8种乳酸菌发酵荔枝汁-大豆蛋白的氨基酸代谢特征研究
2020-12-10陈梅春郑雪芳王阶平
刘 欣,刘 芸,陈梅春,郑雪芳,陈 峥,王阶平,刘 波
(福建省农业科学院农业生物资源研究所,福建福州 350003)
氨基酸是组成蛋白质的基本单位[1],分为必需氨基酸(如赖氨酸、色氨酸、苯丙氨酸、蛋氨酸、苏氨酸、异亮氨酸、亮氨酸、缬氨酸)和非必需氨基酸(如丙氨酸、精氨酸、天冬氨酸、半胱氨酸、谷氨酰胺、谷氨酸、组氨酸、异亮氨酸、甘氨酸、天冬酰胺、亮氨酸、赖氨酸)[2]。研究表明,食用亮氨酸、缬氨酸和异亮氨酸等支链氨基酸能够加速体能恢复,具有促进肌肉生长、减缓肌肉酸痛等功效[3];补充酪氨酸、苯丙氨酸、色氨酸等芳香族氨基酸可改善人体营养状态,对较轻的肝硬化患者具有降血氨作用[4]。
大豆蛋白富含人体必需的8种氨基酸,经乳酸菌发酵后的豆奶饮品具有一定的保健功效,备受消费者青睐[5]。乳酸菌种类、发酵基料、发酵条件对发酵产品的氨基酸种类和含量产生重要影响,进而影响乳制品的风味和口感,如胡珊等[6]从荔枝体内分离出内生乳酸菌和植物乳杆菌用以发酵荔枝汁,发现前者游离氨基酸保留率为87.3%,略高于后者(84.4%),两株菌在发酵过程中各种氨基酸的种类和含量变化有所不同;王全利等[7]发现发酵时间对必需氨基酸、非必需氨基酸、芳香族氨基酸、支链氨基酸等含量产生重要影响;李理等[8]研究发现利用副干酪乳杆菌发酵复配乳蛋白原料,能够将支链氨基酸含量提高12.5倍;李汴生等[9]用植物乳杆菌LP-115 400B分别发酵橙汁、苹果汁、梨汁、葡萄汁和青瓜汁,发酵前后游离氨基酸变化与基料的挥发性风味物质存在相互转换的关联性。
乳酸菌发酵荔枝汁-大豆蛋白18 h生产的乳酸菌饮品其产品风味和储藏特性表现良好[10],其氨基酸代谢特征的研究未见报道。福建省为荔枝的重要产区,近年来由于市场变动,荔枝销售受到阻碍,作者采用乳酸发酵荔枝汁生产乳酸菌饮品,提高荔枝加工的附加值,促进荔枝产业的健康发展。试验采用荔枝汁-大豆蛋白为培养基,选择8种乳酸菌(植物乳杆菌、德氏乳杆菌、干酪乳杆菌、鼠李糖乳杆菌、短乳杆菌、发酵乳杆菌、嗜酸乳杆菌和嗜热链球菌),以5%接菌量分别接种于荔枝汁-大豆蛋白培养基(1∶1),物料发酵18 h,分析发酵液氨基酸代谢特征、构建氨基酸代谢指数、对乳酸菌发酵液氨基酸含量进行聚类分析,以期为荔枝乳酸饮品的研发提供科学依据。
1 材料与方法
1.1 材料与仪器
8株乳酸菌:植物乳杆菌FJAT-13737(LactobacillusplantarumFJAT-13737)、德氏乳杆菌FJAT-43773(L.delbrueckiiFJAT-43773)、副干酪乳杆菌FJAT-13741(L.paracaseiFJAT-13741)、鼠李糖乳杆菌FJAT-13807(L.rhamnosusFJAT-13807)、短乳杆菌FJAT-43776(L.brerisFJAT-43776)、发酵乳杆菌FJAT-13771(L.fermentumFJAT-13771)、嗜酸乳杆菌FJAT-13772(L.acidophilusFJAT-13772)、嗜热链球菌FJAT-43774(StreptococcusthermophilusFJAT-43774 保存于福建省农业科学院农业生物资源研究所菌株资源库;M17肉汤培养基、MRS肉汤培养基 购自北京陆桥公司。
