橡胶籽油对脂多糖刺激蛋鸡产蛋性能、肠道形态和免疫功能的影响
2020-12-21闻治国吴学壮齐志国李复煌杨培龙
闻治国,吴学壮,齐志国,李复煌*,杨培龙*
(1.中国农业科学院饲料研究所,农业部饲料生物技术重点实验室,北京 100081;2.安徽科技学院动物科学学院,安徽蚌埠 233100;3.北京市畜牧总站,北京 100107)
橡胶籽是天然橡胶种植业的副产物,种仁含油量为40%~50%,资源丰富且廉价,但长期以来,橡胶籽并没有得到有效开发利用[1]。橡胶籽经脱壳压榨后产生橡胶籽粕和橡胶籽油(Rubber seed oil,RSO)。RSO 中脂肪酸主要含有亚油酸(C18:2)、亚麻酸(C18:3)、油酸(C18:1)、硬脂酸(C18:0)和棕榈酸(C16:0),其中多不饱和脂肪酸占65.77%,且亚麻酸的含量高达28.72%,是豆油3~4 倍[2]。RSO 没有毒性[3],具有抗氧化功能[4],对小鼠生长发育没有负作用[5]。在畜禽上,RSO 能够提高牛奶ω-3 多不饱和脂肪酸含量[6]、增强奶牛免疫功能[7]以及提高罗非鱼肠道消化酶活性[8]。前期研究发现,日粮中添加4% RSO 能够促进蛋鸡产蛋性能,显著提高蛋黄α-亚麻酸(ALA)、二十二碳六烯酸(DHA)和二十碳五烯酸(EPA)等ω-3 多不饱和脂肪酸含量,饲喂8 周,整枚鸡蛋DHA 含量较对照组提高了45.6%[9]。动物体内蓄积的ω-3 多不饱和脂肪酸在机体抗炎上发挥重要作用。DHA 和EPA 能置换细胞膜磷脂中的花生四烯酸(AA,n-6)、竞争环氧化酶(COX)和脂氧合酶从而减少来源于AA 的炎性介质产生,减轻炎症反应[10],同时能抑制TLR4/NF-κB炎症通路中TLR4和NF-κB等相关基因的表达[11-12],降低TNF-α、IL-1β和IL-6 等炎症因子的生成[13-14]。RSO 是一种典型的富含ω-3 多不饱和脂肪酸的木本植物油,并能促进鸡蛋ω-3 多不饱和脂肪酸蓄积,但RSO 能否缓减蛋鸡炎症反应迄今未见报道。因此,本研究通过脂多糖(LPS)刺激蛋鸡产生炎症反应,并进一步研究RSO对LPS 刺激蛋鸡产蛋性能、肠道形态结构和免疫功能的影响,为RSO 在蛋鸡日粮中应用以及利用RSO 缓减蛋鸡炎症反应提供理论和方法依据。
1 材料与方法
