藻蓝色素蛋白抑制A549细胞活性机制的miRNA 转录组学分析
2020-11-27刘媛璞张佳雯王成涛
郝 帅,李 爽,王 静,刘媛璞,赵 磊,张佳雯,王成涛
(北京市食品营养与人类健康高精尖创新中心,北京市食品添加剂工程技术研究中心,北京工商大学食品与健康学院,北京 100048)
食用藻蓝色素蛋白是一种安全的天然功能性食品,除了作为添加剂用于多种食品着色以外[1],近年来,越来越多的证据表明,藻蓝蛋白还具有抗氧化、抗衰老、调节免疫、抗肿瘤、抗炎等多种生物学功能[2-6]。深入研究藻蓝蛋白的生物活性及其调控机制,能够为其深度开发和利用提供重要的理论参考。
肺癌是目前我国第一大肿瘤疾病。工业废气、汽车尾气的大量排放所造成的环境问题日益严重,是我国肺癌发生率逐年提高的重要诱导因素[7]。同时,随着我国人口老龄化程度的加剧,自1998年到2015年,我国的肺癌发病率和死亡率始终保持不断上升的趋势,肺癌已经成为危害我国居民健康的最主要的恶性肿瘤[8-9]。肺癌的病理分型可以分为小细胞肺癌和非小细胞肺癌,其中小细胞肺癌主要以H446、H524、H20等细胞系为典型代表,发病比例较低;非小细胞肺癌则主要包括肺鳞癌(H226等)、肺腺癌(LTEP-a2、A549、H1299等)和大细胞肺癌(H460等)三大亚类[10-12]。在近年来被诊断的肺癌病例中,约85%属于非小细胞肺癌[13-14]。已有研究表明,藻蓝蛋白对多种非小细胞肺癌具有生长抑制作用,并且能够促进肺癌细胞的凋亡和体外迁移[15-18]。但是,目前大部分的研究仅针对于藻蓝蛋白与单一细胞蛋白或宏观的信号通路之间的关系,与细胞内小分子的调控作用尚不明确。因此,深入探究藻蓝蛋白对非小细胞肺癌的调控机理,可为其预防、诊断甚至治疗提供新的思路和依据。
非编码小RNA(microRNA,miRNA)是非编码RNA的一类,大约由20 个左右的碱基组成高保守调控型的内源性RNA[19-20],在动植物细胞中参与基因的转录后表达调控作用,能够通过靶向结合到生物体mRNA的3’非翻译区上引起mRNA的降解或者翻译抑制作用[21]。研究表明miRNA能够作为原癌因子或抑癌因子在多种肿瘤细胞中参与调控作用,例如,miR-137能够通过下调PAK2蛋白的表达而抑制恶性黑色素瘤细胞的增殖[22],miR-494能够靶向抑制CDC20的表达而诱导神经胶质瘤细胞的凋亡[23];此外,miR-125b则能够分别在恶性血液病肿瘤和实体瘤中发挥原癌因子和抑癌因子的作用[24-25]。