ZHJH-C1209C超净工作台 上海智城分析仪器制造有限公司;Bluepard隔水式恒温培养箱 上海一恒科技有限公司;L-8900全自动氨基酸分析仪 日立公司。
1.2 实验方法
1.2.1 乳酸菌菌种制备 参考文献[11]中关于种子液制备方法,具体方法如下:嗜热链球菌接种于50 mL M17肉汤培养基,其余7种乳酸菌供试菌株接种于50 mL MRS肉汤培养基,分别于37 ℃恒温培养箱中静置培养36 h,而后标定活菌数,将乳酸菌菌种活菌数含量配制到108CFU/mL,制成为菌种接种液。
1.2.2 乳酸菌荔枝汁-大豆蛋白的发酵 荔枝汁-大豆蛋白培养基的制备:选取九成熟新鲜荔枝果实,用淡盐水浸泡10 min后用清水冲洗荔枝,经人工处理去皮、去核得荔枝果肉,将果肉榨汁去渣,取鲜荔枝汁冷冻备用;挑选表面光泽无霉变的黄豆,用蒸馏水浸泡10 h后清洗,加入干豆质量20倍的纯净水,磨浆去渣得豆浆备用。将荔枝汁和豆浆按1∶1比例混合,加入混合液重量2%的白砂糖,为荔枝汁-大豆蛋白培养基,121 ℃高压15 min灭菌后待用。
荔枝汁-大豆蛋白乳酸菌发酵液的制备:将上述8种乳酸菌种子液,以5%接菌量分别接种于荔枝汁-大豆蛋白培养基中,置于37 ℃温度下培养18 h。
乳酸菌发酵过程活菌数的统计:采用稀释平板计数法,分别配制10-5、10-5、10-6浓度发酵液,吸取液体0.1 mL,进行平板涂布,每个浓度梯度重复3次。将平板倒置于恒温培养箱中37 ℃培养2~3 d。按公式统计培养平板上菌落数:

表1 不同乳酸菌发酵荔枝汁-大豆蛋白前后的氨基酸含量
每毫升样品中活菌数=同一稀释度的菌落平均数×稀释倍数×10。
1.2.3 荔枝汁-大豆蛋白乳酸菌发酵液氨基酸含量的测定 根据《GB 5009.124-2016食品中氨基酸的测定》测定荔枝汁-大豆蛋白乳酸菌发酵前后氨基酸含量。
1.3 数据处理
利用DPS软件进行不同乳酸菌生长与氨基酸代谢相关性分析,分析乳酸菌活菌数与必需氨基酸总量、非必需氨基酸总量、氨基酸总量的相关性;分析乳酸菌活菌数与发酵液必需氨基酸和非必需氨基酸组成结构的相关性。
利用DPS软件进行乳酸菌氨基酸代谢特征的聚类分析。基于乳酸菌的氨基酸聚类分析,以氨基酸为样本,乳酸菌为指标,构建矩阵;基于氨基酸的乳酸菌聚类分析,以乳酸菌为样本,氨基酸为指标,构建矩阵。聚类分析方法,数据不转换,欧氏距离为尺度,用可变类平均法进行系统聚类;根据聚类结果,构建乳酸菌氨基酸代谢特征指数(Amino Acid Metabolism Characteristic Index,AAMCI),AAMCI=EAA+BCAA+AAA/TAA。
2 结果与分析
2.1 发酵液氨基酸代谢特征分析