1.1 试验材料 RSO 由西双版纳华坤生物科技有限责任公司提供;LPS 购自国药集团。罗曼青年蛋鸡(15 周龄)购自北京大发正大有限公司,饲喂至产蛋高峰期(25周龄)。
1.2 试验设计及动物 本试验采用2×2 因子完全随机试验设计,将192 只25 周龄健康、产蛋率和体形相近的罗曼蛋鸡随机分为4 组,每组4 个重复,每重复12 只鸡,每个重复2 个鸡笼,每个鸡笼6 只鸡,试验期4 周。对照组饲喂常规基础日粮,LPS 组(模型组)饲喂基础日粮并攻毒LPS,RSO 组饲喂RSO 日粮,RSO+LPS 组饲喂橡胶籽油日粮并攻毒LPS。LPS 攻毒分别在试验第19、21、23、25、27 天09:00 腹腔注射LPS(1 mg/kg体重,用生理盐水溶解),对照组和RSO 组注射等量的生理盐水。本试验经中国农业科学院饲料研究所实验动物伦理委员会批准。
1.3 试验日粮 试验日粮采用玉米-豆粕型基础日粮,参照我国《鸡饲养标准》(NY/T 33-2004)和美国NRC(1994),在试验日粮中添加4%橡胶籽油,并通过调节饲料原料的比例使试验日粮营养水平完全满足产蛋高峰期蛋鸡营养需要,试验日粮组成及营养水平见表1。
1.4 饲养管理 采用封闭式鸡舍,2 层阶梯式笼养,同一处理各重复均匀分布于鸡舍不同位置的上下层,以消除环境、位置对测定指标的影响;自然光照与人工光照相结合,保证每日光照16 h,光照强度10~20 lx;正压横向通风,屋顶进风,温度25~30℃,相对湿度50%~75%;饲料为干粉料,每日布料2 次,匀料4 次,自由采食,乳头式饮水器自由饮水;每日捡蛋1 次;每5 d 清粪、鸡舍消毒1 次,按照常规防疫程序进行防疫。
1.5 测定指标与方法
1.5.1 生产性能 试验期间,每天以重复为单位,记录每个重复的总蛋重、鸡蛋数,每周清料1 次,最后计算整个试验期的产蛋率、总产蛋量、平均蛋重、只日产蛋量、平均日采食量及料蛋比。
1.5.2 血液免疫生理生化指标 试验结束时,每个重复挑选接近平均体重的蛋鸡2 只,采集少许全血于5 mL EDTA 抗凝管,采用德国ABX 血细胞分析仪(Pentra120)测定白细胞计数(WBC)、红细胞计数(RBC)、血红蛋白浓度(HGB)、单核细胞绝对值(MON)、淋巴细胞绝对值(LYM)、中性粒细胞绝对值(MEUT)和嗜碱性粒细胞绝对值(BAS)。同时,采集血液置于10 mL 真空采血管(内含肝素钠)内,室温静置4 h 后,3 500 r/min 离心10 min 制备血浆,分装于1.5 mL EP管中,-20℃下保存备测。