湖北木林子保护区山矾属植物空间分布格局及其关联性
2020-11-24艾训儒吴漫玲刘松柏
王 进,朱 江,2,*,艾训儒,2,姚 兰,2,黄 小,吴漫玲,朱 强,刘松柏
1 湖北民族大学林学园艺学院, 恩施 445000 2 生物资源保护与利用湖北省重点实验室, 恩施 445000 3 湖北木林子国家级自然保护区管理局, 恩施 445000
种群是指一定时期内分布在一定范围的同种生物个体的集合[1]。种群空间格局是种群个体在二维空间的分布状况,是生态学研究的热点之一,有助于了解种群生物学特性和潜在生态学过程[2-3]。种群空间分布格局及其关联性对物种的生长、繁殖、死亡和资源利用具有显著影响[3- 5]。种群空间分布格局不仅因种而异,还表现出同一物种在不同生长阶段存在明显差异[6]。Plotkin等[7]发现种群各生长阶段的聚集程度随空间尺度变化而变化;Comita等[8]和Lai等[9]发现同一物种在不同生长阶段对环境的响应不同。因此,研究种群空间分布格局及关联性有助于探索在群落构建中物种共存机制的作用和潜在的生态学过程[10]。
目前,种群空间格局研究对象主要集中在优势种或主要种[11-12]、珍稀濒危植物[13-14]等类群,而对于同属物种的空间分布格局研究比较少见。同属物种的研究有利于减少系统发生学差异引起的混淆效应,分析同属物种的空间格局及关联性,为揭示同属物种共存机制提供了可能[15-16]。通常同属物种具有共同进化起源,在生态学和形态学特征上具有许多相似之处,可能会以相似的生活史策略获取资源[17],使同属物种对生境的趋同适应性更高。一般在生态学研究中认为同属物种可能存在强烈的种间竞争,从而限制其共存[18-19]。然而,以往的研究也有很多不一致的结论,其中两个方面常常被忽略:第一,样本量不足[20];第二,同属物种是不同区域或不同生境的[21],这就不能合理解释同属物种的共存。因此,基于大型固定监测样地且生活在相同或相似生境范围内的同属物种空间格局及关联性的研究是非常必要的。
湖北木林子国家级自然保护区是常绿落叶阔叶混交林保存最为完好的区域之一[22]。其中,山矾属作为木林子大样地的主要优势属之一,包含山矾(Symplocossumuntia)、光叶山矾(Symplocoslancifolia)、薄叶山矾(Symplocosanomala)、白檀(Symplocospaniculata)和老鼠矢(Symplocosstellaris)5个树种,山矾和光叶山矾是木林子大样地的主要优势种[23-24]。本研究基于木林子保护区的山矾和光叶山矾进行同属物种空间格局及种间关联分析,试图回答以下几个问题:(1)山矾和光叶山矾在种群结构和空间分布格局上是否存在差异?(2)它们的空间分布格局和空间关联性是如何随空间尺度和物种各生长阶段变化的?(3)同属物种之间是否存在强烈竞争?