因此,miRNA对于肿瘤的发生、发展和调控具有至关重要的作用。目前,虽然在肺癌中也发现存在许多诸如miRNA-148a[26]、miR-21[27]、miR-17-92[28]等miRNA的调节,但藻蓝蛋白抑制非小细胞肺癌过程中所参与的miRNA尚不明确,相关文献也鲜有报道。本研究以典型的非小细胞肺癌A549细胞系为模型,在藻蓝蛋白处理后,采用高通量非编码小RNA转录组技术(miRNA-seq)分析潜在的差异miRNA,进一步深入揭示藻蓝蛋白对非小细胞肺癌的调控机制,为功能性藻蓝蛋白的开发和利用提供理论参考。
1 材料与方法
1.1 材料与试剂
人类非小细胞肺癌A549细胞,购于ATCC,由北京市食品添加剂工程技术研究中心实验室保存。
藻蓝蛋白标准品 美国Envirologix公司;DMEM细胞培养基 美国Gibco公司;胎牛血清 天津康源生物技术有限公司;实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qPCR)试剂盒 北京天根生物技术有限公司;TRIzol试剂北京鼎国昌盛生物技术有限公司。
1.2 仪器与设备
细胞培养箱 德国Heraeus公司;漩涡振荡仪德国IKA公司;超净工作台 上海博迅医疗生物仪器股份有限公司;数字型电子天平 德国Sartorius公司;CFX96Touch qPCR仪 美国Bio-Rad公司;Bioanalyzer2100核酸检测仪 美国安捷伦公司;miRNA-seq高通量测序仪 美国Illumina公司。
1.3 方法
1.3.1 细胞培养
A549细胞培养于含有体积分数10%胎牛血清、0.1 mg/mL链霉素以及100 units/mL青霉素的DMEM培养基中,在37 ℃、5% CO2条件下培养。细胞每3~4 d传代一次,取对数生长期的细胞进行实验。
1.3.2 藻蓝蛋白处理A549细胞
提前一天将细胞进行传代处理,细胞密度大约50%。第2天细胞完全贴壁后采用藻蓝蛋白处理A549细胞,做3 个平行藻蓝蛋白处理组与对照组(磷酸盐缓冲液处理细胞)。处理后48 h后收集细胞进行后续实验。研究结果中1~3号样品为对照组;4~6号样品为藻蓝蛋白处理组。
1.3.3 RNA文库构建
采用TRIzol法提取细胞总RNA,使用NanoDrop 2000测定RNA浓度,随后对RNA的完整性进行分析。采用小RNA建库试剂盒进行miRNA文库构建。
1.3.4 miRNA测序与序列分析
