基于转录组的三月李及其红肉突变体Expansin基因家族鉴定及分析
2020-12-16方智振林炎娟姜翠翠周丹蓉潘少霖叶新福
方智振,林炎娟,姜翠翠,周丹蓉,潘少霖,叶新福
(福建省农业科学院果树研究所/福建省落叶果树工程技术研究中心,福建 福州 350013)
0 引言
【研究意义】李(Prunus salicinaLindl.)是我国重要的传统果树。李果实美丽诱人,风味独特,且具有很高的营养价值,深受消费者喜爱。成熟的李果实易软化,难以贮藏运输,不利于优质鲜食李产业的发展。细胞壁结构变化是导致成熟果实软化的重要原因[1]。Expansin 作为调节细胞壁的松弛和伸展的一类重要蛋白质,在柿(Diospyros kakiL.‘Fuping Jianshi’)[2]、芒果(Mangifera indicacv. Dashehari)[3]、番荔枝(Annona cherimolaMill)[4]、番茄(Solanum lycopersicum)[5−7]、山番木瓜(Vasconcellea pubescens)[8]和草莓(Fragaria×ananassa)[9]果实成熟和软化中发挥着重要的作用。因此,系统地鉴定和分析李Expansin家族基因对进一步研究Expansin家族基因在李果实成熟过程中的作用具有重要的意义。【前人研究进展】Expansin 是一类广泛存在于植物中的细胞壁酶,它通过破坏细胞壁多糖之间的非共价键参与细胞壁松弛和伸展调节[10-11]。Expansin 由一个多基因家族编码,该家族由EXPA(α-expansin)、EXPB(β-expansin)、EXPLA(expansin-like A)和EXPLB(expansin-like B)等4 个亚家族组成[12]。典型的Expansin 蛋白质在N 端有一个长度为20~30 个氨基酸残基的信号肽和具有多个保守的半胱氨酸残基和HFD 基序的domain1,C 端含有一个与第二组花粉过敏原蛋白同源的多糖结合结构域[13−15]。大量研究表明Expansin 与植物细胞生长[16]、叶片生长发育[17−18]、根系发育[19−20]、果实发育和质地变化[21−26]等诸多生物学过程密切相关。此外,Expansin 还与脱落酸、赤霉素、生长素、油菜素内酯和乙烯等植物激素引起的细胞膨大和细胞壁结构改变有关[27−30]。目前,科研人员已对多种植物的Expansin基因家族进行鉴定与分析,如拟南芥(Arabidopsis thaliana)[31]、烟草(Nicotiana tabacum)[32]、水 稻(Oryza sativa)[33]、小 麦(Triticum aestivum)[14,34]、 番 茄(Solanum lycopersicum)[35]、杨树(Populus trichocarpa)[36]、葡萄(Vitis vinifera)[37]、苹果(Malus×domestica)[38]、桃(Prunus persica)[39]和枣(Ziziphus jujubaMill.)[13]。【本研究切入点】Expansin 在果实发育和成熟中的重要作用已有相关研究,但李Expansin基因家族的鉴定和分析尚未见报道。【拟解决的关键问题】本研究以三月李,以及课题组在前期研究中通过对三月李进行辐射诱变获得的一个果实性状发生显著变化的迟熟突变体[40]为材料,采用生物信息学方法从三月李及其红肉突变体果实成熟过程的转录组数据中鉴定李Expansin家族基因,并对其蛋白质特征、系统进化关系及其在三月李和迟熟突变体果实成熟过程中的表达模式进行比较分析,为研究李Expansin家族基因在果实成熟过程中的功能奠定基础。