1 研究区概况
湖北木林子国家级自然保护区位于恩施土家族苗族自治州鹤峰县境内,地理坐标109°59′30″—110°17′58″E,29°55′59″—30°10′47″N,海拔1100.0—2095.6 m,属武陵山脉北支余脉,地势由西北和东南向中间逐渐倾斜。该区属于亚热带季风性湿润气候,雨热同期,时空分布不均。土壤为黄棕壤、棕壤和黄壤。主要植被类型为亚热带常绿落叶阔叶混交林[25]。
木林子大样地位于湖北木林子国家级自然保护区核心区内,距最高峰——牛池峰(海拔2095.6 m)约500 m。样地内蒸发小,湿度大,年降水量1733 mm,年平均相对湿度为82%,无霜期270—279 d[22],年平均气温为15.5℃,全年有效积温(≥10℃)约4925.4℃。海拔介于1588.5—1780.2 m,坡度介于13.6°—49.6°,坡向介于94.8—226.8,土壤为黄棕壤,pH值为4.5—6.5。
2 研究方法
2.1 样地设置及调查
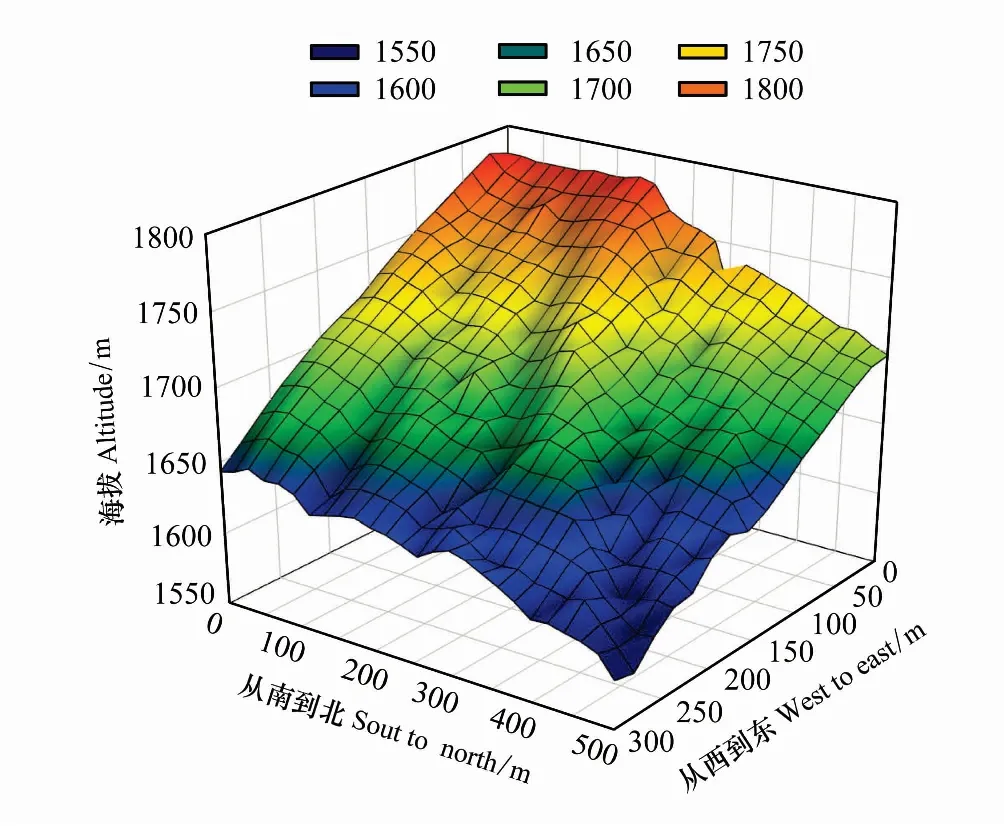
图1 木林子大样地地形图Fig.1 Topographic map of Mulinzi large sample plot
2013年在木林子国家级自然保护区选择地势相对平缓、内部地形相对一致的区域,按照热带林业科学中心(CTFS)样地建设标准和监测技术规范[26],建立东西长300 m、南北长500 m的15 hm2固定监测样地(图1)。对样地内胸径(DBH)≥1.0 cm的山矾和光叶山矾进行每木检尺(DBH<1.0 cm的山矾和光叶山矾幼苗直接记数统计),在1.3 m处油漆标记、挂牌,测定DBH、树高,记录植株萌生状况,观测植物个体的相对坐标值和海拔、坡度、坡向等生境因子。
2.2 优势种径级划分
参照相关研究的径级划分[27],结合山矾和光叶山矾的胸径特征,划分4个径级:1 cm≤径级Ⅰ<2.5 cm、2.5 cm≤径级Ⅱ<5 cm、5 cm≤径级Ⅲ<10 cm、径级Ⅳ≥10 cm。为分析山矾和光叶山矾各生长阶段的空间分布格局及关联性,分为幼树(径级Ⅰ)、小树(径级Ⅱ)、中树(径级Ⅲ)和成年树(径级Ⅳ)4个生长阶段。
2.3 点格局分析方法