采用Illumina Hiseq 2000进行miRNA的测序。对所获得的原始序列进行过滤,去除低质量的序列;随后去除5’端和3’端接头序列、污染序列,以及小于18 nt和大于30 nt的序列,得到纯净序列。统计长度在16~27 bp的RNA序列长度的分布情况,然后选取GenBank数据库和Rfam数据库分别注释序列,尽可能发现并去除样本中的rRNA、snRNA、scRNA等。最后通过基因组比对除去重复序列和mRNA的降解片段。
1.3.5 miRNA的鉴定与差异分析
通过比对miRbase中已经收录的miRNA,鉴定到的为已知miRNA;结合参考序列进行发卡结构的预测,鉴定到的为新的miRNA。结合miRNA在各样本中的表达情况,得到全部miRNA的表达谱。将表达量变化2 倍以上且P<0.05的miRNA定义为差异表达的miRNA。
1.3.6 miRNA的靶基因预测和功能富集
使用Pathmatch软件进行靶基因的预测,然后对靶基因进行人类基因组和NCBI数据库注释,以及基因本体(gene ontology,GO)和京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)分析。
1.3.7 qPCR验证miRNA的表达
对每一个检测样品需要将待检测的miRNA与其内参U6分别进行反转录反应。根据RNA浓度计算2 μg所需要的体积,与DEPC混合至9 μL,每管中再分别加入2 μL的RT引物工作液(500 nmol/L),其中逆转录miRNA组中加入miRNA逆转录引物,逆转录U6组中加入U6逆转录引物。70 ℃、10 min后立即置于冰上。随后将第一步反应产物进行逆转录反应:42 ℃ 60 min,70 ℃ 10 min,预冷至4 ℃。逆转录结束后即可进行miRNA的扩增反应,检测miRNA时加入miRNA正向通用引物与Bulge-Loop miR反向通用引物,检测U6时加入U6正向引物与反向引物。
1.4 数据统计分析
实验数据均以平均值±标准差表示,采用SPSS 11.0软件进行单因素方差分析,其中P<0.05为显著性差异,P<0.01为极显著差异。
2 结果与分析
2.1 藻蓝蛋白对A549细胞体外增殖活性的影响