1 材料与方法
1.1 数据来源及Expansin家族基因的鉴定
本研究所使用的数据为本课题组获得的三月李(盛花后95、105 和115 d)及其红肉突变体(盛花后95、105、115 和125 d)的转录组数据[41]。拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)和毛果杨(Populus trichocarpa)的Expansin 蛋白质序列从EXPANSIN CENTRAL(http://www.personal.psu.edu/fsl/ExpCentral/)下载。以拟南芥、水稻和毛果杨Expansin 蛋白质序列为查询序列,采用TBtools[42]的BLAST 工具从三月李及其红肉突变体蛋白质数据库中检索Expansin 家族成员。同时,从Pfam 数据库(http://pfam.xfam.org/)下载DPBB_1 和Pollen_allerg_1 结构域的HMM 模型PF03330 和PF01357,并采用HMMER3.0 鉴定含有保守结构域DPBB_1 和Pollen_allerg_1 的蛋白质。
1.2 Expansin 家族分析
Expansin 家族蛋白质生物信息学分析、保守基序分析与多重序列比对、系统进化树的构建、基因的差异表达模式分析参考文献[40]进行。
2 结果与分析
2.1 三月李及其红肉突变体Expansin基因家族的鉴定及蛋白质特性分析
为了鉴定三月李及其红肉突变体的Expansin 家族成员,以拟南芥(35 个)、水稻(58 个)和毛果杨(36 个)Expansin 家族成员的蛋白质序列为查询序列,通过BLAST 分析从三月李及其红肉突变体蛋白质数据库中检索Expansin 蛋白质,初步鉴定出35 个Expansin 蛋白质(E-value<1e−5)。随后,采用隐马尔可夫模型进行Expansin 蛋白质鉴定,去除不含保守结构域DPBB_1 和Pollen_allerg_1 的蛋白质。最终得到33 个Expansin 家族蛋白质(表1)。除了EVM0014217,其余Expansin基因均包含完整的编码序列。李Expansin 蛋白质序列的长度在176(EVM 0014217)~460 aa(EVM0020356),预测蛋白的相对分子量在19.21~51.33 kD,等电点介于4.62~9.83。蛋白质不稳定系数分析结果表明,除了EVM0026183、EVM0000548、 EVM0026545、 EVM0018557、 EVM 0001651 和EVM0020356,其余李Expansin 蛋白质的不稳定系数均小于40,表明大部分李Expansin 蛋白质均为稳定蛋白;平均亲水系数均为负数,表明33 个蛋白质均为亲水蛋白质。亚细胞定位预测结果表明,除了EVM0020356 可分布于质膜与细胞外,其余李Expansin 蛋白质均位于细胞外。
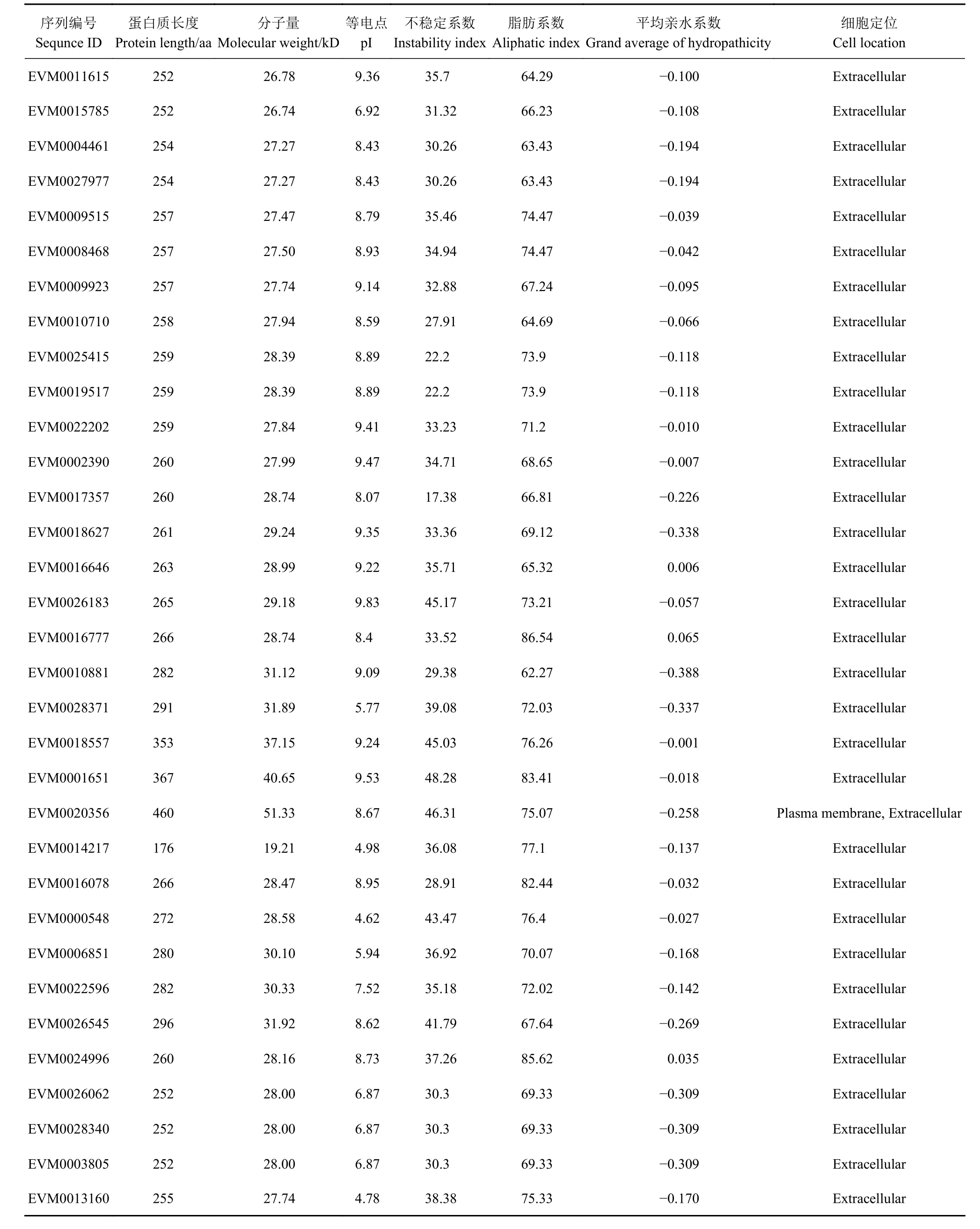