利用成对相关函数g(r)进行点格局分析。g(r)函数由RipleyK函数K(r)=πr2推演而来[28]。g(r)函数能较为敏感地判断某一尺度上点的实际分布偏差期望值的程度,排除计算过程产生的累积效应,是评估聚集程度的重要分析方法。利用单变量g(r)函数分析物种的空间聚集程度,函数为:
式中,r为空间尺度距离。
采用双变量g12(r)函数分析2个不同群体的空间关联性,函数为:
选择具有明确生态学意义、能准确描述数据偏离理论程度的零假设模型(null model)对空间点格局分析非常关键[29]。本研究选用以下零模型[29-30]:(1)完全空间随机模型(complete spatial randomness, CSR):假设物种空间分布不受任何生物或非生物过程影响,在研究区域内各点出现的概率相同。(2)异质泊松模型(heterogeneous Poisson, HP):依据密度函数λ(s)(s表示样地内任意一点的位置,用坐标(x,y)表示)决定个体的分布,运用高斯核函数[31]进行密度估计,选取sigma=25 m消除大尺度上的生境异质性的影响。(3)前提条件模型(antecedent condition, AC):让高龄级的位置固定不变,低龄级的位置随机变化。单变量g(r)函数采用完全空间随机模型和异质泊松模型;双变量g12(r)函数采用完全空间随机模型和前提条件模型。通过99次Monte Carlo随机模拟,分别利用模拟的最大值和最小值生成上下两条包迹线,计算99%的置信区间。当g(r)值和g12(r)值在包迹线上方时,呈聚集分布或正关联;在包迹线之间时,呈随机分布或无关联;在包迹线下方时,呈均匀分布或负关联。采用R 3.4.1中“spatstat”程序包进行数据分析;采用Origin 2017、SigmaPlot 12.5等软件进行绘图。
3 结果与分析
3.1 山矾与光叶山矾的径级结构
在15 hm2样地中,山矾有3415株,光叶山矾有2724株,分别占样地树木总株数的4.1%和3.2%。尽管山矾和光叶山矾在个体数上存在较大差异,但都呈典型金字塔形结构(图2)。从生长阶段来看,个体数较多的山矾的中树、成年树个体数较少,而个体数较少的光叶山矾中树、成年树个体数却相对较多。综上,样地内山矾和光叶山矾属增长型种群,林下存在丰富的幼树和小树等幼龄个体,种群更新状况良好。
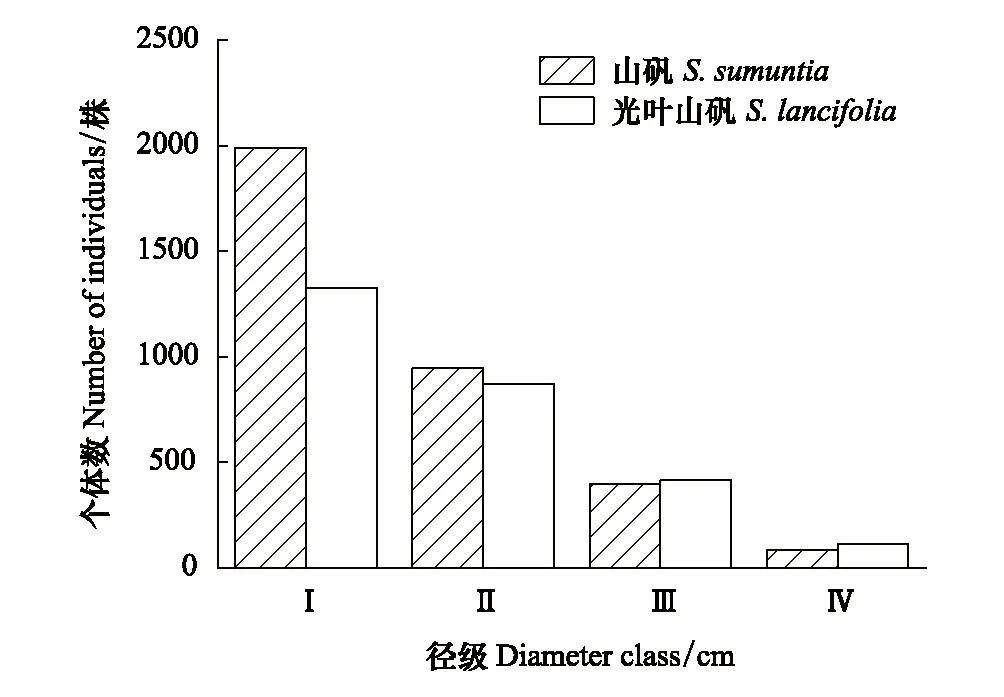
