红锥2代种子园半同胞子代苗生长差异分析*
2020-12-07张卫华杨晓慧薄文浩
徐 放 张卫华 杨晓慧 薄文浩 潘 文
杨会肖1 廖焕琴1 徐 斌1 朱报著1 王裕霞1
(1. 广东省森林培育与保护利用重点试验室/广东省林业科学研究院, 广东 广州 510520; 2. 北京林业大学 生物科学与技术学院,北京 100083)
红锥Castanopsis hystrix 为壳斗科Fagaceae 锥属常绿乔木,广泛分布于我国南方大部分地区,生长速度快,材性优良,是华南地区重要的乡土珍贵阔叶用材树种和高效多用途树种,具有极好的环境适应性,已被广东、福建、广西等省区列为重点发展的树种[1-3];红锥种子富含淀粉和多种次生代谢物,是提取淀粉和化工原料的良好材料,含有大量淀粉的种子也为野生动物提供了大量的食物,有利于维系稳定的森林生态[4];枝叶茂盛耐荫蔽,可与松树、杉木等进行混合经营,具有良好的经济效益[5-7]。红锥的轮伐期为25~30 年,优良品种对木材质量、产量和提升林地经营收益至关重要[8],而目前我国红锥选育工作较为落后,为了改善红锥研究的局面,推进红锥新品种的选育工作,本研究对红锥2 代种子园中收集的种子构建14 个半同胞家系,开展苗期8 个重要生长测定并开展优良家系的初步评选,旨在为后续的红锥优良品系的选育提供重要的参考。
1 材料与方法
1.1 试验地概况
本试验地位于广东省林业科学研究院苗圃(23°36′5″N、113°9′35″E),属亚热带季风性气候类型,季节性非常明显,近10 年平均年降水量2 015 mm,平均相对湿度69%。每年4 月到6 月为雨季,雨季平均月降雨量可达200 mm 以上,7—9 月气温较高,平均最高气温可达32.8 ℃。平均年日照时数1 636.8 h,年平均气温22.3 ℃。
1.2 试验材料
于2017 年12 月采集广州市龙眼洞林场内的红锥2 代种子园内14 个家系单株的种子,采集全部果实后利用日光进行干燥处理,果实开裂后快速收集所有种子,避免种壳因过度干燥开裂造成种子失死亡。利用水对种子的饱满程度进行分选,选择沉入水中的种子,浸泡过夜后播种在以黄心土为基质的苗床上,覆盖2 cm 厚的河沙,种子发芽后至5 cm 高后移至7 cm×12 cm 的无纺布育苗袋中,基质为等体积的黄心土和泥炭土混合基质。在育苗过程中每45 天施用水溶性复合肥(多普瑞斯,N:P:K=19%:19%:19%)1 次,单次用量为0.2 g/株。
1.3 苗期表型测定与分析
于2019 年5 月初,随机选取14 个家系中的各30 个单株,设置3 次重复,每个重复10 个单株,开展生长表型分析。利用激光测距仪测定株高;数显游标卡尺测定地径;叶绿素含量利用SPAD-502 plus 叶绿素测量仪测定,每个单株测定4 个不同方向和位置的叶片,获得每个叶片的SPAD 值;统计各单株的叶片数量,并摘取每个单株的10 个叶片,利用扫描仪扫描后获取叶片图像,利用Digmizer v4.0 软件中长度和面积计算模块统计各叶片的长度、宽度和叶面积,并计算叶片长宽比。测量数据利用Excel 进行整理,利用R 软件[9]中的agricolae进行方差分析和多重比较,利用R 软件自编代码完成相关系数计算。方差分析计算公式:
yij= μ+ ∂i+ βi+ εij,其中 yij为单株表型实际测量值,μ 为总体平均值, ∂i为重复效应, βi为家系间遗传差异, εij为误差。相关系数计算公式:
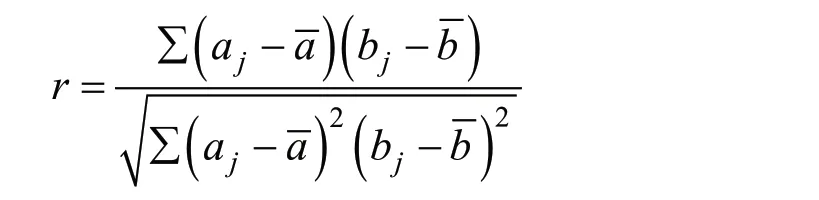
其中a 和b 分别表示不同表型的平均值。
2 结果与分析
2.1 不同半同胞家系间各表型方差分析
对不同家系间株高、地径、叶片数、SPAD值、叶片长度、叶片宽度、叶片长宽比和叶面积等8 个性状进行方差分析,结果显示在不同的半同胞家系间,以上8 个性状均存在极显著差异(P<0.01 或P<0.001),该结果说明,在种植和管理条件一致的前提下,不同家系的差异导致了生长表型的显著差异(表1)。8 个性状总体变异系数在13.8%至59.7%之间,SPAD 值在所有试验样本中差异最小,叶片数量变化幅度最大,株高和地径的总体变异系数分别为32.2%和24.4%。
2.2 不同半同胞家系生长差异性分析
14 个半同胞家系的株高、地径、叶绿素含量(SPAD 值)和叶片数调查结果和多重比较结果见表2。各家系平均株高28.6~56.8 cm,标准差为9.4 cm,总体平均值为41.3 cm,其中C11 家系平均株高最高,C13 家系次之,C09 家系长势最弱,株高最低;株高变异系数最小家系为C03(12.8%),该家系株高相对整齐,家系内差异最小,C06 家系株高变异系数最大,为39.9%;各家系地径3.7~6.3 mm,标准差为0.6 mm,平均地径为4.9 mm,变异系数为7.9%至40.2%。各家系SPAD 值在33.1 至43.9 之间,变异系数为7.4%~16.1%。各家系最小平均叶片数量为18.0 片(C04),最多为84.2 片(C13),平均变异系数为39.7%(表2)。
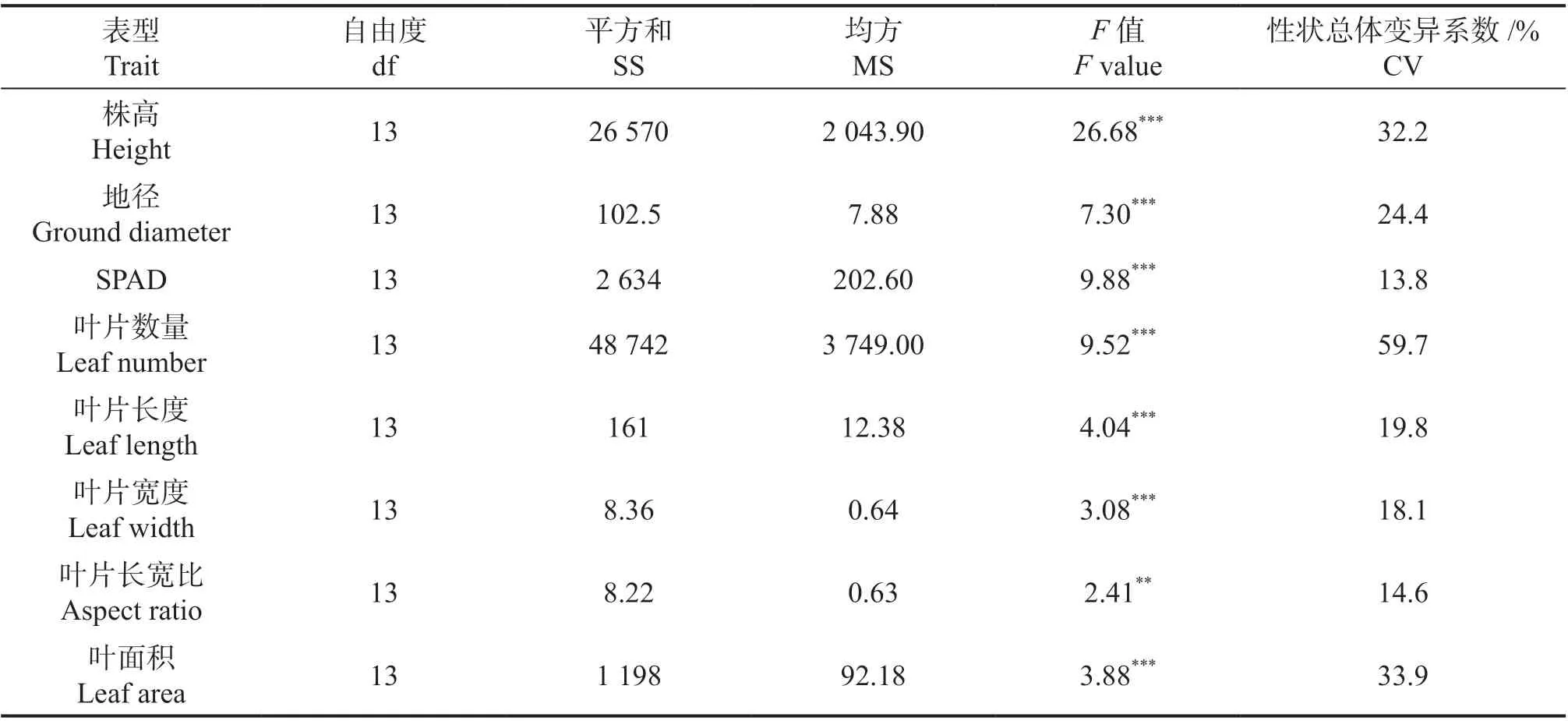
表1 红锥14 个家系不同性状方差分析结果Table 1 Traits ANOVA results of C. hystrix families

表2 红锥不同家系株高、地径、叶片数和叶绿素含量及变异系数Table 2 Plant height, ground diameter, leaf number, chlorophyll content and coefficient of variation of different C. hystrix families
2.3 不同半同胞家系叶片形态差异性分析