然后使用试剂盒测定谷草转氨酶(AST)、谷丙转氨酶(ALT)、总蛋白(TP)、白蛋白(ALB)、球蛋白(GLB)、免疫球蛋白M(IgM)、免疫球蛋白G(IgG)、免疫球蛋白A(IgA)、白细胞介素6(IL-6)、肿瘤坏死因子α(TNF-α),试剂盒购自南京建成生物工程研究所。
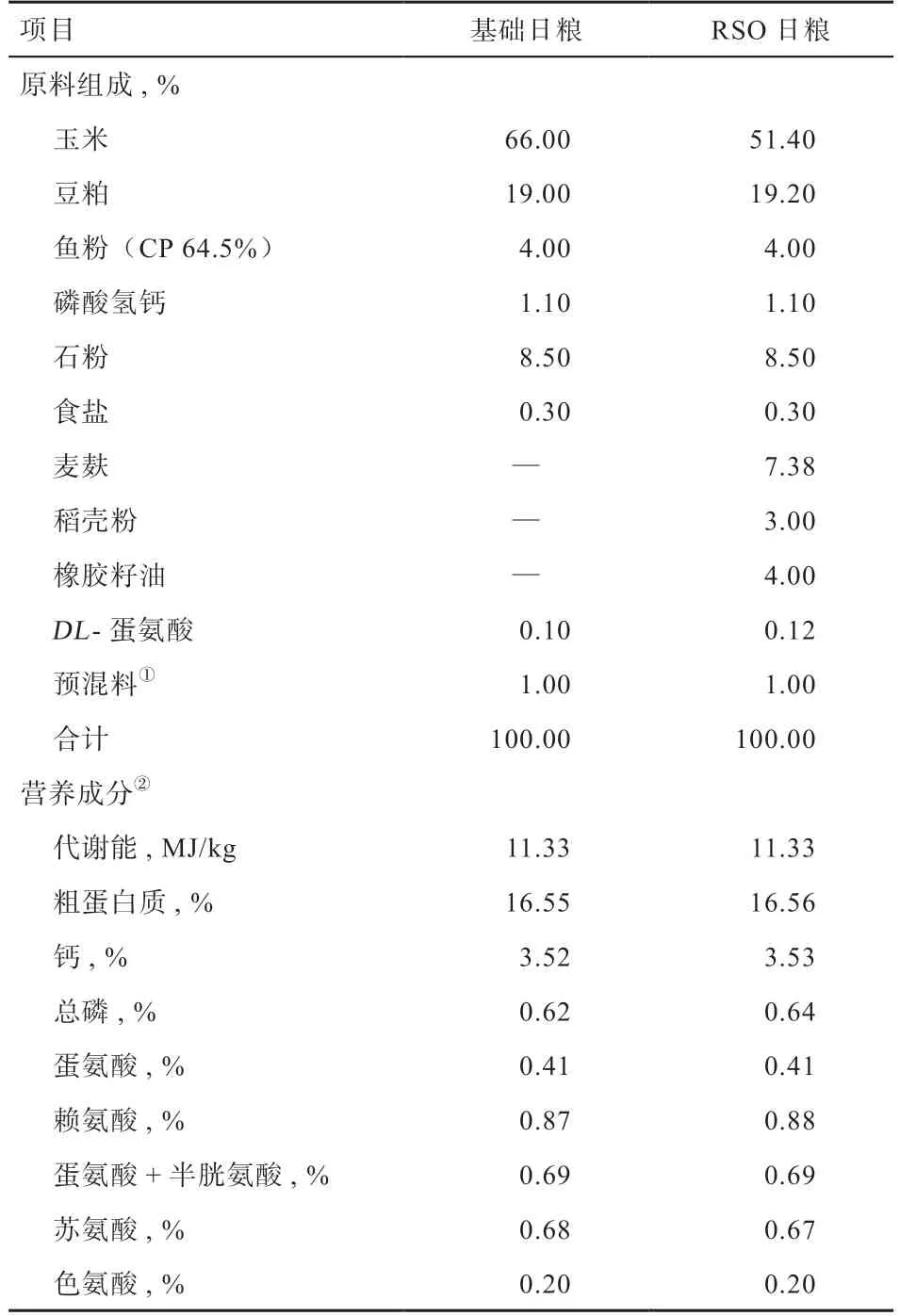
表1 日粮组成及营养水平(风干基础)
1.5.3 炎症基因mRNA 表达分析 采集血液后的蛋鸡,用手术剪刀打开腹腔,无菌操作取出少许肝脏、脾脏和回肠中段组织,分别置于灭菌的1.5 mL 冻存管中,立即保存于-196℃液氮速冻后转移到-80℃冰箱中,采用SYBR Green 实时荧光定量RT-PCR 法测定IL-6、IL-1β、TLR4、NF-κB基因mRNA 表达量,并用核酸测定仪(Nanodrop®ND-2000)测定总RNA 纯度和浓度,然后应用反转录试剂盒(TaKaRa 公司,日本)获取cDNA,-80 ℃保存待用。按照SYBR Premix Ex TaqTM(TaKaRa 公司,日本)试剂盒说明书进行荧光定量PCR,反应仪器为7500 荧光检测系统(Applied Biosystems)。PCR 扩增完成后用熔解曲线和琼脂糖凝胶电泳法分析验证产物的特异性。炎症基因IL-6、IL-1β、TLR4、NF-κB和内参基因GAPDH的引物序列见表2,基因表达的结果采用2-△△CT法进行分析。

表2 引物序列及序列号
1.5.6 肠道形态结构 用手术刀取回肠中段约2 cm 置于10% 福尔马林固定24 h 后,经浸洗、脱水、透明、浸蜡、包埋、切片、染色等步骤制作组织切片,切片厚度为6 μm,经苏木精-伊红(HE)染色后,在显微镜下观察组织病理学形态变化,并随机选取完整、走向平直的10 根绒毛,应用图像分析软件(Image pro-plus 6.0)分析绒毛顶端到隐窝入口处的长度(绒毛高度)和绒毛基部到黏膜下层长度(隐窝深度),然后计算绒毛高度和隐窝深度比值。
1.6 统计分析 试验数据采用SAS 9.0 中Two-Way ANOVA进行统计分析,当RSO 和LPS 交互作用显著时,采用LSD 法进行多重比较,结果以平均值和标准误(SEM)表示,以P<0.05 作为差异显著性判断标准。