图2 木林子大样地山矾与光叶山矾的径级结构 Fig.2 Diameter structure of Symplocos sumuntia and Symplocos lancifolia in Mulinzi large sample plotⅠ—Ⅳ表示山矾和光叶山矾的径级结构划分为第1—4径级,Ⅰ—Ⅳindicates that the diameter structure of Symplocos sumuntia and Symplocos lancifolia as 1st to 4th grade
3.2 山矾与光叶山矾及各生长阶段的空间分布及空间点格局分析
山矾和光叶山矾各生长阶段的空间分布如图3所示。山矾和光叶山矾在样地内的空间分布具有较为明显的聚集斑块状分布特征。其中,山矾种群各生长阶段主要分布在样地海拔较低的东部和东北部;光叶山矾种群各生长阶段主要分布在样地海拔较高的西部和西南部,在样地北部也有部分聚集分布。
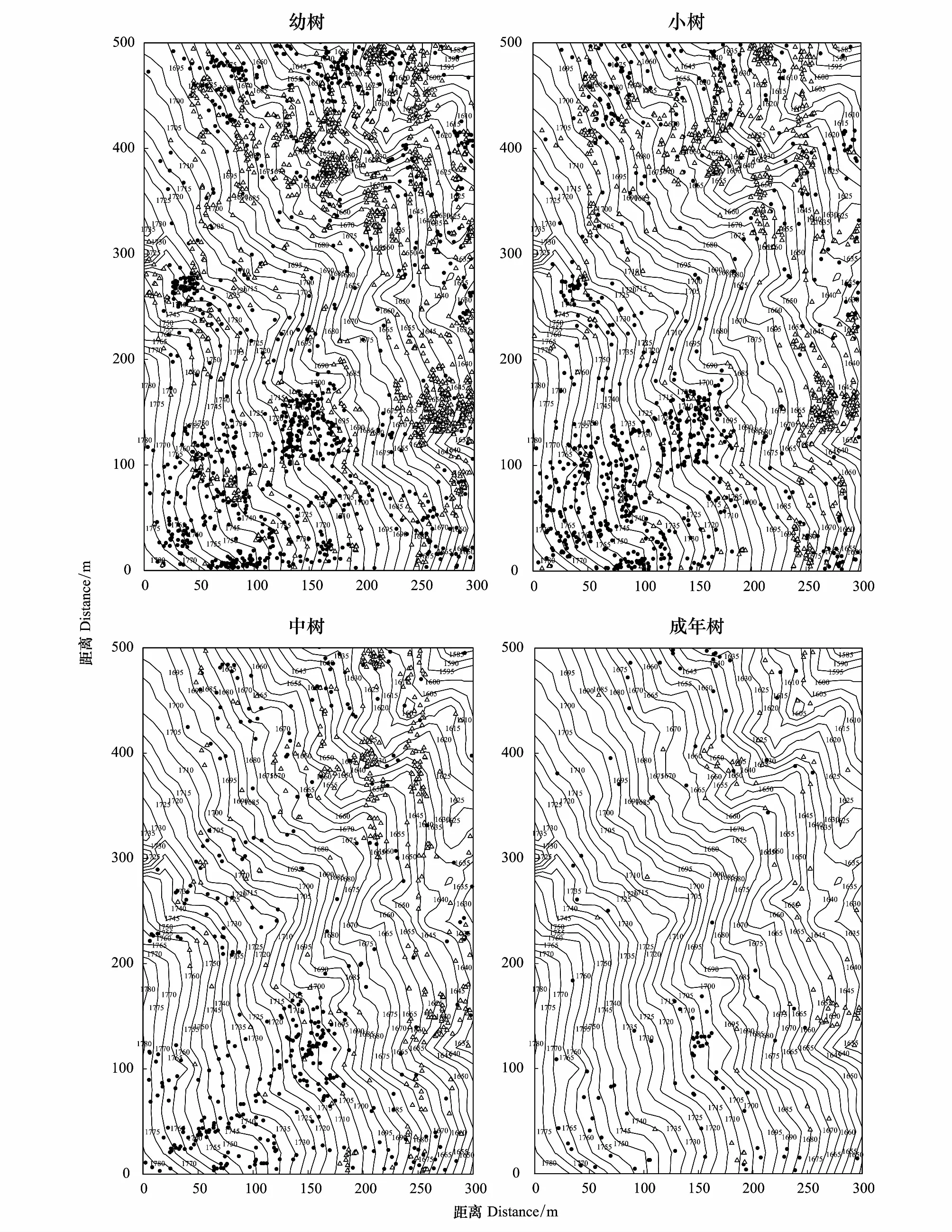
图3 山矾与光叶山矾各生长阶段的空间分布Fig.3 Spatial distribution of Symplocos sumuntia and Symplocos lancifolia in each growth stages△为山矾;·为光叶山矾
基于完全空间随机分布(CSR, complete spatial randomness)和异质泊松分布(HP, heterogeneous Poisson)模型分析表明,山矾及各生长阶段的空间聚集程度随着尺度增大而逐渐减小,分布特征由聚集分布向均匀分布过渡(图4、图5)。CSR模型分析显示山矾及各生长阶段的聚集程度分别是0—125、0—120、0—100、0—113、0—48 m;山矾成年树在r>49 m后呈随机分布。HP模型排除25 m以上的大尺度生境异质性的影响,结果显示山矾及各生长阶段的聚集分布尺度分别是0—36、0—36、0—18、0—18、0—11 m;山矾在47—75、94—108、129—150 m表现为均匀分布;其幼树、小树、中树和成年树也在不同尺度上表现出均匀分布特征。
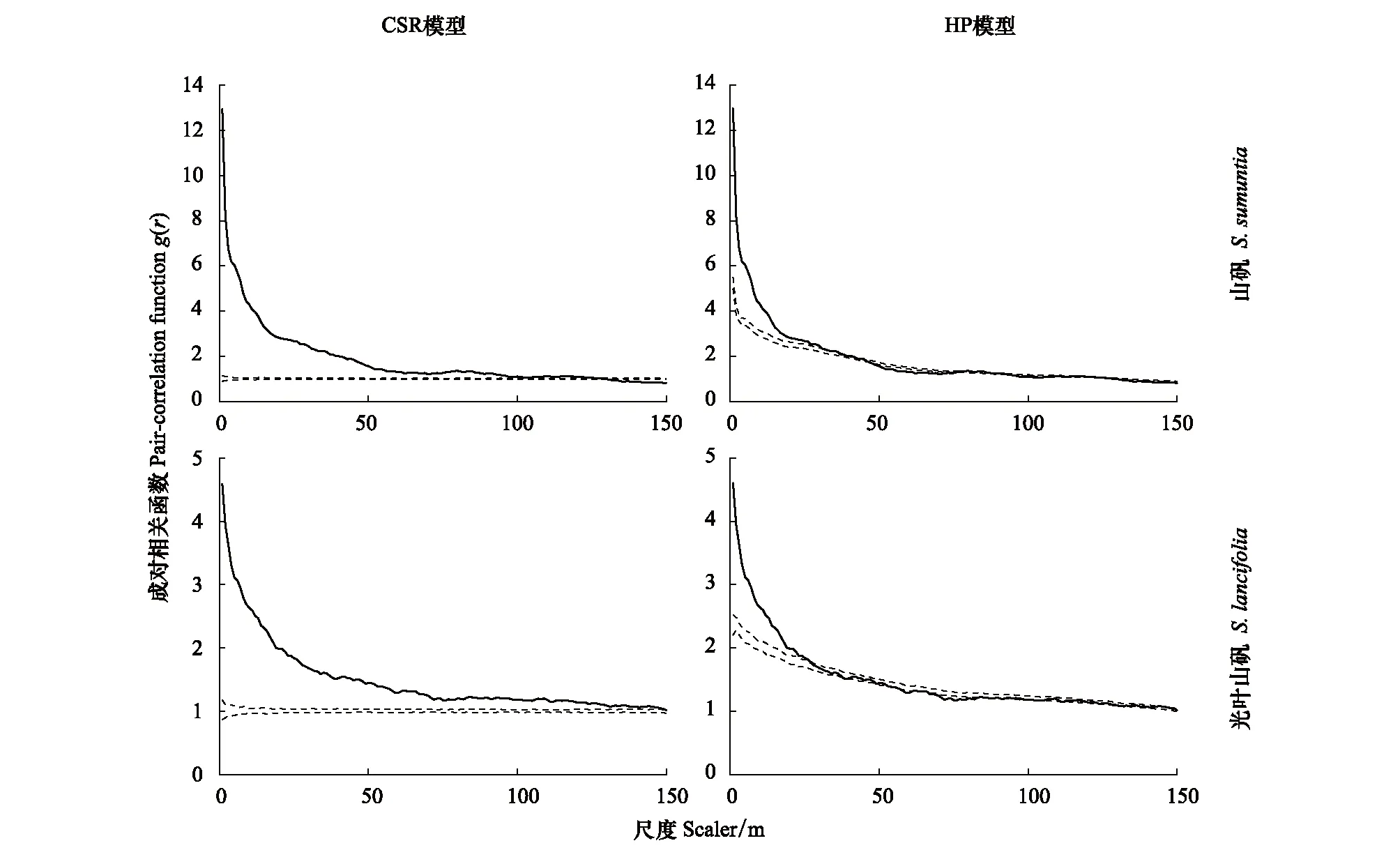
图4 山矾与光叶山矾空间分布格局Fig.4 The Spatial distribution patterns of Symplocos sumuntia and Symplocos lancifolia实线表示成对相关函数;虚线表示99%包迹线; 零模型为完全空间随机模型(CSR, complete spatial randomness)和异质泊松模型(HP, heterogeneous Poisson)