2.1.1 乳酸菌发酵前后氨基酸含量的测定 试验结果见表1。荔枝汁-大豆蛋白培养基乳酸菌发酵前后物料共检测到17种氨基酸,其中必需氨基酸(EAA)8种,非必需氨基酸(NEAA)9种,支链氨基酸(BCAA)3种,芳香族氨基酸(AAA)2种;发酵前培养基中氨基酸总量(582.78±4.16) mg/100 g显著高于各乳酸菌发酵后发酵液中氨基酸总量(P<0.05);不同乳酸菌发酵18 h后发酵液的氨基酸总量存在差异,乳酸菌发酵后发酵液氨基酸含量最高的菌为德氏乳杆菌FJAT-43773(481.52±6.25 mg/100 g),最低的为副干酪乳杆菌FJAT-13741(365.96±5.78 mg/100 g);除了LAB1(植物乳杆菌FJAT-13737)与LAB5(短乳杆菌FJAT-43776)、LAB6(发酵乳杆菌FJAT-13771),LAB2(德氏乳杆菌FJAT-43773)与LAB4(鼠李糖乳杆菌)、LAB7(嗜酸乳杆菌FJAT-13772),LAB3(副干酪乳杆菌FJAT-13741)与LAB8(嗜热链球菌FJAT-43774)之间氨基酸总量(TAA)差异不显著(P>0.05)外,其余乳酸菌之间发酵后氨基酸总量差异显著(P<0.05)。
2.1.2 乳酸菌发酵后物料氨基酸平均含量的变化 基于表1统计结果见表2。将8种乳酸菌发酵18 h后发酵液氨基酸含量作平均值与发酵前培养基中氨基酸含量进行比较,结果表明,总体上,经乳酸菌发酵培养基中的氨基酸除了脯氨酸外,其余氨基酸呈下降趋势,如含量最高的谷氨酸,发酵前含量为(108.03±5.49) mg/100 g,发酵后含量平均值为(89.98±8.17) mg/100 g,下降了16.70%;含量最低的胱氨酸,发酵前含量为(2.96±0.11) mg/100 g,发酵后含量平均值为(2.08±0.36) mg/100 g,下降了29.83%;发酵前后培养基氨基酸下降率最高的是精氨酸(42.17%),最低的为蛋氨酸(14.91%),总氨基酸(TAA)含量下降了26.31%。乳酸菌发酵的结果产生的氨基酸代谢消耗了培养基中的氨基酸,且不同种类氨基酸的消耗水平差异明显。

表2 乳酸菌发酵荔枝汁-大豆蛋白物料后氨基酸平均含量
2.1.3 不同乳酸菌发酵液特征氨基酸含量的比较 分析结果见图1。从发酵液中必需氨基酸(蛋氨酸+组氨酸+异亮氨酸+苏氨酸+苯丙氨酸+缬氨酸+亮氨酸+赖氨酸)含量看,不同乳酸菌之间存在差异,含量最高的为LAB2(德氏乳杆菌FJAT-43773,169.51 mg/100 g),最低的为LAB3(副干酪乳杆菌FJAT-13741,118.78 mg/100 g);除了LAB1和LAB6,LAB2和LAB7,LAB3和LAB8,LAB4和LAB5之间无显著差异外(P>0.05),其余存在显著差异(P<0.05)。
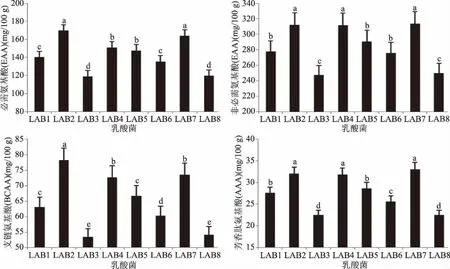
图1 不同乳酸菌发酵液特征氨基酸含量比较
从发酵液中非必需氨基酸(胱氨酸+酪氨酸+甘氨酸+精氨酸+丝氨酸+脯氨酸+天门冬氨酸+丙氨酸+谷氨酸)含量看,不同乳酸菌之间存在差异,含量最高的为LAB7(嗜酸乳杆菌FJAT-13772,313.68 mg/100 g),最低的为LAB3(副干酪乳杆菌FJAT-13741,247.17 mg/100 g);除了LAB2和LAB4及LAB7,LAB1和LAB5及LAB6,LAB3和LAB8之间差异不显著外(P>0.05),其余存在显著差异(P<0.05)。