2 结果
2.1 RSO 对LPS 刺激蛋鸡产蛋性能的影响 由表3 可知,与对照组相比,日粮中添加RSO 能够提高蛋鸡产蛋率、总产蛋量、只日产蛋量和平均日采食量(P<0.05),LPS 攻毒后蛋鸡产蛋率、总产蛋量、只日产蛋量和平均日采食量降低(P<0.05);与LPS 组相比,RSO+LPS组蛋鸡产蛋率、总产蛋量、只日产蛋量、平均日采食量升高(P<0.05);添加RSO 和LPS 攻毒均对平均蛋重和料蛋比无显著作用;RSO 和LPS 在蛋鸡产蛋性能上不存在显著交互作用。
2.2 RSO 对LPS 刺激蛋鸡血液免疫生理生化指标的影响由表4 可知,与对照组相比,LPS 攻毒能够提高蛋鸡血液WBC、LYM、AST、IgA、IL-6 和TNF-α水平(P<0.05),日粮中添加RSO 能够降低蛋鸡血液IgA、AST、IL-6 和TNF-α水平(P<0.05);与LPS 组相比,RSO+LPS 组蛋鸡血液IgA、AST、IL-6 和TNF-α水平降低(P<0.05);日粮RSO 和LPS 攻毒在血液IgA 和TNF-α上存在交互作用(P<0.05),添加RSO 能够缓减蛋鸡LPS 攻毒诱导的IgA 和TNF-α升高情况;蛋鸡血液其他免疫生理生化指标变化不大(P>0.05)。
2.3 RSO 对LPS 刺激蛋鸡免疫器官基因mRNA 表达的影响 由表5 可知,与对照组相比,LPS 攻毒提高蛋鸡回肠、肝脏和脾脏TLR4、NF-κB、IL-1β和IL-6基因mRNA 表达量(P<0.05),日粮中添加RSO 能够降低肝脏和脾脏TLR4、NF-κB、IL-1β和IL-6基因以及回肠TLR4、IL-1β和IL-6基因mRNA 表达量(P<0.05);与LPS 组相比,RSO+LPS 组蛋鸡回肠、肝脏和脾脏IL-6、IL-1β和TLR4基因mRNA 表达量降低(P<0.05);RSO 和LPS 在回肠TLR4和脾脏IL-1β基因上存在交互作用(P<0.05),同时在肝脏IL-6、NF-κB和TLR4基因上有互作效应(P<0.05)。表明日粮中添加RSO能够降低LPS 刺激蛋鸡组织TLR4/NF-κB信号通路中关键基因表达。