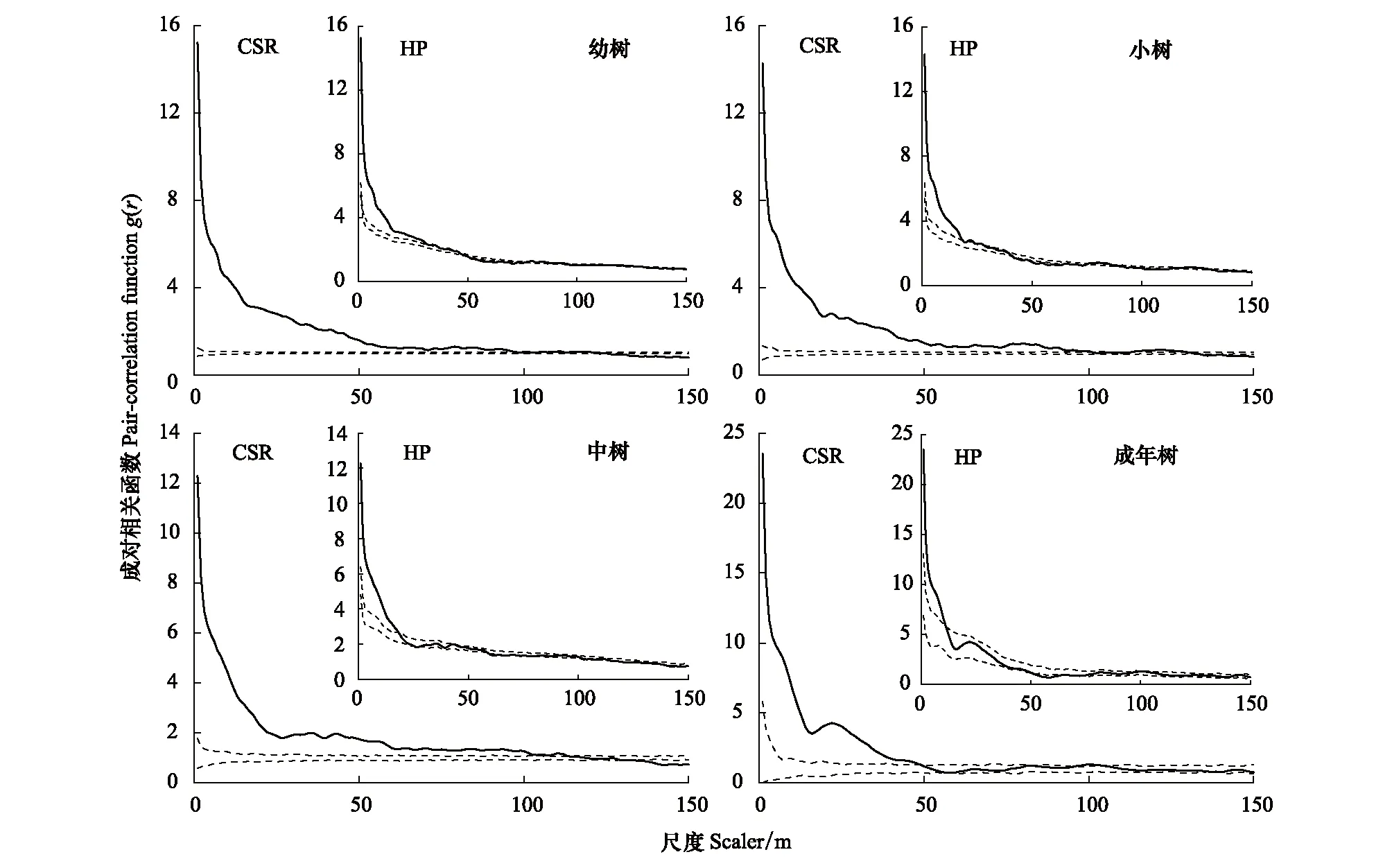
图5 山矾各生长阶段的空间分布格局Fig.5 Spatial distribution patterns of Symplocos sumuntia in each growth stages实线表示成对相关函数;虚线表示99%包迹线; 零模型为完全空间随机模型(CSR)和异质泊松模型(HP)
光叶山矾及各生长阶段的空间聚集程度随尺度增大而逐渐减小,分布特征由聚集分布向随机分布过渡(图4、图6)。CSR模型分析显示光叶山矾及各生长阶段在0—150 m尺度上总体表现为聚集分布,其中幼树在130—150 m、小树在143—150 m区域内由聚集分布过渡为随机分布,而成年树在65—73、93—110、140—148 m有微弱的随机分布特征。HP模型结果显示,光叶山矾及各生长阶段的聚集分布尺度分别是0—26、0—25、0—20、0—20、0—15 m;光叶山矾在67—83 m;幼树在31—42、57—61、68—81 m之间表现出均匀分布特征。
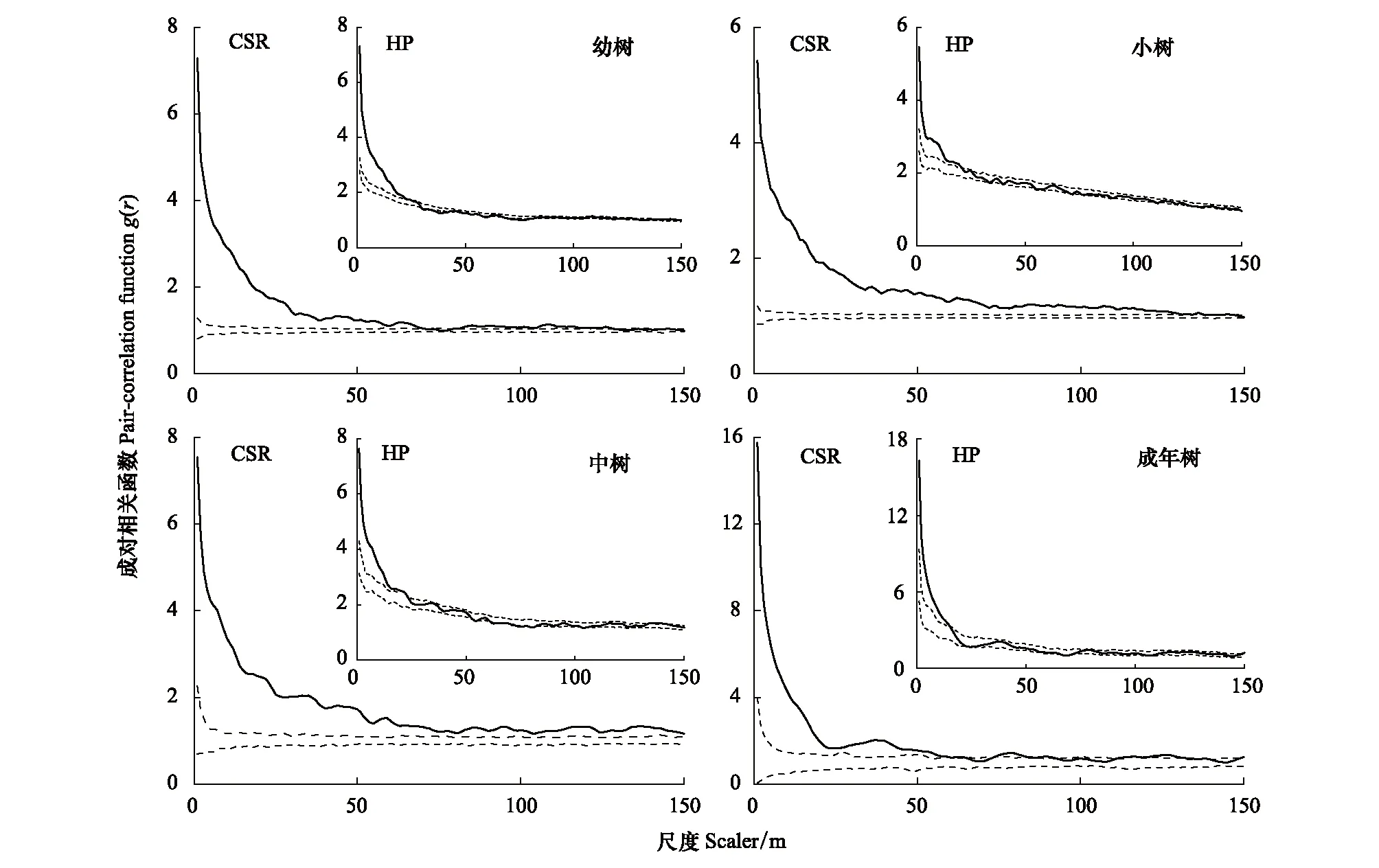
图6 光叶山矾各生长阶段的空间分布格局Fig.6 Spatial distribution patterns of Symplocos lancifolia in each growth stages实线表示成对相关函数;虚线表示99%包迹线; 零模型为完全空间随机模型(CSR)和异质泊松模型(HP)
3.3 山矾与光叶山矾各生长阶段的种内关联性
基于完全空间随机模型的双变量g12(r)函数分析结果表明(图7、图8):山矾幼树、小树、中树和成年树的空间关联性呈正关联,随尺度增大,逐渐由正关联向负关联过渡;而成年树与小树、中树在较大尺度表现出微弱的负关联。光叶山矾除小树与成年树在139—150 m尺度上表现出负关联外,其余生长阶段间都呈正关联,并随尺度增大,正关联的强度逐渐降低。
基于前提条件模型排除生境异质性影响的分析结果表明(图7、图8):山矾幼树与小树在0—18、115—132 m呈正关联,49—72 m呈负关联,其余尺度无关联;幼树与中树在0—16 m正关联,111—125 m微弱的正关联波动,59—73 m负关联,其余尺度无关联;幼树与成年树在0—29 m正关联,43—74 m负关联,其余尺度无关联;小树与中树、小树与成年树、中树与成年树分别在0—17、0—27、0—15 m呈正关联,其余尺度几乎无关联。虽然山矾各生长阶段间的空间关联性都呈正关联,但幼树与中树间正关联的程度最低只达到7.75。