从发酵液中支链氨基酸(异亮氨酸+缬氨酸+亮氨酸)含量看,不同乳酸菌之间存在差异,含量最高的为LAB2(德氏乳杆菌FJAT-43773,78.26 mg/100 g),含量最低的为LAB3(副干酪乳杆菌FJAT-13741,53.52 mg/100 g);除了LAB1和LAB5,LAB3和LAB8,LAB4和LAB7之间差异不显著外(P>0.05),其余存在显著差异(P<0.05)。
从发酵液中芳香族氨基酸(苯丙氨酸+酪氨酸)含量看,不同乳酸菌之间存在差异,含量最高的为LAB7(嗜酸乳杆菌FJAT-13772,33.00 mg/100 g),最低的为LAB3(副干酪乳杆菌FJAT-13741,22.46 mg/100 g);除了LAB1和LAB5,LAB2、LAB4和LAB7,LAB3和LAB8之间差异不显著外(P>0.05),其余存在显著差异(P<0.05)。
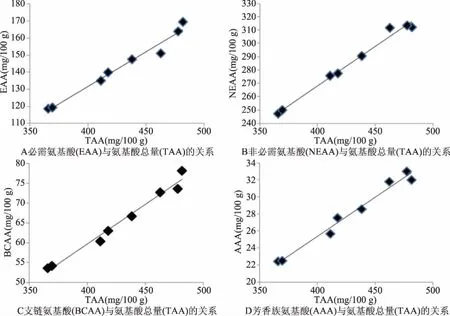
图2 发酵液中特征氨基酸与氨基酸总量的关系
2.1.4 特征氨基酸与氨基酸总量的关系 分析结果见图2。乳酸菌发酵液中特征氨基酸变化与氨基酸总量之间存在着线性关系;必需氨基酸(EAA)含量与氨基酸总量呈线性关系(P<0.01),方程为y=0.4071x-31.041(R2=0.9740)(图2A);非必需氨基酸(NEAA)含量与氨基酸总量呈线性关系(P<0.01),方程为y=0.5929x+31.041(R2=0.9876)(图2B);支链氨基酸(BCAA)含量与氨基酸总量呈线性关系(P<0.01),方程为y=0.2006x-20.542(R2=0.9800)(图2C);芳香族氨基酸(AAA)含量与氨基酸总量呈线性关系(P<0.01),方程为y=0.0915x-11.213(R2=0.9797)(图2D)。研究表明,特征氨基酸变化与氨基酸总量存在着一定比例,如必需氨基酸含量高时,其氨基酸总量也相应提高,如必需氨基酸含降低时,其氨基酸总量也相应降低;同样的规律也出现在非必需氨基酸、支链氨基酸和芳香族氨基酸。
2.2 发酵液氨基酸代谢指数构建
2.2.1 必需氨基酸与非必需氨基酸的相互关系 分析结果见图3。不同乳酸菌发酵液必需氨基酸(EAA)含量与非必需氨基酸含量(NEAA)呈显著线性关系(P<0.05),方程为y=0.6656x-46.391(R2=0.9270)(图3)。不同乳酸菌的必需氨基酸(EAA)与非必需氨基酸(NEAA)的比值(EAA/NEAA)差异显著(图4),其中比值最高的为德氏乳杆菌FJAT-43773(54.32%),最低的为嗜热链球菌FJAT-43774(47.82%)。

图3 必需氨基酸与非必需氨基酸的关系

图4 不同乳酸菌必需氨基酸和非必需氨基酸比值