首先利用不同浓度的藻蓝蛋白(0、0.6、1.2、2.4、4.8 μmol/L)处理体外培养的A549细胞,检测细胞的存活率。由图1A可知,随着藻蓝蛋白浓度的增大,细胞的存活率显著降低,呈现明显的剂量依赖效应。因此,在后续的实验中,采用效果极显著的藻蓝蛋白浓度(4.8 μmol/L)进行细胞处理实验。由图1B可以发现,采用4.8 μmol/L的藻蓝蛋白处理细胞,能够显著抑制A549细胞的体外增殖能力。以上结果表明藻蓝蛋白对A549细胞的生长具有明显抑制效果,随后对其具体的调控机制进行了深入的测序分析。

图1 藻蓝蛋白对A549细胞体外增殖活性的影响Fig. 1 Effect of phycocyanin on in vitro proliferation of A549 cells
2.2 RNA样品的质量检测
首先对藻蓝蛋白处理后的细胞进行了总RNA的提取,并进行转录组测序前的质量检测。图2分别显示了对照组(图2A)和藻蓝蛋白处理组(图2B)的RNA完整性分析结果。结果显示,RNA的18S和28S亚基均出现了完整的峰,且集中程度较高,初步提示了RNA结构的完整性。此外,表1显示了RNA质量检测的基本参数,结果表明,6 个样品的RNA总量均超过了20 μg,OD260nm/280nm均大于1.8,RNA完整性参数(RNA integrity number,RIN)值均大于9,表明提取的RNA样本结构完整,没有发生明显降解,满足miRNA转录组测序的后续实验。
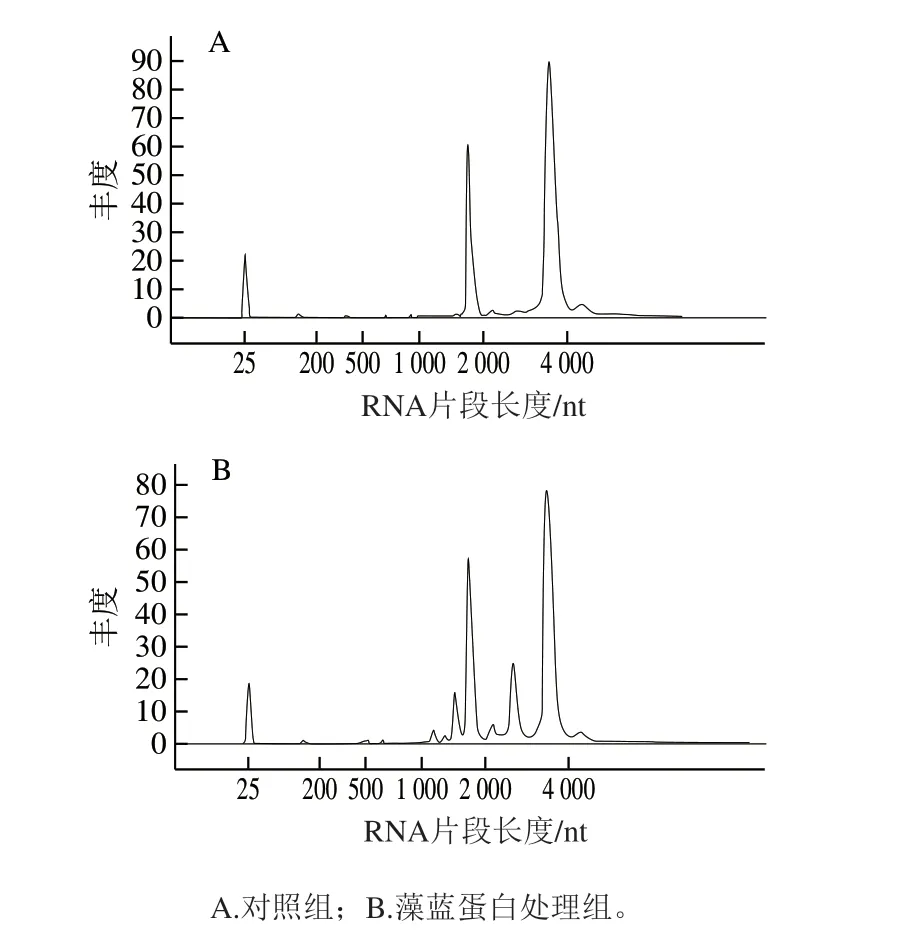
图2 RNA完整性检测结果Fig. 2 Analysis of the integrity of RNA
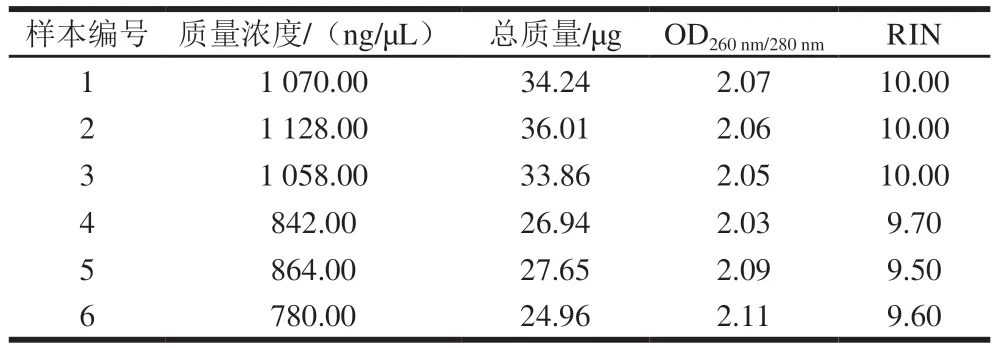
表1 不同样本RNA的质量检测数据Table 1 Quality evaluation of different RNA samples
2.3 sRNA的测序分析结果
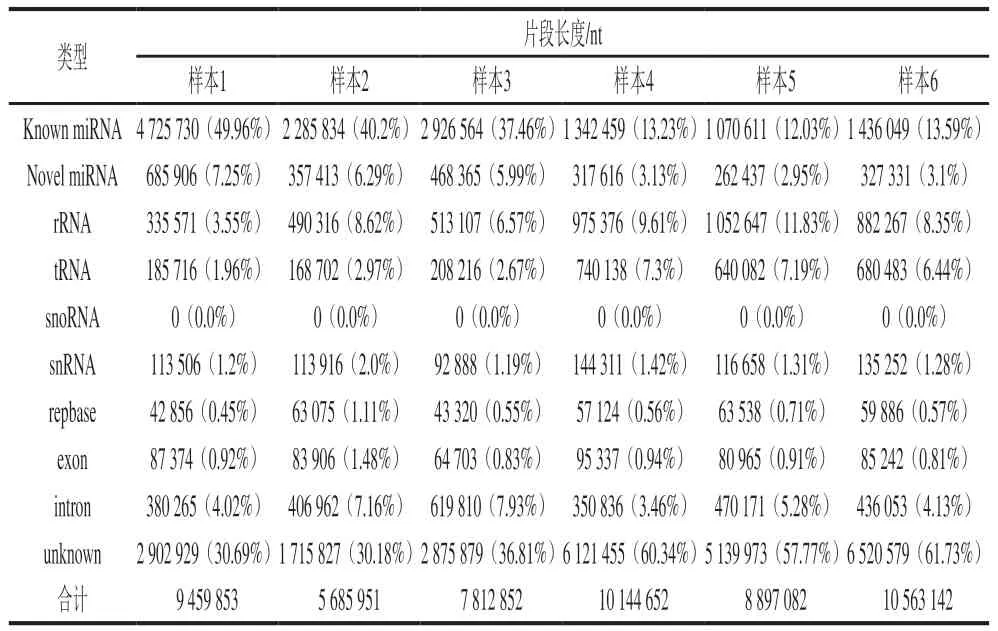
表2 不同样本中sRNA的统计结果Table 2 Statistical data of sRNA in different samples
miRNA-seq的测序结果不仅能提供miRNA的读长,还能提供其他类型RNA的相关信息。表2显示了包括miRNA在内的不同类型RNA的测序读长。结果显示,除了已知miRNA(known miRNA)和未知的新预测miRNA(novel miRNA)以外,核糖体RNA(rRNA)、转运RNA(tRNA)、核仁小分子(snoRNA)、小核RNA(snRNA)、重复序列(repbase)、外显子(exon)、内含子(intron)以及未知的sRNA序列(unknown)都被测序出来。