不同家系叶片表型测量和计算结果如表3 所示,14 个半同胞家系总体叶片平均长度为9.0 cm,平均叶片长度最小的家系为C09(8.1 cm),平均叶片长度最大的家系为C14(10.1 cm),总体标准差为0.66 cm;叶片宽度最小的家系为C09,最大家系为C07 和C14,变化区间为2.3 cm 至2.8 cm,总体平均值为2.6 cm,标准差为0.14 cm;叶片长宽比最小值为3.2,最大值为3.8,平均为3.6,标准差为0.15;平均叶面积最小的家系为C09(11.3 cm2),最 大 的 家 系 为C14(17.3 cm2),各 家 系叶面积平均值为15.0 cm2,叶面积标准差为1.80 cm2。叶片长度、叶片宽度、叶片长宽比和叶面积在不同家系间的平均变异系数分别为18.6%、17.5%,13.8%和32.7%。
2.4 不同表型间相关性分析
对8 组生长表型进行相关性分析,相关系数结果如表4 所示。其中株高与地径(0.63)和叶片宽度与叶面积(0.60)2 组生长表型两两性状之间存在显著的正相关(P<0.05);株高与叶片数量(0.76)、地径与叶片数量(0.75)、叶片长度与叶片宽度(0.94)、叶片长度与长宽比(0.96)和叶片宽度与长宽比(0.85)等5 组生长表型两两性状之间存在极显著的正相关(P<0.01)。其余各生长表型性状间相关系数均未达到显著水平,相关系数为-0.21 至0.31 之间。
2.5 苗期表现优良家系初选
鉴于用材树种新品种选育中所关注的最重要性状为生长量,即株高和地径两个表型,另外光合能力相关表型如叶绿素含量、叶片数量和叶面积等均会对生长量产生间接影响,但根据本文2.4中计算结果可知,叶片数量与株高、叶片数量与地径呈显著正相关,叶绿素含量、叶片长宽和叶面积等性状与株高、地径相对独立,无明显关联。因此,本研究选择株高和地径两个表型,作为苗期生长优良家系初选关注的生长表型,以两表型在14 个家系间的平均值与标准差之和作为筛选标准;家系C11(56.8 cm)、C13(53.1 cm)C10(52.4 cm)的平均株高大于总体平均株高(41.3 cm)与标准差(9.4 cm)之和50.7 cm,共有4 个家系的平均地径高于总体地径(4.9 mm)和标准差(0.6 cm)之和5.5 mm,分别为C02(6.3 mm)、C13(5.8 mm)和C12(5.6 mm)。其中C13 家系株高和地径两个表型均超过平均值与标准差之和的筛选标准;另外,C10 和 C11 两个家系平均地径虽然略低于筛选标准,但其平均株高均大于筛选标准;C12 家系的平均株高为49.7 cm,接近筛选标准,C02 家系的地径最大(6.3 mm),其平均株高(40.4 cm)接近14 个家系的平均株高。综上所述,共评选C02、C10、C11、C12 和C13 共5 个家系为苗期生长优良的家系,可作为后续新品种选育需关注的重要遗传资源。