2.4 RSO 对LPS 刺激蛋鸡回肠形态结构的影响 由图1可知,与对照组相比,LPS 组蛋鸡回肠组织绒毛单层柱状上皮细胞脱落、断裂,杯状细胞结构消失,损伤累积隐窝浅层,黏膜层及黏膜下层结构均正常;RSO 添加组蛋鸡回肠组织绒毛上皮细胞的单层柱状上皮细胞排列规则有序,杯状细胞数量丰富,隐窝结构清晰,黏膜层及黏膜下层结构均正常,小肠结构整体较正常。由表6可知,用图像软件分析后发现,与对照相比,LPS 组蛋鸡回肠隐窝深度提高(P<0.05),绒毛高度和绒毛高度/隐窝深度降低(P<0.05),RSO 组蛋鸡回肠绒毛高度提高,绒毛高度/隐窝深度提高(P<0.05)。与LPS 组相比,RSO+LPS 组蛋鸡回肠绒毛高度升高,隐窝深度降低(P<0.05)。结果表明日粮中添加RSO 能有效维持LPS 刺激下蛋鸡回肠组织形态结构。
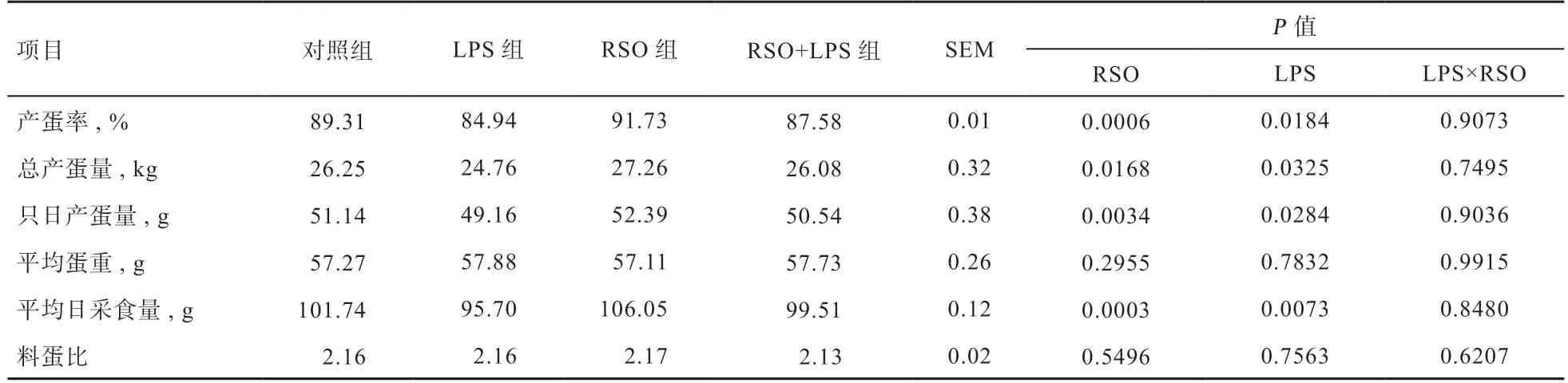
表3 RSO 对LPS 刺激蛋鸡产蛋性能的影响
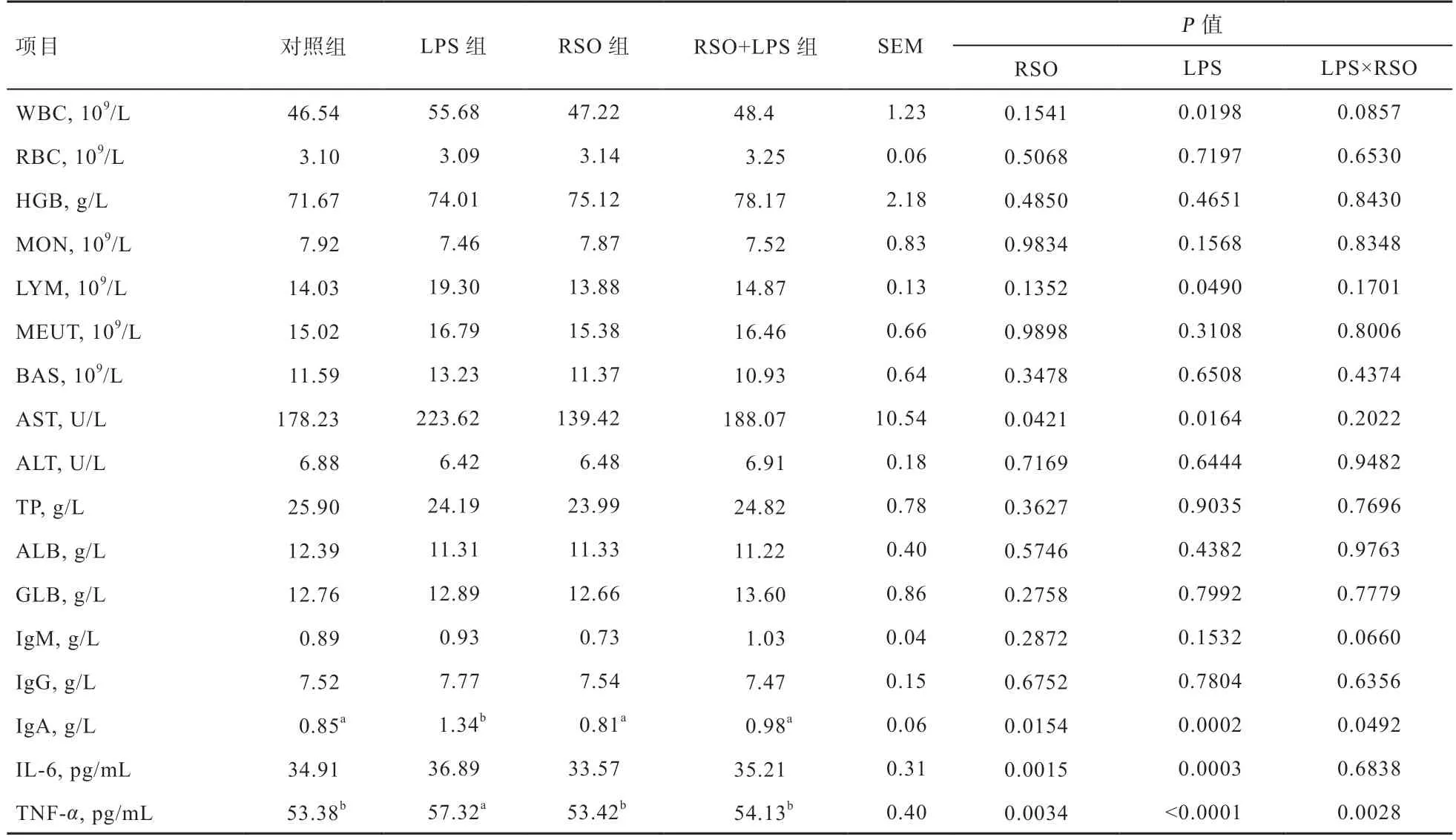
表4 RSO 对LPS 刺激蛋鸡血液免疫生理生化指标的影响
3 讨 论
3.1 RSO 对LPS 刺激蛋鸡产蛋性能的影响 前期研究发现,蛋鸡日粮中添加4% RSO 蛋鸡产蛋率、日产蛋量和采食量较对照组分别提高了6.52%、5.12%和9.03%[9]。本试验中,日粮中添加4% RSO 能显著提高蛋鸡采食量,同时产蛋量和日产蛋量有升高的趋势,与前期研究结果一致。关于ω-3 不饱和脂肪酸植物油对蛋鸡产蛋性能影响的研究一直是争论的话题。Petrović 等[15]研究发现1%亚麻籽油能够提高鸡蛋重量,进一步添加会降低蛋重。而有的研究发现蛋鸡饲料中添加ω-3 不饱和脂肪酸植物油(如橄榄油[16]和榛子油[17])对蛋鸡产蛋性能无显著影响,甚至一些研究结果显示蛋鸡产蛋性能与日粮中ω-3 不饱和脂肪酸植物油含量负相关[18-19]。造成这些差异的原因可能与日粮成分、蛋鸡品种、营养水平等不同相关。LPS 又称内毒素,是革兰氏阴性菌细胞壁的成分,能够成功诱导家禽产生炎症反应,并降低家禽生产性能[20]。本试验结果发现,与对照组相比,LPS 组蛋鸡产蛋率、产蛋量和料蛋比均显著降低;与LPS 组相比,RSO+LPS 组蛋鸡产蛋率和采食量显著提高,这说明RSO 能够缓减LPS 刺激导致的蛋鸡生产性能降低的现象,主要原因可能是RSO 能够抑制蛋鸡肝脏和脾脏等免疫器官炎症基因mRNA 表达,进而缓减蛋鸡机体炎症反应,血液免疫因子分泌量降低,同时维持了蛋鸡肠道形态结构,以此来保持蛋鸡正常生长发育。