在对照组中,已知m i R N A 占所有sRNA测序读长的(42.53±6.56)%,新预测的miRNA则占(6.51±0.66)%;在藻蓝蛋白处理组中,已知miRNA和新预测miRNA则分别占所有sRNA测序读长的(12.59±0.82)%和(3.06±0.09)%。虽然在测序读长信息上,对照组和藻蓝蛋白处理组存在较大的差异,但二者在miRNA成熟体上均测序出了相近的数量(表3)。结果显示,对照组和藻蓝蛋白处理后,miRNA成熟体(包含已知miRNA和新预测的miRNA)数量均达到了1 000 条以上。以上研究结果表明,miRNA-seq成功地筛选出了一批潜在的miRNA,构建了miRNA分析文库,是进一步寻找藻蓝蛋白调控差异miRNA的基础。
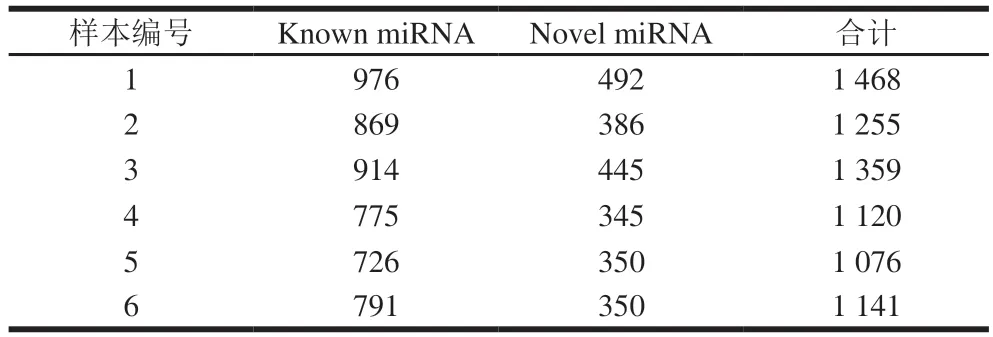
表3 不同样本中miRNA成熟体数量统计Table 3 Statistics of mature miRNAs in different samples
2.4 miRNA在不同样本中的表达量分析结果
不同类型的miRNA在细胞中发挥着不同的生理功能,所占的比例也大不相同[29]。本研究分析了对照组和藻蓝蛋白处理组中的miRNA丰度,筛选出了各样本中表达量前10 位的miRNAs。图3结果显示,高丰度的新预测miRNA在不同样本中的表达模式基本一致,其中丰度前5 位分别为:18_34113、20_36370、14_27440、22_37559以及16_30535,表明这些新预测的miRNA在藻蓝蛋白处理前后较稳定,可能不是藻蓝蛋白主要的调控因子;相反,一些其他新预测的miRNA(例如17_31507)在藻蓝蛋白处理组均有一定量的表达,但并没有出现在对照组中,提示这一类新预测的miRNA可能受到了藻蓝蛋白的影响,出现表达量的差异性变化。新的miRNA仅仅是测序信息的预测结果,具体是否可靠还需要进一步的验证。此外,对于已知的miRNA,图3结果显示,6 个研究样本中,高丰度的已知miRNA(例如miR-21-5p、miR-27b-3p等)均出现较一致的表达模式。并且有研究表明,miR-21-5p能够通过靶向调控TGFBI和SMAD7抑制肺癌细胞的生长[30-31],miR-27b-3p也被报道在肺癌的发生过程中起到调控作用[32]。以上结果表明,藻蓝蛋白在A549细胞中的调控很可能是作用于一些低丰度的特异性miRNA,因此,对不同处理的样本进行了差异miRNA的筛选分析,进一步筛查藻蓝蛋白可能调控的miRNA。