表 1 三月李及其红肉突变体Expansin 蛋白质的理化性质和亚细胞定位Table 1 Physicochemical properties and subcellular localization of Expansins in Sanyueli plum and its red-flesh mutant
2.2 三月李及其红肉突变体Expansin 家族蛋白质进化分析
为了解三月李及其红肉突变体Expansin 家族蛋白质与桃、拟南芥和水稻的进化关系,采用MEGAX软件构建了拟南芥、水稻、桃和李Expansin 家族蛋白质的系统进化树。如图1 所示,拟南芥、水稻、桃和李Expansin 家族蛋白质可分为4 组,分别为EXPA、EXPB、EXPLA 和EXPLB。从图1 可以看出,每个桃Expansin 蛋白质都有与其对应的同源李Expansin蛋白质。EXPA 的李Expansin 家族蛋白质数量最多,共有22 个。其次为EXPB,共含有6 个李Expansin家族蛋白质;仅有1 个李Expansin 蛋白质(EVM 0024996)属于EXPLA 组。EVM0026062、EVM0028340、EVM0003805 和EVM0013160 属于EXPLB 组。

图 1 李、桃、拟南芥和水稻Expansin 家族蛋白质系统进化树Fig. 1 Phylogenetic tree of Expansin family in plum, peach,Arabidopsis, and rice
2.3 三月李及其红肉突变体Expansin 家族蛋白质保守结构域分析
对李Expansin 蛋白质的保守结构域进行分析,发现33 个李Expansin 蛋白质均含有DPBB_1 和Pollen_allerg_1 结构域(图2)。为了进一步了解李Expansin 蛋白质的保守结构域,利用在线软件MEME 对李Expansin家族蛋白质进行分析,结果表明李Expansin 家族蛋白质共含有15 个保守基序(图3)。Expansin 蛋白质含有的基序数量从5~10 个不等。其中,EVM0011615、EVM0028371、 EVM0001651、 EVM0006851、 EVM 0022596 和EVM0026545 含有的基序数量最多(10个),EVM0014217 含有的基序数量最少(5 个)。所有李Expansin家族蛋白质均含有基序3 和5(图3)。除了EVM0014217 之外,其他李Expansin 家族蛋白质均含有基序1 和7(图3)。基序2 是EXPA 成员特有的基序,基序10 为EXPB、EXPLA 和EXPLB 成员特有的基序,基序12 为EXPLA 和EXPLB 成员特有的基序(图3)。含有基序14 的Expansin 蛋白质数量最少,仅3 个EXPLB 成员(EVM0026062、EVM0028340和EVM0003805)含有该基序,EXPA、EXPB 和EXPLA均不含该基序(图3)。

图 2 Expansin 家族蛋白质进化树及保守结构域分析Fig. 2 Phylogenetic tree and conserved domain of Expansin family

图 3 李Expansin 家族蛋白质保守基序及结构域序列Logo Fig. 3 Conserved motifs and domain sequence logo of Expansin family