2.2.2 氨基酸代谢特征指数的构建 构建的乳酸菌氨基酸代谢特征指数如表3。氨基酸代谢特征指数(AAMCI)代表了功能性氨基酸:必需氨基酸(EAA)、支链氨基酸(BCAA)、芳香族氨基酸(AAA)的总和占氨基酸总量的比例。指数高代表功能氨基酸占比高;分析结果表明,不同乳酸菌发酵液氨基酸代谢指数差异不显著(P>0.05),嗜热链球菌FJAT-43774氨基酸代谢特征指数最低为53.10%,德氏乳杆菌FJAT-43773最高为58.10%,说明不同乳酸菌通过功能氨基酸代谢平衡形成氨基酸代谢指数的相对稳定。
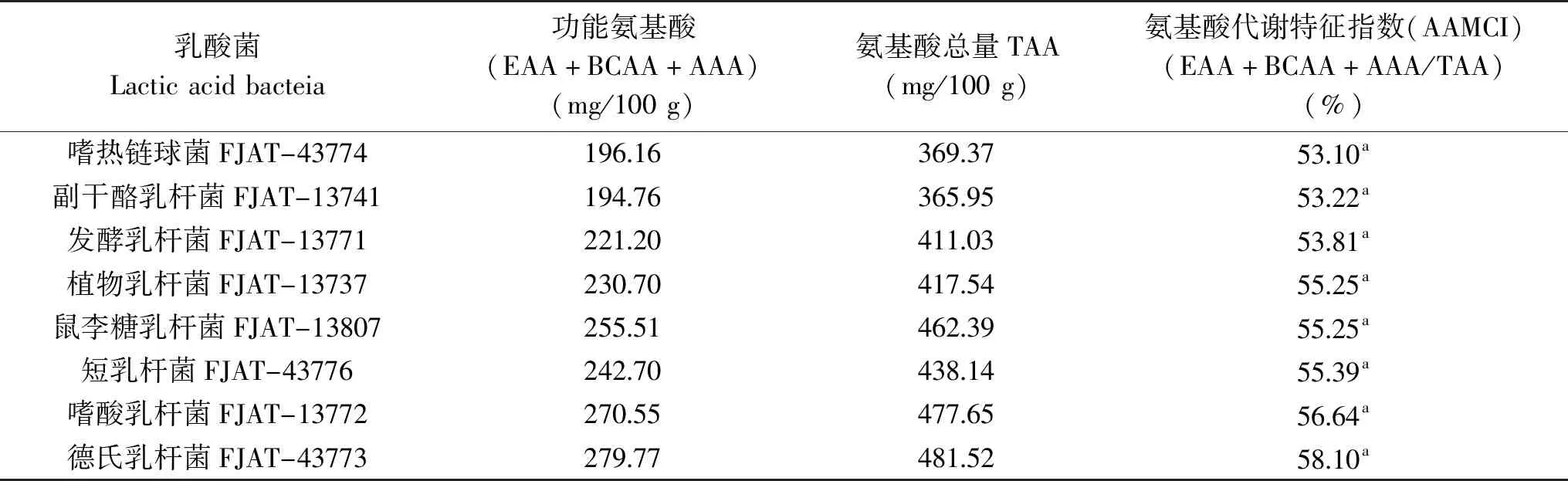
表3 乳酸菌氨基酸代谢特征指数构建
2.3 乳酸菌发酵液氨基酸种类分布聚类分析
2.3.1 基于乳酸菌发酵液的氨基酸代谢分布 不同的氨基酸在乳酸菌发酵液中的分布差异显著,以表1为数据矩阵,氨基酸为样本,乳酸菌为指标,数据不转换,欧氏距离为尺度,用可变类平均法进行系统聚类;分析结果见图5和图6;结果表明,依据乳酸菌发酵液中氨基酸含量将氨基酸分为3组。

图5 乳酸菌发酵液氨基酸分布聚类分析(欧氏距离)
第1组低含量氨基酸组,氨基酸含量范围在1~26 mg/100 g,包括了11种氨基酸,其中6种必需氨基酸和5种非必需氨基酸,即蛋氨酸、组氨酸、异亮氨酸、苏氨酸、苯丙氨酸、缬氨酸、精氨酸、胱氨酸、酪氨酸、甘氨酸、丝氨酸。含量最低的氨基酸为胱氨酸,在植物乳杆菌FJAT-13737、德氏乳杆菌FJAT-43773、副干酪乳杆菌FJAT-13741、鼠李糖乳杆菌FJAT-13807、短乳杆菌FJAT-43776、发酵乳杆菌FJAT-13771、嗜酸乳杆菌FJAT-13772、嗜热链球菌FJAT-43774发酵液中含量分布分别为2.83、2.56、1.51、2.33、2.23、1.87、2.20、1.84 mg/100 g;含量最高的为丝氨酸,在上述乳酸菌发酵液中含量分布分别为21.72、25.96、18.71、25.33、23.34、21.54、24.58、19.53 mg/100 g。
第2组中含量氨基酸组,氨基酸含量范围在26~57 mg/100 g,包括了5种氨基酸,2种必需氨基酸,3种非必需氨基酸,即亮氨酸、赖氨酸、脯氨酸、天门冬氨酸、丙氨酸。含量最低的氨基酸为亮氨酸,在植物乳杆菌FJAT-13737、德氏乳杆菌FJAT-43773、副干酪乳杆菌FJAT-13741、鼠李糖乳杆菌FJAT-13807、短乳杆菌FJAT-43776、发酵乳杆菌FJAT-13771、嗜酸乳杆菌FJAT-13772、嗜热链球菌FJAT-43774发酵液中含量分布分别为27.46、34.16、22.94、31.63、28.94、26.76、33.12、23.19 mg/100 g;含量最高的为丙氨酸,在上述乳酸菌发酵液中含量分布分别为49.32、53.04、46.44、53.98、52.52、52.71、56.68、47.15 mg/100 g。
第3组高含量氨基酸组,氨基酸含量范围在79~98 mg/100 g,包括了1种非必需氨基酸,即谷氨酸,在植物乳杆菌FJAT-13737、德氏乳杆菌FJAT-43773、副干酪乳杆菌FJAT-13741、鼠李糖乳杆菌FJAT-13807、短乳杆菌FJAT-43776、发酵乳杆菌FJAT-13771、嗜酸乳杆菌FJAT-13772、嗜热链球菌FJAT-43774发酵液中含量分布分别为88.41、96.83、79.15、97.13、92.40、87.55、97.60、79.23 mg/100 g。