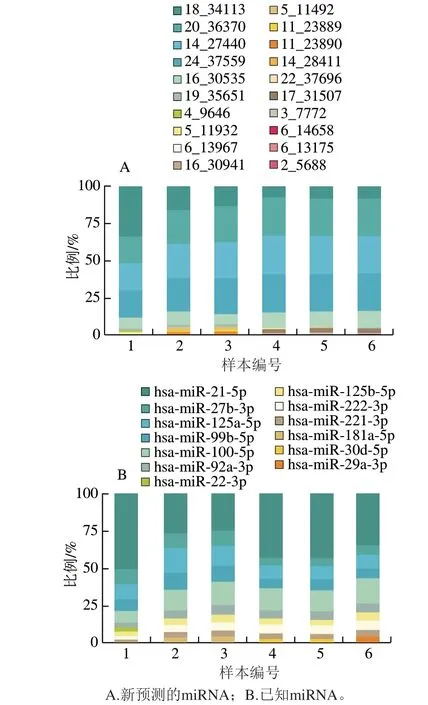
图3 各样本中表达量前10 位的miRNAsFig. 3 Top 10 most expressed miRNAs in each sample
2.5 差异miRNA的筛选分析结果

图4 藻蓝蛋白处理A549细胞后差异miRNA的火山分布图Fig. 4 Volcano map of differentially expressed miRNAs in A549 cells after undergoing phycocyanin treatment
采用miRNA-seq高通量测序,对藻蓝蛋白处理后的差异miRNA进行了筛选分析。图4结果表明,藻蓝蛋白处理后,均有miRNA出现了上调或下调的表达。通过统计分析,共有136 个差异miRNA被筛选出来,其中藻蓝蛋白处理后显著上调的miRNA有74 个,显著下调的miRNA有62 个。表4显示了样本间差异程度最高的19 个已知miRNA。由分析结果可以看出,miR-4521、miR-27b-5p、miR-7974、miR-574-5p等在藻蓝蛋白处理A549细胞后表达量出现显著下调;而miR-4485-3p、miR-146a-5p、miR-27a-5p、miR-93-5p等的表达量则在藻蓝蛋白处理后出现了明显上调。有研究表明,miR-4521对人类慢性淋巴细胞白血病发生和发展有调控作用[33],但对肺癌细胞的生长还没有明确的报道;而miR-27a-5p、miR-93-5p等则被发现对肺癌细胞有一定的抑制作用[34-35]。Zhou Rui等报道miR-574-5p能够通过PTPRU促进肺癌细胞的转移[36],而本研究发现藻蓝蛋白处理肺癌细胞后miR-573-5p出现了明显的下调表达,这与已知的文献报道一致。

表4 不同样本间差异程度最高的19 个已知miRNATable 4 Top 19 most differentially expressed known miRNAs in different samples
随后对这些差异miRNA进行了子聚类分析,图5显示,根据各样本中差异miRNA表达模式的不同,这些miRNA可以聚类为4 类:与对照组相比,显著下调表达的miRNA(A类)、极显著下调表达的miRNA(B类)、显著上调表达的miRNA(C类)以及极显著上调表达的miRNA(D类)。本研究通过miRNA-seq筛选出了一批藻蓝蛋白处理后差异性表达的miRNA,这些差异miRNA的发现为研究藻蓝蛋白在非小细胞肺癌中的调控机制提供了重要的理论参考。