为了分析家族蛋白质保守结构域的特征,采用DNAMAN7 对Expansin 家族蛋白质进行多重序列比对分析。结果表明大多数Expansin 蛋白质由3 个部分(信号肽、Domain1 和Domain3)组成。大部分EXPA 成 员(EVM0016777 除 外)和 EXPB 成 员(EVM0016078 除外)在Domain1 部分都含有一个保守的HFD 基序,而EXPLA 和EXPLB 在对应位置的特征序列则分别为DFV 和DFI(图4)。HFD 基序两侧存在两个插入片段,分别为EXPA 的α-Insertion和EXPB、EXPLA 和EXPLB 特有的插入片段(图4)。李Expansin 家族蛋白质含有多个保守的半胱氨酸和色氨酸(图4)。

图 4 李Expansin 家族蛋白质的多重序列比对Fig. 4 Multiple sequence alignment analysis on Expansin family
2.4 Expansin家族基因在三月李及其红肉突变体果实成熟过程中的表达模式分析
对33 个Expansin家族基因的表达模式进行分析,发现三月李及其红肉突变体果实成熟过程中仅有9 个Expansin基因存在差异表达,包括7 个EXPA(EVM0015785、EVM0010710、EVM0022202、EVM0002390、EVM0016777、EVM0018557 和EVM 0001651)、1 个EXPB(EVM0016078)和1 个EXPLA(EVM0024996)(图5)。三月李果肉中EVM0015785和EVM0016777 的表达量随果实成熟逐渐降低,但其表达量在红肉突变体果实成熟过程中无显著变化。EVM0010710 和EVM0022202 在成熟三月李果肉中的表达量也显著降低,而在成熟红肉突变体果肉中的表达量明显升高。红肉突变体果肉中EVM0002390的表达量随果实成熟而升高,但其表达量仅在花后105 d的三月李果肉中显著升高,花后115 d的三月李果肉中的表达量又下降至与花后95 d 相当的水平。成熟三月李果肉中EVM0024996 的表达量显著升高,而在红肉突变体果实成熟过程中无显著变化。

图 5 Expansin家族基因在三月李及其红肉突变体果实成熟过程中的表达模式Fig. 5 Expression pattern ofExpansinsin Sanyueli plum and its red-flesh mutant during fruit ripening
3 讨论与结论
Expansin 不仅是在植物生长发育诸多方面都发挥着重要生物学功能的细胞壁蛋白[17,18,20,22−26],还是研究植物发育进化的重要候选基因[34,37,43]。因此,Expansin 获得了广泛的关注。本研究从三月李及其红肉突变体果实成熟过程的转录组中鉴定出33 个Expansin基因,少于玉米(Zea myas, 93 个)[44]、小麦(Triticum aestivum, 87 个)[34]、水稻(Oryza sativa,58 个)[33]、烟 草(Nicotiana tabacum, 52 个)[32]、番茄(Solanum lycopersicum, 43 个)[38]、苹果(Malus×domestica, 41 个)[38]、 杨 树(Populus trichocarpa,36 个)[36]、黄瓜(Cucumis sativus, 35 个)[45]和模式植物拟南芥(Arabidopsis thaliana, 35 个)[31],但多于枣(Ziziphus jujubaMill., 30 个)[13]、 葡 萄(Vitis vinifera, 29 个)[37]及其近缘物种桃(Prunus persica,27 个)[39]。