图6 基于发酵液氨基酸分布的乳酸菌聚类分析(欧氏距离)
2.3.2 基于氨基酸代谢的乳酸菌聚类分析 乳酸菌发酵过程氨基酸的组成差异显著,根据氨基酸含量对乳酸菌进行聚类,分析乳酸菌氨基酸代谢特性。以表1为数据矩阵,乳酸菌为样本,氨基酸为指标,数据不转换,欧氏距离为尺度,用可变类平均法进行系统聚类;分析结果见图6。结果表明,依据氨基酸组成可将乳酸菌为3类。第1类包含了3种乳酸菌,即短乳杆菌FJAT-43776、发酵乳杆菌FJAT-13771、植物乳杆菌FJAT-13737,其特征为:较高的氨基酸总量,平均值高达422.23 mg/100 g;表明该类乳酸菌中含量氨基酸代谢水平较高。第2类包含了3种乳酸菌,即德氏乳杆菌FJAT-43773、鼠李糖乳杆菌FJAT-13807、嗜酸乳杆菌FJAT-13772,其特征为:中等的氨基酸总量,平均值为348.82 mg/100 g;表明该类乳酸菌中含量氨基酸代谢水平居中。第3类包含了2种乳酸菌,即副干酪乳杆菌FJAT-13741、嗜热链球菌FJAT-43774,其特征为:较低的氨基酸总量,平均值为265.01 mg/100 g;表明该类组乳酸菌中含量氨基酸代谢水平较低。
3 讨论
乳酸菌发酵植物原料前后氨基酸含量变化与乳酸菌种类、培养基料特性等密切相关:培养基本身氨基酸含量高的,经乳酸菌发酵后氨基酸含量表现出下降,培养基含量氨基酸低的,发酵后能提升氨基酸含量。苏能能等[12]利用乳酸菌发酵桑葚浆,其游离氨基酸总含量下降了3.6倍,但蛋氨酸和谷氨酸含量变化不显著;杨冲[13]采用乳酸菌发酵南丰蜜桔汁,发酵后氨基酸总量减少了13.9%;邵齐等[14]利用乳酸菌发酵对芒果浆,发现发酵前后氨基酸总量变化不显著(P>0.05);施帅等[15]用乳酸菌发酵玫瑰花,发现游离氨基酸发酵后含量增加了16.46%;菅田田等[16]报道麸皮高酸海棠果饮料乳酸菌发酵后氨基酸含量增加27.5%;王振斌等[17]报道葛根汁乳酸菌发酵前后必需氨基酸总量比葛根汁增加了65.58%;魏一星等[18]报道了小麦胚芽乳酸菌发酵前后总游离氨基酸的含量提高了13.64倍。本研究中供试的8种乳酸菌发酵过程产生17种氨基酸,发酵前培养基氨基酸总量达582.78 mg/100 g,发酵后下降了26.31%。
乳酸菌发酵产生的必需氨基酸、支链氨基酸、芳香族氨基酸等都属于功能性氨基酸,本研究发现尽管不同乳酸菌发酵液必需氨基酸与非必需氨基酸的比值差异显著,但乳酸菌通过功能氨基酸代谢平衡形成氨基酸代谢指数的相对稳定。在诸多氨基酸中,支链氨基酸如亮氨酸、缬氨酸和异亮氨酸特别引人关注,这类氨基酸对人体具有显著的营养保健作用[3]。乳酸菌发酵产生的支链氨基酸含量与发酵时间密切相关,王剑飞等[19]研究发现产蛋白酶乳酸菌发酵乳清蛋白5 h后开始产生支链氨基酸,其含量在26 h达到最大值(31.26 mg/100 g),随后缓慢减少。本研究中,不同乳酸菌产生的支链氨基酸总量有所差异,植物乳杆菌FJAT-13737、德氏乳杆菌FJAT-43773、副干酪乳杆菌FJAT-13741、鼠李糖乳杆菌FJAT-13807、短乳杆菌FJAT-43776、发酵乳杆菌FJAT-13771、嗜酸乳杆菌FJAT-13772、嗜热链球菌FJAT-43774的支链氨基酸总量分别为63.09、78.26、53.52、72.79、66.72、60.34、73.58、54.15 mg/100 g,在植物基原料乳酸菌发酵中属于支链氨基酸总量较高的。