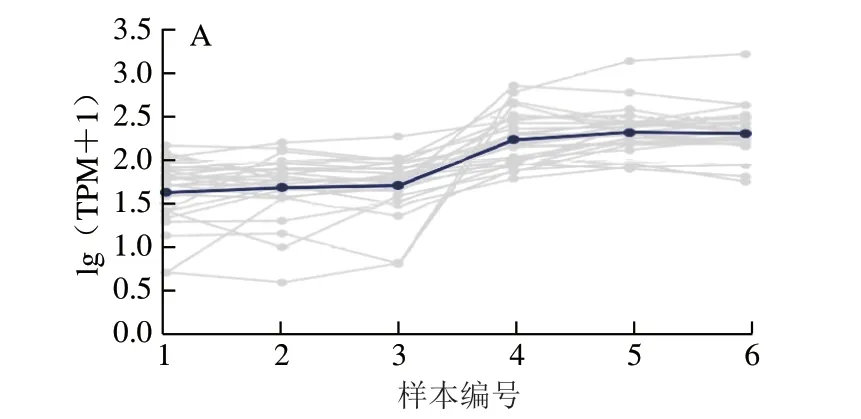
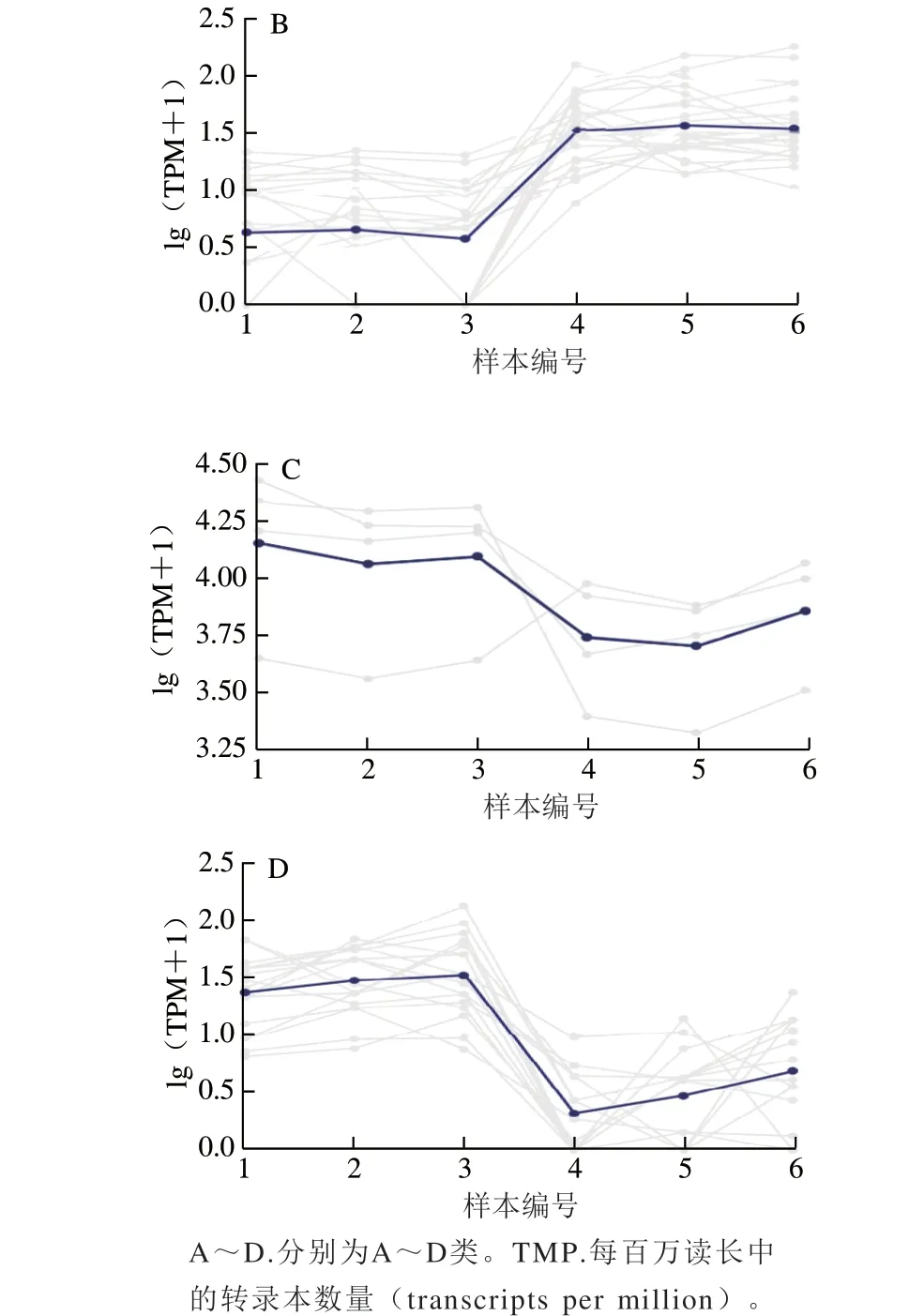
图5 差异miRNA的不同表达模式分析Fig. 5 Expression pattern analysis of differentially expressed miRNAs in different samples
2.6 差异miRNA的验证
对筛选得到的几个代表性的差异miRNA进行了qPCR的生物学验证。图6结果显示,藻蓝蛋白处理A549细胞后,与对照组相比,miR-4521、miR-27b、miR-7974、miR-574的表达量均出现显著下调;miR-4485、miR-146、miR-27a以及miR-93的表达量出现显著上调,此结果与miRNA-seq的测序结果是一致的,进一步证实了这些差异miRNA确实受到了藻蓝蛋白的影响,可能在藻蓝蛋白抑制非小细胞肺癌过程中起到调控作用。
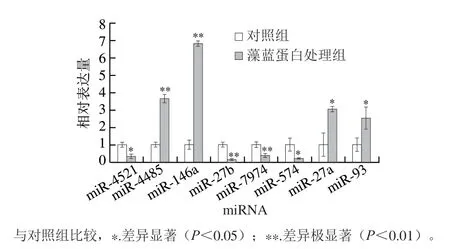
图6 部分差异miRNA的qPCR验证Fig. 6 qPCR analysis of some differentially expressed miRNAs
2.7 miRNA的靶基因功能分析结果
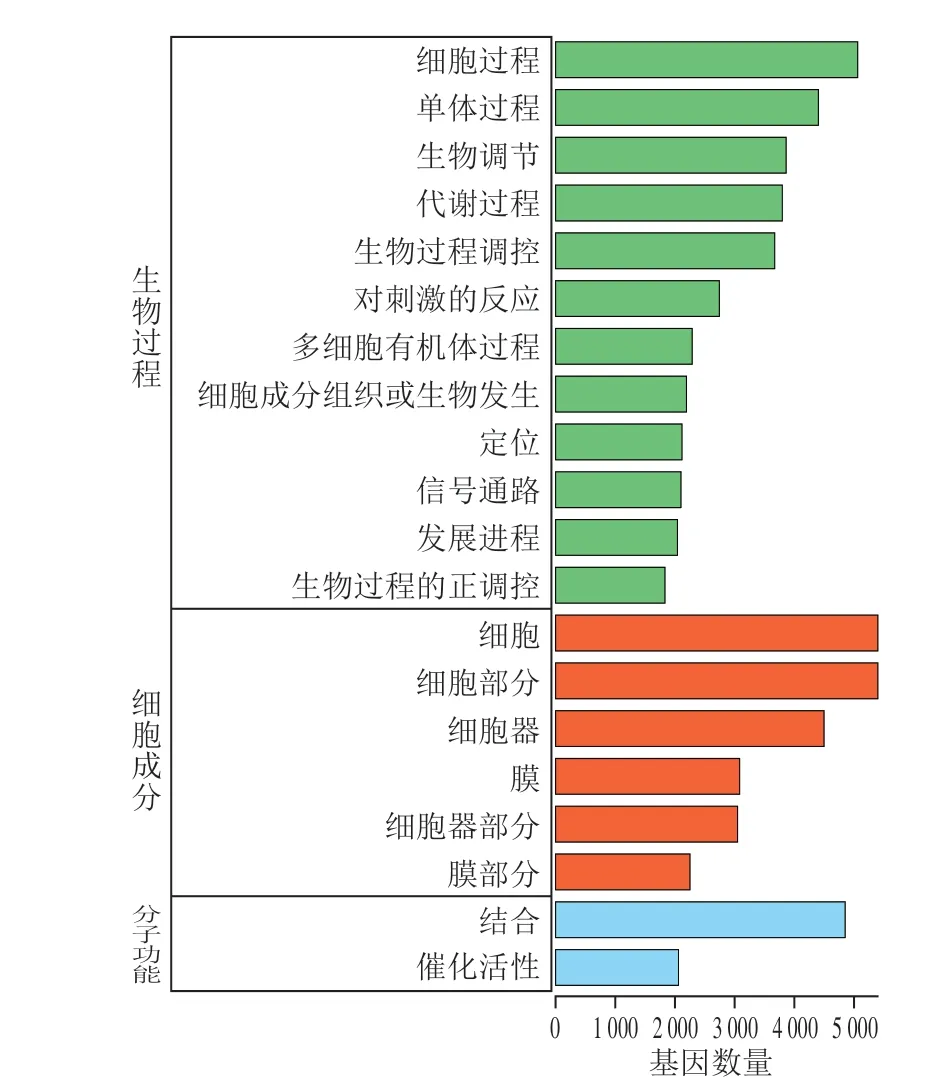
图7 miRNA靶基因的GO功能注释分析Fig. 7 GO annotations analysis of target genes for miRNAs
使用miRanda软件对筛选出的miRNA进行靶基因的预测分析,共预测得到了5 186 个潜在的靶基因。首先对这些靶基因进行了GO注释分析,结果如图7所示。GO注释可将基因分为三大类:生物过程、细胞组分和分子功能。在生物过程中,靶基因功能主要集中在代谢调控、应激反应和信号转导等;在细胞组分和分子功能中,主要集中在分子相互作用以及酶催化活性等。GO功能注释提供了靶基因的功能分类,但缺乏具体的功能信息,因此,为全面分析受到藻蓝蛋白调控miRNA靶基因的功能,又对这些基因进行了GO富集分析(图8)。结果显示,细胞活动相关的许多生物学过程都在GO富集分析中出现显著性差异,主要包括细胞离子转运过程、细胞膜结构的调控过程、细胞增殖信号转导调控过程、细胞骨架系统形态发生过程等。藻蓝蛋白对A549细胞具有明显的增殖抑制作用,而GO富集分析中细胞增殖信号转导过程与本研究相一致,进一步表明了测序结果的准确性。