李的近缘物种桃的27 个Expansin 蛋白质在李转录组中均能找到对应的同源蛋白质,可见本研究鉴定的Expansin基因家族完整性较高,但李基因组中Expansin基因的准确数量还有待通过基因组数据分析进一步确定。研究表明不同Expansin 亚家族的成员数量存在很大的差异[32],且大多数情况下EXPA 的数量多于EXPB 的数量[46]。李Expansin 家族的成员组成(22 个EXPA、6 个EXPBs、1 个EXLA和4 个EXLB)与拟南芥(26 个EXPA、5 个EXPB、3 个EXLA 和1 个EXLB)相近,但与水稻(34 个EXPA、19 个EXPB、4 个EXLA 和1 个EXLB)之间存在较大的差异。不同植物间Expansin 家族成员数量和组成的差异可能与植物生长发育和环境适应的不同要求而引起的生物进化有关[35]。
33 个李Expansin基因中有9 个在三月李及其红肉突变体果实成熟过程中存在差异表达,且表达模式各异。Lu 等[35]研究发现不同番茄Expansin家族基因在果实中的表达模式也存在较大的差异。Expansin家族基因在三月李及其红肉突变体果肉中存在多样的表达模式意味着这些基因在果实成熟过程中可能扮演着不同的角色。番茄(Solanum lycopersicum)[47]、草莓(Fragaria×ananassa)[48]、杏(Prunus armeniacaL.)[49]、 桃(Prunus persica)[50]、 香 蕉(Musa acuminate)[51]和山番木瓜(Vasconcellea pubescens)[8]等的研究均表明Expansin的表达与果实软化有关。Palapol 等[24]研究发现DzEXP1和DzEXP2的表达与榴莲果实软化呈正相关,乙烯可促进果肉软化和二者的表达,而1-MCP 则抑制果肉软化和二者的表达。抑制Sl-EXP1的表达可显著提高番茄果实的硬度,而超表达Sl-EXP1则使番茄果实变软[5,25]。这些
研究均表明Expansin直接或间接参与果实软化。成熟的三月李及其红肉突变体果肉均迅速变软。但值得注意的是,本研究鉴定的9 个差异表达Expansin的表达均未呈现与三月李及其红肉突变体果实成熟和软化一致的趋势。Zerpa-Catanho 等[52]在番木瓜上也发现了类似结果,分析了成熟相关基因与采后番木瓜软化的关系,发现Expansin与果实软化不存在相关性。不同植物Expansin在果实成熟软化过程中的功能差异可能与物种有关。Expansin在李果实成熟过程中的具体功能及其是否直接参与李果实软化过程还有待进一步研究。
本研究通过生物信息学分析从三月李及其红肉突变体果实成熟过程的转录组数据中鉴定出33 个Expansin家族基因,并对Expansin 蛋白质的特征和基因表达模式进行分析,为今后进一步研究Expansin家族基因在李果实成熟过程中的功能奠定基础。