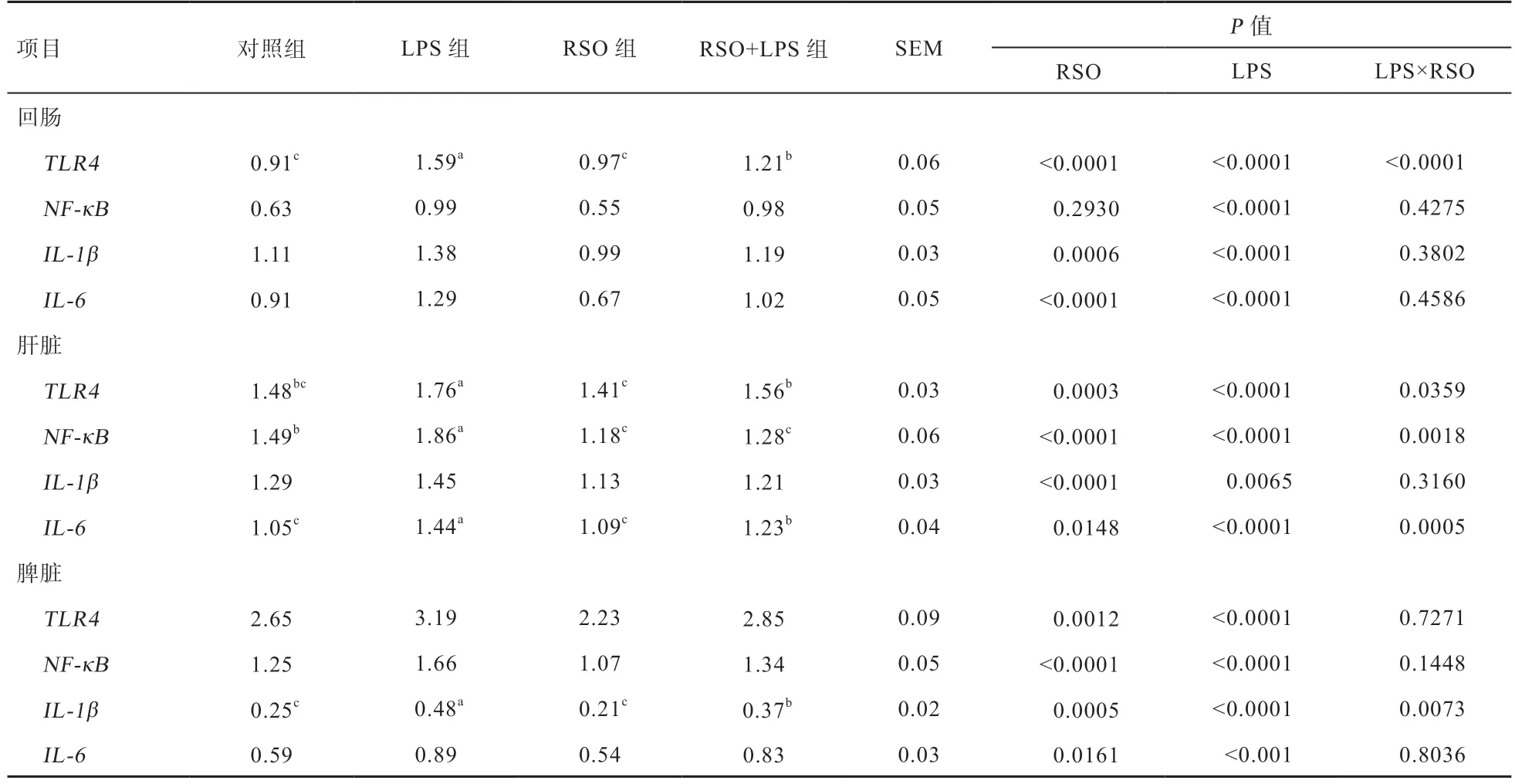
表5 RSO 对LPS 刺激蛋鸡组织基因mRNA 表达量的影响
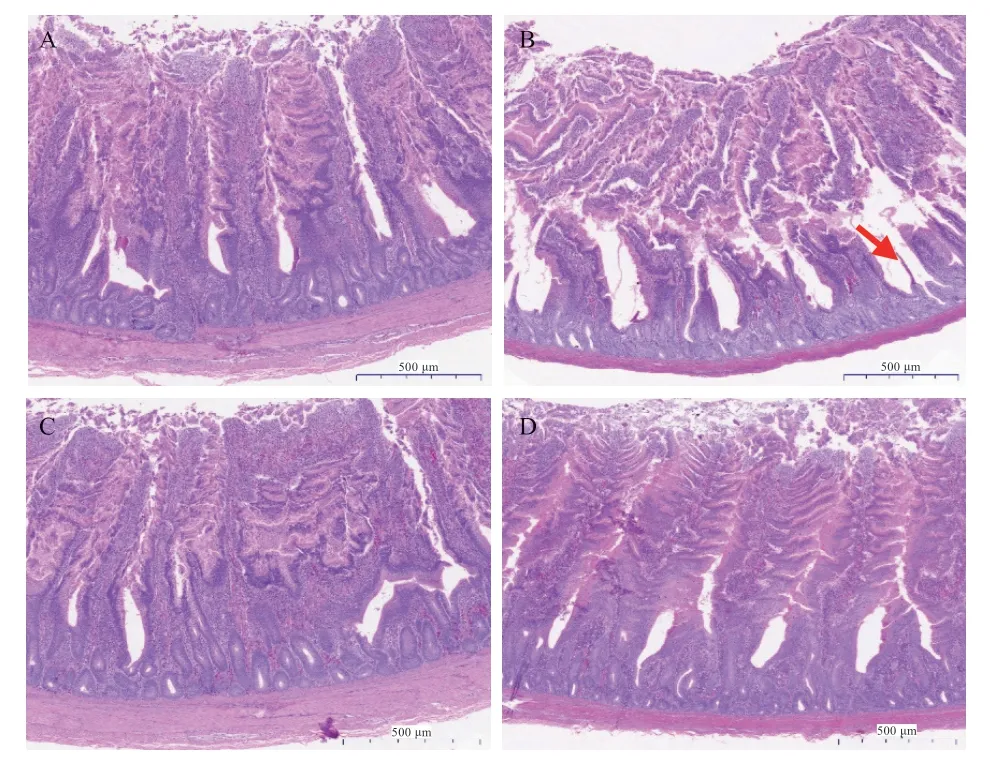
图1 RSO 对LPS 刺激蛋鸡回肠组织形态结构的影响
3.2 RSO 对LPS 刺激蛋鸡血液免疫生理生化指标的影响血液生理生化指标的含量和变化规律是反映动物机体新陈代谢的重要指标。机体免疫应答过程中,白细胞是发挥作用的最主要免疫细胞,是衡量机体免疫反应的重要指标,能反映机体免疫功能水平的高低。本试验中,与对照组相比,LPS 组蛋鸡血液WBC 和LYM 显著升高;与LPS 组相比,RSO+LPS 组血液WBC 和LYM 显著降低,表明RSO 能够提高LPS 刺激蛋鸡的免疫能力。Gandhi 等[21]研究发现,与花生油组相比,橡胶籽油组小鼠血液WBC 含量显著降低。血浆转氨酶检查是评估肝细胞损伤的重要途径,而血浆免疫球蛋白含量是体现动物机体免疫功能的重要标志。LPS 诱导蛋鸡产生炎症反应,肝脏细胞通透性增加,血浆AST 含量显著升高,与对照组相比,RSO 组蛋鸡血液AST 含量降低,与张新党等[8]在罗非鱼上的研究结果相反,该研究显示添加橡胶籽油后罗非鱼血浆和肝脏ALT 和AST 活性无显著差异。IgA 是机体黏膜局部抗感染免疫的主要抗体,LPS 诱导蛋鸡血浆IgA 含量升高,添加RSO 后蛋鸡血浆IgA 含量显著下降,血浆IL-6与IgA 趋势相同,表明日粮RSO 能够缓解蛋鸡LPS 诱导的炎症反应。Pi 等[7]研究发现,奶牛基础日粮中添加RSO,血浆IgA 含量无明显变化,与本试验结果相反,原因可能是家禽与反刍动物生理代谢上存在差异。
TNF-α是一种单核因子,主要由活化的巨噬细胞产生,主要作用为抗肿瘤和抗感染,LPS 是TNF-α的激活剂。研究发现,ω-3 多不饱和脂肪酸能够抑制T 淋巴细胞增殖分化,并且在基因水平上降低炎症因子表达,降低炎症因子分泌,起到抗炎和促进炎症消退的作用[22]。本试验中,RSO 能够发挥ω-3 多不饱和脂肪酸抗炎的作用,日粮添加RSO 能降低LPS 刺激蛋鸡血浆TNF-α水平。在LPS 诱导的小鼠急性肺损伤模型中,DHA 代谢产物能够抑制中性粒细胞的浸润和黏附,降低炎症因子TNF-α和IL-6的生成,与本试验中RSO 缓解蛋鸡LPS 诱导的炎症反应结果一致。