在预测的靶基因中,p21激活磷酸激酶4(p21 activated kinase 4,PAK4)在细胞生长与增殖过程中参与重要的调控作用,是miR-6721-5p的预测靶点;分裂原激活的蛋白激酶13(mitogen-activated protein kinase 13,MAPK 13)参与了细胞有丝分裂过程信号调控的级联放大反应,是miR-5008-5p的预测靶点;Ras作为一种重要的GTP结合蛋白,对细胞的生长、分化、细胞骨架和蛋白运输都有重要的影响,研究筛选得到的Ras结合结构域家族10蛋白(Ras association domain family member 10,RASSF10)被预测为miR-4769-3p的靶基因,也是藻蓝蛋白抑制A549细胞生长可能的一条重要途径。
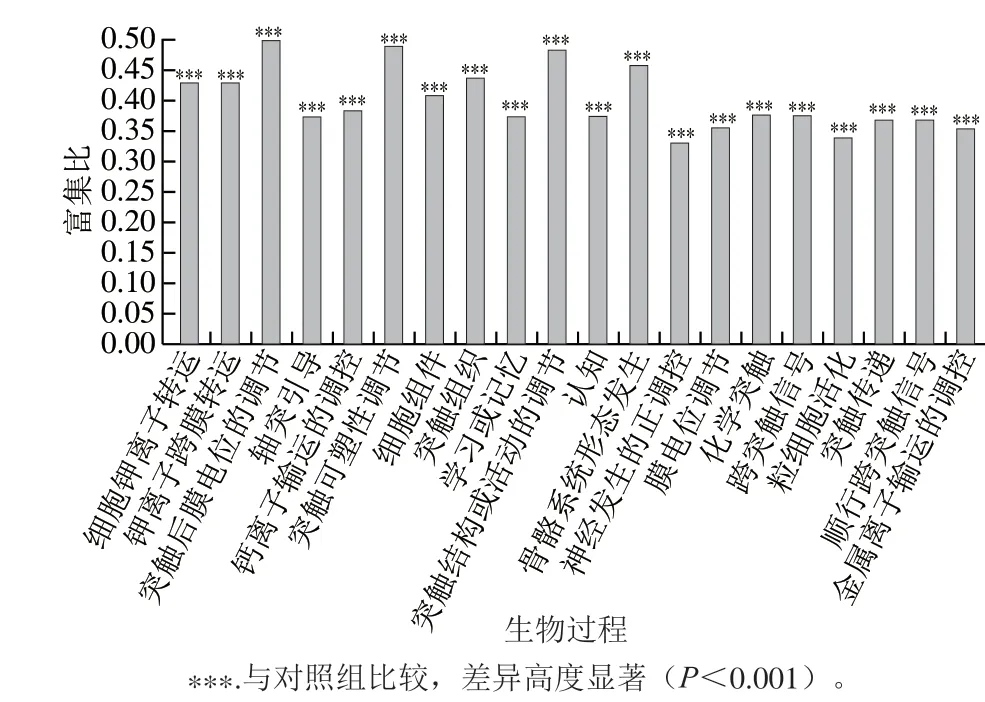
图8 miRNA靶基因的GO富集分析Fig. 8 GO enrichment analysis of target genes for miRNAs
3 结 论
本研究以高通量miRNA转录组技术为研究手段,对藻蓝蛋白处理后的非小细胞肺癌A549细胞进行了测序分析。结果表明,已知和新预测的大量miRNA都被测序出,并且在不同的处理样本中具有不同的表达模式;通过统计学分析筛选得到了136 个表达差异的miRNA,其中藻蓝蛋白处理后显著上调的miRNA有74 个,显著下调的miRNA有62 个。根据这些差异miRNA在不同样本中的表达情况,可以聚类为4 类不同的表达模式;qPCR对这些miRNA进行表达量的验证,与转录组测序结果相吻合;此外,miRNA潜在靶基因的GO注释和功能富集分析结果显示,与细胞增殖信号转导调控过程相关的靶基因例如Pak4、Mapk13、Rassf10等均被预测为差异miRNA的全新靶点,可能在藻蓝蛋白抑制非小细胞肺癌增殖过程中起到重要的调控作用。不同于传统的mRNA测序,本研究采用小分子miRNA测序技术探究了藻蓝蛋白抑制肺癌细胞生长的调控机制,为抗肿瘤类功能食品因子的利用提供了理论依据,同时为非小细胞肺癌的靶向治疗提供了重要的理论参考。