以上5 个家系,平均株高为50.12 cm,与2 代种子园中14 个半同胞家系的平均株高(41.3 cm)相比提升了21.3%;平均地径为5.54 mm,高出14个半同胞家系总体平均地径(4.9 mm)13.6%;在 所 选 取 的C02、C10、C11、C12 和C13 家 系中,各家系株高的变异系数分别24.3%、18.1%、16.5%、21.4% 和18.6%,平均值为19.4%,5 个家系的变异系数均低于21.9%的总体平均值;5个家系地径的变异系数为18.9%、11.3%、15.5%、18.7%和16.5%,5 个家系的平均值为16.4%,低于18.1%的平均值,以上数据表明评选出的5 个苗期表现较好的家系,其子代苗木的生长也具有较好的一致性。

表3 红锥不同家系叶片形态测量结果Table 3 Leaf morphology measurement results of different C. hystrix families

表4 红锥不同表型间相关分析Table 4 Correlation analysis between different C. hystrix traits
3 结论与讨论
本研究的结果表明,红锥2 代种子园中的14个半同胞家系在幼苗期的生长表现出明显差异,根据方差分析的结果可知,苗期生长表型差异的主要因素为半同胞家系间的遗传差异,这也与红锥种源遗传差异分析获得的结果一致[10-11],该结果也证明了现阶段红锥具有很好的育种潜力,可以通过优良家系无性系化的组培繁育或建立改良种子园等方式实现遗传改良,进而提升红锥苗木质量和红锥木材产量。如家系C10,其株高变异系数为18.1%,地径变异系数为11.3%,后者已经接近以往杨树Populus tomentosa 无性系选育研究中部分无性育苗过程中单株生长量的变异系数(10%)[12];农林业生产中,种苗的一致性是产品最为重要的经济指标之一[13]。当前,红锥主要利用种子园收获的种子进行繁育和造林,通过苗期生长表现优良家系的评选,提升苗木生长量和一致性,可以更好的实现种苗生产,便于育苗期和造林初期进行水肥控制,提升造林成果率,降低营林成本,显著提升全周期林地经营的收益[2]。本研究完成的红锥2 代种子园中14 个半同胞家系苗期生长量差异分析,可以为后续的育种提供重要的参考。
通过8 个生长表型的相关系数的计算发现呈显著正相关的表型主要有:(1)株高-地径-叶片数和(2)叶片长度-叶片宽度,其中叶片长宽与生长表型并无显著关联,方差分析结果显示所有表型的差异的主要来源为不同家系的差异,因此选择株高与地径作为苗期生长优良家系的评选表型。
由于红锥生长周期较长,轮伐期25~30 年,对本研究评选出的苗期生长表现优良的材料造林后生长量进行长期跟踪调查,需进一步分析其苗期生长、早期生长和后期生长的相关性,开发有效的早期选择方法。随着红锥组培技术的不断深入研究,未来红锥种苗生产可采用种子园繁育与组培无性繁育相结合的方式进行[14-17],从已有的优良家系子代测定林中不断评选优良单株,通过组培无性繁殖的方式进行优质种苗生产,同时选择优良无性系进行人工控制授粉以创制优良新种质,不断推进红锥育种进程。