光叶山矾幼树与小树、幼树与中树、小树与中树、中树与成年树在0—23、0—25、0—18、0—19 m呈正关联,其余尺度上无关联;幼树与成年树在0—2、9—31、96—150 m呈正关联,其余尺度无关联;小树与成年树在0—17、110—146 m呈正关联,其余无关联。成年树与幼树、小树随着尺度增大从正关联逐渐过渡到无关联,而在较大尺度时则又表现出正关联特征。
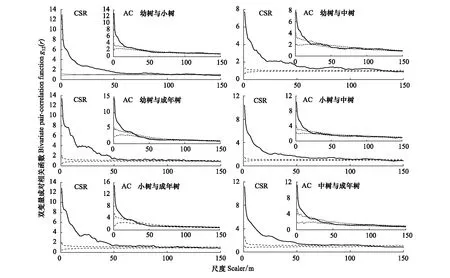
图7 山矾各生长阶段的种内关联性Fig.7 The intraspecific relevance in each growth stages of Symplocos sumuntia实线表示双变量成对相关函数;虚线表示99%包迹线; 零模型为完全空间随机模型(CSR)和前提条件模型(AC, antecedent condition)
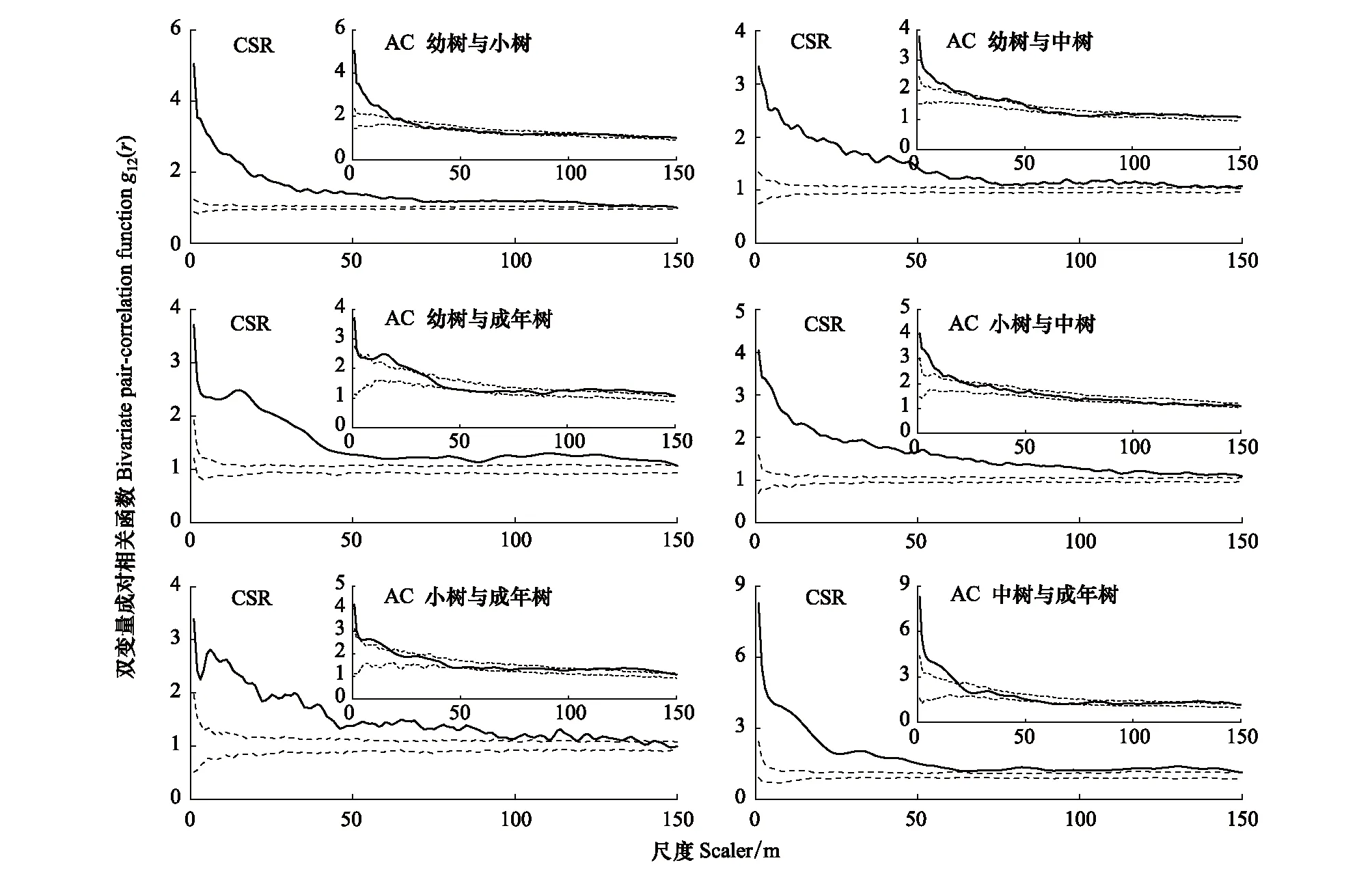
图8 光叶山矾各生长阶段的种内关联性Fig.8 The intraspecific relevance in each growth stages of Symplocos lancifolia实线表示双变量成对相关函数;虚线表示99%包迹线; 零模型为完全空间随机模型(CSR)和前提条件模型(AC)
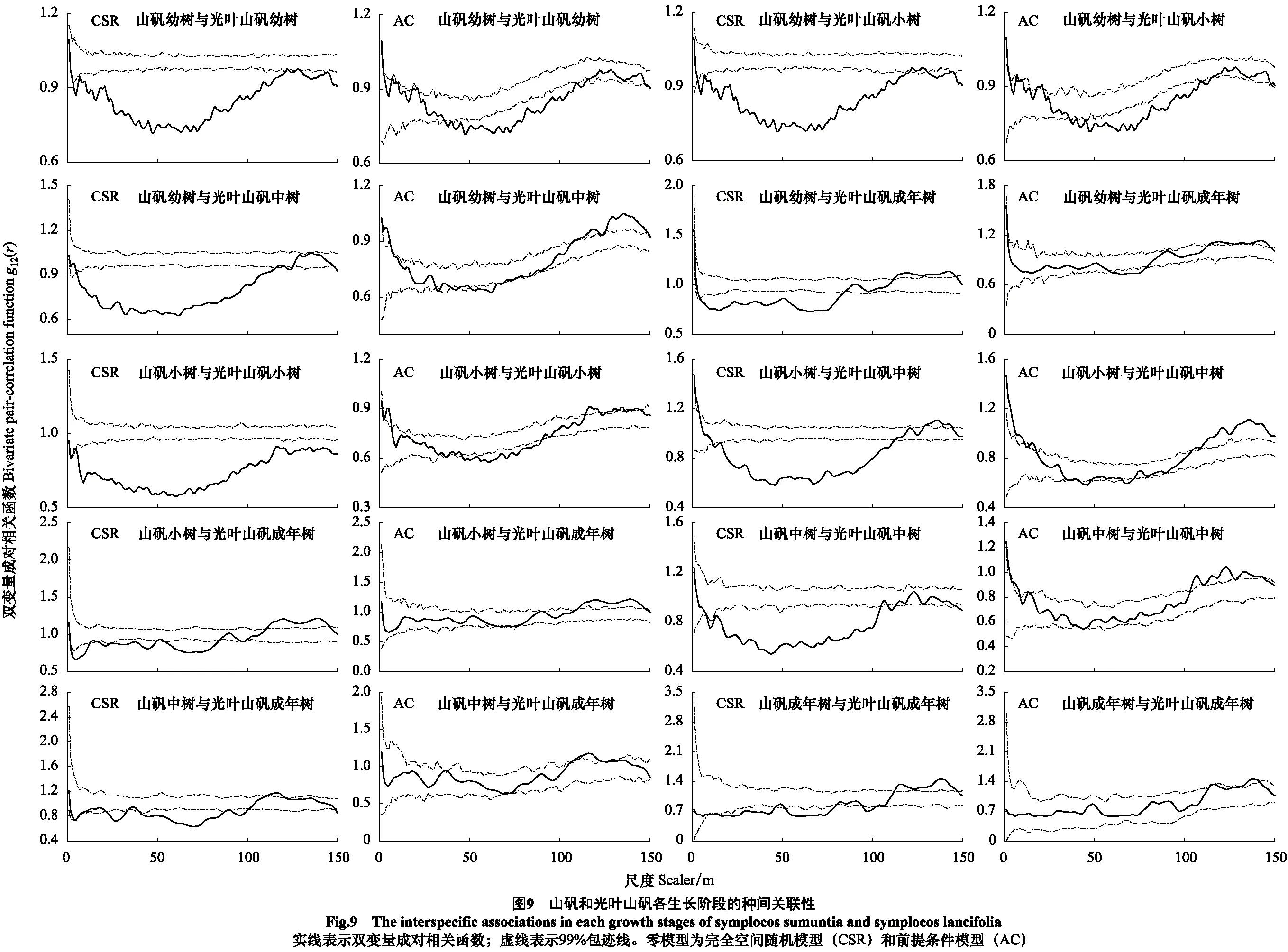
3.4 山矾与光叶山矾各生长阶段的种间关联性
基于完全空间随机模型的双变量g12(r)函数分析结果表明(图9):山矾幼树和光叶山矾成年树在5—84 m呈负关联,在112—146 m呈正关联,其余尺度上无关联性;山矾小树与光叶山矾成年树、山矾成年树与光叶山矾成年树在113—147、111—145 m表现出由负关联向正关联过渡的特征。山矾和光叶山矾相同生长阶段间除成年树随空间尺度增大表现出由负关联向正关联过渡特征外,主要表现为负关联。
基于前提条件模型排除生境异质性影响的分析结果表明(图9):山矾幼树与光叶山矾幼树、小树、成年树在37—115、37—114、58—78 m呈负关联,其余尺度无关联;山矾小树与光叶山矾小树在46—89 m为负关联,其余尺度几乎无关联;山矾幼树与光叶山矾中树在0—5、109—150 m,山矾小树与光叶山矾中树在0—17、104—150 m,山矾中树与光叶山矾中树在0—8、103—143 m呈正关联,其余尺度均为无关联,总体呈“U”型趋势;而山矾小树与成年树除111—146 m呈正关联外,其余尺度无关联;山矾中树、成年树与光叶山矾成年树间几乎无关联,但在较大尺度上表现出微弱的正关联特征,这可能是随机波动引起的。