同时,本试验对照组和RSO 组之间血浆TNF-α水平无显著差异,与Pi 等[7]在奶牛上的研究结果相反,该结果显示,基础日粮中添加RSO 后,奶牛血浆TNF-α水平显著下降。在其他ω-3 多不饱和脂肪酸研究中,不同剂量鱼油补充能够降低中老年人血液C-反应蛋白水平,但对TNF-α和IL-6 含量无显著影响[23]。RSO 作为一种ω-3 多不饱和脂肪酸植物油,可能通过影响LPS 刺激蛋鸡肝脏和脾脏中NF-κB和TLR4基因表达,进而调控TLR4/NF-κB信号通路中下游炎症因子表达,从而调节炎症因子表达量。
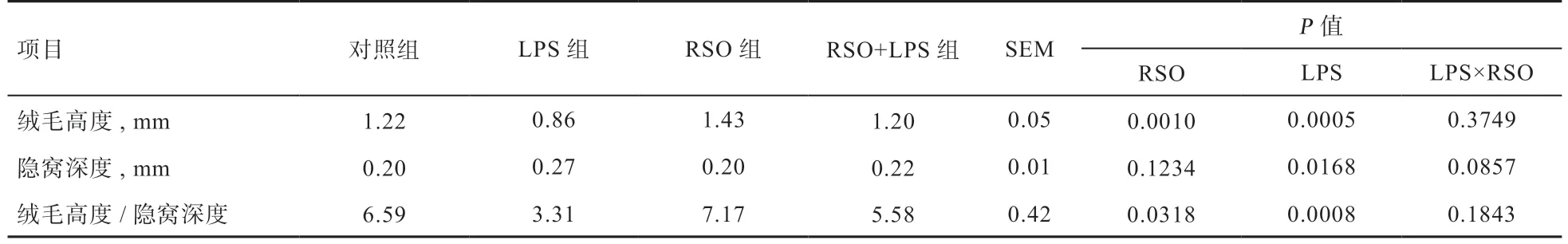
表6 RSO 对LPS 刺激蛋鸡回肠绒毛高度和隐窝深度的影响
3.3 RSO 对LPS 刺激蛋鸡免疫器官基因mRNA 表达的影响 TLR 是天然免疫受体家族,其中TLR4是自然免疫系统识别病原微生物的最主要受体,主要表达于树突状细胞、巨噬细胞、淋巴细胞等表面[24]。肠道上皮细胞TLR4 蛋白是LPS 的结合受体,LPS 是TLR4 蛋白的激活剂,细胞是否能对LPS 的刺激发生反应与该细胞膜上TLR4基因表达水平有密切的关系[20,25]。本试验中,LPS 刺激蛋鸡后,蛋鸡肝脏、回肠和脾脏TLR4基因mRNA 表达量显著上升。Avlas 等[25]研究发现,LPS 诱导了小鼠肠道炎症因子释放和肠道黏膜结构损伤,而TLR4敲除(TLR4-/-)小鼠受到LPS 诱导后,肠道炎症因子表达量显著降低。LPS 诱导动物产生炎症反应通过TLR4/NF-κB信号通路发挥作用[26-27]。LPS 进入机体后,首先与LPS 结合蛋白(LBP)结合成LPSLBP 复合物,使LPS 的生物活性显著增强[28]。LPSLBP 复合物能够被细胞表面的CD14 和髓样分化蛋白2(MD2)受体识别并结合,接着CD14 将LPS-LBP 复合物递呈给TLR4,TLR4 通过髓样分化因子88(myeloid differentiation factor 88,MyD88)的介导,激活IL-1受体相关激酶(IL-1 receptor-associated kinase,IRAK),激活的IRAK 进一步激活NF-κB,其与相关基因的启动子作用,从而促进细胞因子和炎性介质的合成和释放,如TNF-α、INF-γ、IL-1β、IL-6 和氧自由基等,引起炎症反应[24,27]。