科昆虫触角中首次报道,其功能尚不明确,故本文不做探讨。本研究表明,红棕象甲成虫触角感器的种类在雌雄之间没有显著性差异,但是部分感器的着生位置和数量却存在明显的性二型现象。
毛形感器是昆虫触角上一种常见的感受器,据现有资料显示,几乎所有象甲科昆虫都具有该类型感器。目前,该感器已被证实具有感受性信息素的功能[28-29],并且有研究表明其为感受性信息素的主要感器类型[30];也有学者认为毛形感器可以帮助昆虫感受寄主挥发物从而寻找合适的产卵场所[31]。本研究中于红棕象甲触角上共发现3种毛形感器,但其数量在雌雄间差异不显著,本文认为该虫触角的毛形感器可能兼具有感受性信息素和寄主挥发物的作用。
绝大多数昆虫触角都有刺形感器,其一般长于其他感器,凸出在外,通常认为该感器最先触碰到外界,属于感知机械刺激类的感器[32];但也有学者研究表明刺形感器也可以感受化学信息,属于化学感受器[33-34]。本研究中,雄虫触角鞭节第4~5亚节上刺形感器Ⅰ型的数量显著多于雌虫,可能与雄虫与雌虫交配时感知雌虫反抗活动的机械刺激有关,这一观点同褐纹甘蔗象触角感器的研究基本一致[15]。本研究发现雄虫触角柄节约有25根刺形感器Ⅱ型,但雌虫的相同部位却未见该类型感器,认为可能与雄虫在相互打斗竞争交配权时感知对方攻击的机械刺激有关;红棕象甲雌虫触角鞭节第6亚节上刺形感器Ⅲ型和刺形感器Ⅳ型的数量均显著多于雄虫,本文认同前人对同科昆虫松梢象Pissodes nitidus Roel.的观点,认为可能是执行嗅觉功能的化学感受器[25],与雌虫在寻找产卵场所时感受寄主植物挥发的化学信息素有关。
芽孢形感器是红棕象甲触角上分布最广的一种感器,前人研究表明,当触角受到外界机械刺激时,芽孢形感器可以起到缓冲重力的功能[14, 35]。本研究基本认同前人观点,该虫触角受到外界机械刺激时会主动收缩,该感器可能与触角折叠收缩时感知各节的机械刺激从而调整收缩程度有关,这一观点与同科昆虫褐纹甘蔗象触角感器的研究基本一致[15]。
叉形感器是红棕象甲触角上分布数量最少的一种感器,雌雄虫仅于触角鞭节第4~6亚节有发现。有研究表明该感器为一种机械感器,控制触角的运动[36];也有国外文献指出叉形感器可能是一种特殊的毛形感器,能够感知化学信息[37];有学者在对同科昆虫茶丽纹象甲的研究中指出,叉形感器在茶丽纹象甲与茶树之间的化学通讯过程中起到重要作用,可能用于感受寄主挥发物以及与种内信息素相关的化学物质[21];前人在针对同科昆虫米象Sitophilus oryzae的研究中发现雌虫叉形感器数量显著多于雄虫,认为可能与雌虫对粮食蛀孔形成卵窝以及产卵有关[17],本文中雌虫触角鞭节第6亚节上叉形感器的数量显著多于雄虫,基本认同该观点,认为该感器可能与红棕象甲雌虫识别寄主挥发物从而寻找合适的产卵场所有关。
本文结合前人的研究对红棕象甲触角部分感器的功能进行探讨,发现刺形感器Ⅲ型、刺形感器Ⅳ型以及叉形感器可能与雌虫寻找产卵场所时识别寄主挥发物有关。但具体各类型感器在该虫行为中所发挥的作用还有待进一步通过透射电镜、单细胞技术、电生理试验以及结合昆虫行为学测定来验证,以期从本质上研究各感器在红棕象甲行为反应中的作用。
参考文献
[1] 张润志, 任立, 孙江华, 等. 椰子大害虫—锈色棕榈象及其近缘种的鉴别(鞘翅目:象虫科)[J]. 中国森林病虫, 2003(2): 3-6.