4 讨论
通常来说,植物种群结构和空间分布格局是其自身生物学特性、种内种间相互关系及环境因素共同决定的,可以反映出种群动态变化规律和群落未来发展趋势[32]。研究表明,同属物种山矾与光叶山矾种群结构基本一致,呈典型金字塔形。湖北木林子国家级自然保护区是以保护典型中亚热带常绿落叶阔叶混交林为主,该区受人为干扰较少,山矾和光叶山矾种质资源充沛,样地内地势相对平缓,水热条件较好,适宜幼苗、幼树定居和存活,故同属物种山矾和光叶山矾种群结构呈增长态势。
已有研究表明,种群空间分布格局与空间尺度关系密切,种群在小尺度下可能呈聚集分布,而在大尺度下则可能为随机分布或均匀分布[3, 33]。小尺度上,生境异质性可能仅对少量物种的分布有显著影响[34],而在大尺度上生境异质性被认为是产生植被空间分布格局的主要原因[35-36]。本研究中(图4、图5、图6),在完全随机分布模型下,山矾和光叶山矾及其各生长阶段主要为聚集分布,其中山矾及其幼树、小树、中树在较大尺度上由聚集分布向均匀分布过渡,这与前人研究结果一致[29, 37]。Getzin等[10]研究表明在大于10 m的尺度上,如果呈聚集分布,就可解释为受到生境异质性的影响。本研究中在0—100 m尺度上呈聚集分布(图4),各生长阶段除了成年树在r>50 m之后呈随机分布外,幼树、小树和中树也表现出聚集分布特征(图5、图6)。结合空间分布图,可以直观地发现山矾和光叶山矾具有明显的生境偏好。同时,在异质泊松分布模型下,山矾和光叶山矾在不同尺度上从聚集分布过渡到随机分布(图4)。综上,可以看出生境异质性是影响山矾和光叶山矾空间格局的重要因素。
种群个体在各生长阶段存在空间分布格局差异,这使得更好的利用有限资源[38]。一般认为,幼龄个体的分布格局呈聚集分布,随着尺度增加,种群向均匀分布过渡;种群年龄增加,趋向于随机分布[30]。本研究结果符合该规律。另外,我们发现随龄级增加,各生长阶段聚集分布尺度都有下降趋势(图5、图6),这与梁爽等[29]、李立等[39]的研究结果一致。具体表现在幼树、小树阶段很大尺度上呈聚集分布,一方面,可能是母树周围适宜幼龄个体生长的生境;另一方面,受扩散限制影响,大部分种子不会扩散到离母树较远的位置。在中树阶段,仅光叶山矾中树0—150 m尺度上呈聚集分布(CSR),根据已有研究发现[40],除幼龄阶段的自疏外,从中龄林到成熟林,大约有80%个体被稀疏掉,强烈的第二次自疏可能致使光叶山矾中树呈聚集分布。