因此,本试验中LPS 刺激蛋鸡后,蛋鸡回肠、肝脏和脾脏NF-κB、IL-6、IL-1β基因mRNA 表达量显著上升。前人研究发现,ω-3 多不饱和脂肪酸能够介导TLR4/NF-κB信号通路调控动物肠道炎症反应[11],降低TNF-α、IL-1β和IL-6 等炎症因子的生成[13-14]。RSO 能够促进蛋鸡ω-3 多不饱和脂肪酸蓄积[9],降低LPS 刺激蛋鸡组织IL-6、NF-κB和TLR4基因mRNA表达量,与前人研究结果一致。综上所述,日粮RSO能够通过TLR4/NF-κB信号通路缓解蛋鸡LPS 诱导产生的炎症反应。
3.4 RSO 对LPS 刺激蛋鸡回肠形态结构的影响 家禽肠道是营养物质消化和吸收的主要场所,也是体内最大的免疫器官,是机体防御体系的第一道屏障,在维持机体正常营养代谢和免疫防御等方面发挥着重要作用。肠道黏膜作为家禽机体最重要的黏膜系统,其形态结构和功能的完整性,是维护肠道健康的有效屏障[29]。小肠绒毛高度和隐窝深度是评价肠道黏膜功能的敏感指标。本试验中,LPS 诱导蛋鸡回肠组织绒毛单层柱状上皮细胞脱落、断裂,杯状细胞结构消失,损伤累积隐窝浅层,LPS 组蛋鸡回肠绒毛高度降低,隐窝深度加深,绒毛高度和隐窝深度比值降低,表明LPS 刺激能够引起蛋鸡肠道炎症反应,炎症因子大量释放损坏肠道黏膜结构,肠道组织炎症基因表达量升高,这与在肉鸡[20]和猪[26]上研究结果一致。同时,RSO 能够缓解蛋鸡LPS 诱导的肠道损伤状况,RSO+LPS 组蛋鸡回肠组织绒毛上皮细胞的单层柱状上皮细胞排列规则有序,杯状细胞数量丰富,隐窝结构清晰,黏膜层及黏膜下层结构均正常,小肠结构整体较正常。Chwen 等[30]研究发现,日粮添加中链脂肪酸能够缓解LPS 诱导的猪肠道结构损伤,可增加猪十二指肠、空肠和回肠绒毛高度。RSO 能够缓减LPS 刺激蛋鸡肠道黏膜损伤,原因可能是ω-3 多不饱和脂肪酸除了能够抑制炎症缓减肠道损伤外,也是细胞膜的结构成分,能通过改变膜磷脂的脂肪酸结构进而改变膜流动性,达到调节膜相关信号分子受体功能[22]。
4 结 论
本试验条件下,日粮中添加RSO 能够缓减蛋鸡LPS 诱导的产蛋性能下降和肠道形态结构损伤,其机制是通过TLR4/NF-κB信号通路抑制炎症相关基因表达,提高机体免疫机能。