[2] LI Lei, QIN Weiquan, MA Zilong, et al. Effect of temperature on the population growth of Rhynchophorus ferrugineus (Coleoptera: Curculionidae) on sugarcane [J]. Environmental Entomology, 2010, 39(3): 999-1003.
[3] 陈义群, 年晓丽, 陈庆. 棕榈科植物杀手—红棕象甲的研究进展[J]. 热带林业, 2011, 39(2): 24-28.
[4] FIABOE K K M, PETERSON A T, KAIRO M T K, et al. Predicting the potential world wide distribution of the red palm weevil Rhynchophorus ferrugineus (Olivier) [J]. Florida Entomologist, 2012, 95(3): 659-673.
[5] 李朝緒, 阎伟, 黄山春, 等. 10种杀虫剂对红棕象甲高龄幼虫的毒力测定[J]. 林业科技开发, 2013, 27(5): 72-74.
[6] 覃伟权, 李朝绪, 黄山春. 红棕象甲在中国的风险性分析[J]. 江西农业学报, 2009, 21(9): 79-82.
[7] 万婕, 阎伟, 刘丽, 等. 红棕象甲高龄幼虫冷驯化有效降温速率的确定[J]. 热带作物学报, 2014, 35(6): 1192-1197.
[8] 刘永耀. 成都市西南部林业有害昆虫调查及四种有害昆虫风险性评估[D]. 雅安:四川农业大学, 2017.
[9] 王谨, 王禹. 厦门园博园红棕象甲Rhynchophorus ferrugineus体表螨的种类和分布[J]. 福建农业学报, 2018, 33(12): 1307-1311.
[10]徐卫平. 赣州市红棕象甲发生情况初步调查及防治对策[J]. 现代园艺, 2019(11): 170-171.
[11]余海忠. 昆虫触角感受器研究进展[J]. 安徽农业科学, 2007, 35(14): 4238-4240.
[12]段云博, 吴海盼, 刘俊延, 等. 钩纹皮蠹触角感器的扫描电镜观察[J]. 植物保护, 2018, 44(1): 95-100.
[13]孙虹霞, 胡新军, 舒迎花, 等. 白背飞虱触角感器的扫描电镜观察[J]. 昆虫学报, 2006, 49(2): 349-354.
[14]张锋, 洪波, 王远征, 等. 枣食芽象甲触角感器的扫描电镜观察[J]. 西北农业学报, 2019, 28(8): 1373-1379.
[15]段云博, 董子舒, 张玉静, 等. 褐纹甘蔗象触角感器的扫描电镜观察[J]. 南方农业学报, 2017, 48(12): 2190-2196.
[16]姚富姣, 何磊, 李亚霄, 等. 米象触角及其感器的超微结构观察[J]. 陕西师范大学学报(自然科学版), 2016, 44(3): 79-84.
[17]李非凡, 段锦艳, 吴海晶, 等. 米象触角感器的扫描电镜观察[J]. 粮食储藏, 2017, 46(2): 1-8.
[18]顾天滋, 张丛丛, 苏鹏, 等. 香樟齿喙象成虫触角感器的微观特征分析[J]. 南京林业大学学报(自然科学版), 2017, 41(4): 89-94.
[19]赵海燕, 梁延坡, 陆永跃. 香蕉假茎象甲触角感受器的类型及分布[J]. 安徽农业科学, 2016, 44(8): 157-160.
[20]常立儒, 张璐, 王涛, 等. 杨干象触角感器的扫描电镜观察[J]. 植物检疫, 2014, 28(3): 23-29.
[21]高宇, 陈宗懋, 孙晓玲. 茶丽纹象甲触角感器的扫描电镜观察[J]. 植物保护, 2013, 39(3): 45-50.