乳酸菌氨基酸代谢能力与乳酸菌的种类密切相关,Simova等[20]发现瑞士乳杆菌MP12(L.helveticus)发酵牛奶16 h氨基酸总量达53.38 mg/100 g,是保加利亚乳杆菌HP1(L.bulgaricus)的3.7倍;双歧杆菌BL1发酵马铃薯酸奶氨基酸总量达136.64 mg/100 g[21];植物乳杆菌NCU137发酵桑葚浆60 h氨基酸总量19.12 mg/100 g[12]。已有研究表明,乳酸菌生长过程消耗和产生氨基酸同时发生,处于动态平衡[22],本研究的结果也显示了这一趋势,不同乳酸菌发酵终点发酵液必需氨基酸、非必需氨基酸、支链氨基酸、芳香族氨基酸与氨基酸总量呈线性相关。本研究进一步对乳酸菌的代谢特征进行聚类分析,发现8种乳酸菌的氨基酸代谢特征分为3组,即高含量组1种氨基酸、中含量组5种氨基酸、低含量组11种氨基酸,中含量氨基酸组主导着氨基酸总量的变化。根据不同氨基酸组可将8种乳酸菌分为3类,第1类包含了3种乳酸菌,其特征为氨基酸总量平均值最高达422.23 mg/100 g,氨基酸代谢特征指数54.82%。第2类3种乳酸菌其特征为氨基酸总量平均值最高达348.82 mg/100 g,氨基酸代谢特征指数56.66%。第3类2种乳酸菌,其特征为氨基酸总量平均值最高达265.01 mg/100 g,氨基酸代谢特征指数53.16%。研究为高品质乳酸饮品的研发提供基础数据支撑。
4 结论
本研究利用不同乳酸菌发酵荔枝汁-大豆蛋白物料,于发酵前后共检测到氨基酸种类17种,发酵后氨基酸含量从582.79 mg/100 g下降为427.97 mg/100 g。该物料经不同乳酸菌发酵后的氨基酸总量差异显著,副干酪乳杆菌FJAT-13741(365.95 mg/100 g)<嗜热链球菌FJAT-43774(369.36 mg/100 g)<发酵乳杆菌FJAT-13771(411.03 mg/100 g)<植物乳杆菌FJAT-13737(417.54 mg/100 g)<短乳杆菌FJAT-43776(438.14 mg/100 g)<鼠李糖乳杆菌FJAT-13807(462.38 mg/100 g)<嗜酸乳杆菌FJAT-13772(477.65 mg/100 g)<德氏乳杆菌FJAT-43773(481.52 mg/100 g),利用上述乳酸菌发酵荔枝汁-大豆蛋白物料,获得的支链氨基酸总量高于常规植物基原料。构建了氨基酸代谢特征指数(AAMCI),用以评价乳酸菌饮品发酵过程功能性氨基酸占氨基酸总量的比值,其中比值最高的为德氏乳杆菌FJAT-43773(54.32%),最低的为嗜热链球菌FJAT-43774(47.82%)。研究结果为荔枝乳酸菌饮品研制菌株的筛选提供理论依据。