山矾和光叶山矾到成年树阶段,树冠基本定型,根部延伸范围广,资源的获取能力和抵御危害的能力增强,密度制约因素导致的种内种间竞争作用加剧,致使成年树种群密度下降,由聚集分布过渡到随机分布[41-42]。综上,各生长阶段表现出不同的空间格局,有利于植物个体获得充足的资源,是种群的一种生存策略和适应机制[43]。种群空间聚集尺度随龄级的增大而降低,这也是Janzen-Connell空间效应的间接证据之一。
物种各生长阶段的空间关联性可以反映种群的现状和空间分布关系以及个体间的功能关系[44]。本研究中,完全随机分布模型下,山矾种内关联性在较大尺度内呈正关联,随尺度增大,正关联程度逐渐降低表现为无关联;前提条件模型下,仅在小尺度上呈正关联(图7)。而光叶山矾表现出不同的空间关联特征,除小树与成年树在较大尺度无关联外,其余均呈正关联(图8)。虽有研究表明,种群的主要竞争压力来自种内[45],但山矾和光叶山矾各生长阶段的种内关联性几乎没有相互排斥的现象,这可能是同一树种的个体生物学特性的一致性决定的,体现了同属植物利用环境资源的相似性和生态位重叠以及种群内部正向和相互有利的生态关系[46],有利于同属物种种群的共存和发展。
空间分离是物种共存的重要驱动因素之一[47-48],本研究中,山矾和光叶山矾种间关联性的结果显示,完全随机分布模型下,山矾幼树与光叶山矾幼树、小树、中树以及山矾小树与光叶山矾小树几个物种对均呈负关联;而在中树、成年树的物种对间的负关联较弱。前提条件模型下,除山矾幼树与光叶山矾幼树、小树间表现出负关联外,其他物种对呈无关联。究其原因可能是生态位空间隔离导致的。种群的空间分离格局减少不同物种间相遇的几率,减少种间竞争能阻止或减缓竞争优势物种对竞争劣势物种的竞争排除作用[45],有利于同属物种维持共存,提高森林群落生物多样性。
5 结论
湖北木林子国家级自然保护区中,同属物种山矾和光叶山矾均属增长型种群,幼龄个体丰富,种群更新状况良好;空间分布特征随着径级增大,聚集分布向均匀分布过渡;山矾和光叶山矾呈种内聚集、种间分离的空间构型。山矾和光叶山矾作为木林子常绿落叶阔叶混交林动态监测样地的优势树种,正处于稳定增长趋势,不存在生存风险,这对木林子保护区整个森林群落结构稳定和生物多样性维持有重要意义。湖北木林子15 hm2固定监测样地作为亚热带常绿落叶阔叶混交林一个很好的研究平台,可以从多个尺度探讨森林空间格局与生态过程的相互关系。同属植物的空间格局及其种内种间关联性分析只是研究森林空间格局与生态过程之间的关系和物种多样性维持机制的第一步,其他空间格局问题仍需进一步探讨。