[22]ZHAO Hongying, WANG Zhiying, GAO Yu, et al. Antennal morphology and sensillum distribution in Pissodes validirostris (Coleoptera: Curculionidae) [J]. Indian Journal of Agricultural Research, 2012, 46(1): 70-74.
[23]HIX R L, JOHNSON D T, BERNHARDT J L. Antennal sensory structures of Lissorhoptrus oryzophilus (Coleoptera: Curculionidae) with notes on aquatic adaptations [J]. Coleopterists Bulletin, 2003, 57(1): 85-94.
[24]KANG Guijuan, ZHU Zengrong, CHENG Jiaan, et al. Antennal sensilla of parthenogenetic and bisexual Lissorhoptrus oryzophilus (Coleoptera: Curculionidae) [J]. Florida Entomologist, 2012, 95(1): 8-15.
[25]YAN Shanchun, MENG Zhaojun, PENG Lu, et al. Antennal sensilla of the pine weevil Pissodes nitidus Roel. (Coleoptera: Curculionidae) [J]. Microscopy Research and Technique, 2011, 74(5): 389-396.
[26]廖任娅, 张金平, 唐睿, 等. 铃木氏果蝇触角和下颚须感器的超微形态特征及分布[J].植物保护, 2017, 43(1): 68-75.
[27]SCHNEIDER D. Insect antennae [J]. Annual Review of Entomology, 1964, 9(1):103-122.
[28]HILLIER N K, VICKERS N J. Physiology and antennal lobe projections of olfactory receptor neurons from sexually isomorphic sensilla on male Heliothis virescens [J]. Journal of Comparative Physiology A, 2007, 193(6): 649-663.
[29]BINYAMEEN M, ANDERSON P, IGNELL R, et al. Spatial organization of antennal olfactory sensory neurons in the female Spodoptera littoralis Moth: Differences in sensitivity and temporal characteristics [J]. Chemical Senses, 2012, 37(7): 613-629.
[30]ZHANG Lu, REN Lili, LUO Youqing, et al. Scanning electron microscopy analysis of the cephalic sensilla of Chrysolina aeruginosa Fald (Coleoptera: Chrysomelidae) [J]. Microscopy Research and Technique, 2013, 76(4): 423-431.
[31]徐偉, 毕嘉瑞, 刘梅, 等. 杨背麦蛾(鳞翅目:麦蛾科)触角感器的超微结构[J]. 林业科学, 2019, 55(5): 95-103.
[32]OCHIENG S A, PARK K C, ZHU J W, et al. Functional morphology of antennal chemoreceptors of the parasitoid Microplitis croceipes (Hymenoptera: Braconidae) [J]. Arthropod Structure and Development, 2000, 29(3): 231-240.
[33]SEADA M A. Antennal morphology and sensillum distribution of female cotton leaf worm Spodoptera littoralis (Lepidoptera: Noctuidae) [J]. The Journal of Basic & Applied Zoology, 2015, 68: 10-18.
[34]YANG S, LIU H, ZHANG J T, et al. Scanning electron microscopy study of the antennal sensilla of Monema flavescens Walker (Lepidoptera: Limacodidae) [J]. Neotropical Entomology, 2017, 46(2): 175-181.
[35]BARTLET E, ROMANI R, WILLIAMS I H, et al. Functional anatomy of sensory structures on the antennae of Psylliodes chrysocephala L. (Coleoptera: Chrysomelidae) [J]. International Journal of Insect Morphology and Embryology, 1999, 28(4): 291-300.
[36]王全坡. 秦岭华山松两种主要小蠹虫触角感器的超微结构[D]. 杨凌:西北农林科技大学, 2013.
[37]WANG Pingyan, ZHANG Zhen, KONG Xiangbo, et al. Antennal morphology and sensilla ultrastructure of three Tomicus species (Coleoptera: Curculionidae, Scolytinae) [J]. Microscopy Research and Technique, 2012, 75(12): 1672-1681.
(责任编辑:田 喆)
