西藏河流裂腹鱼类分布格局及环境影响因素
——以怒江和雅鲁藏布江为例
2020-11-23刘明典马波张驰汤婷刘绍平段辛斌李雷朱峰跃王念民陈大庆
刘明典,马波,张驰,汤婷,刘绍平,段辛斌,李雷,朱峰跃,王念民,陈大庆*
1.中国水产科学研究院长江水产研究所,湖北 武汉 430223;2.中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070;3.西藏自治区农牧科学院水产科学研究所,西藏 拉萨 850000
西藏自治区位于中国西南部,区内江河众多,水系格局极为复杂(张驰等,2014;周建设等,2013)。怒江和雅鲁藏布江是西藏境内两条主要的大型淡水性河流。怒江流域丰富生物资源的原始性、自然性和唯一性使其成为中国生物多样性和天然基因的宝库(董哲仁,2006);雅鲁藏布江是西藏最大的河流,也是世界上平均海拔最高的大河(李海东等,2017),流域气候复杂多样,生物多样性丰富。但近几十年来,随着西藏经济社会的快速发展,怒江和雅鲁藏布江流域的鱼类资源也面临着国内其他水域出现的一些共性问题,如环境污染、过度捕捞、水利设施建设、外来物种入侵等,造成土著鱼类减少,流域水资源开发利用与渔业可持续发展、鱼类资源利用与生态保护的矛盾日益突出(何明华,2005;杨汉运等,2010)。因此,保护怒江和雅鲁藏布江流域的生物多样性和遗传基因具有极其重要的意义。
裂腹鱼亚科(Schizothoracinae)隶属于鲤形目(Cypriniformes)、鲤科(Cyprinidae),特产于亚洲高原地区,其共同特征是腹部肛门附近具有特化形成裂隙的成排鳞片,因此,称之为“裂腹”。在怒江和雅鲁藏布江流域,裂腹鱼类不论在种类数还是数量上都占有绝对优势(张驰等,2014)。裂腹鱼类属于高原冷水性鱼类,其生长远远缓慢于相同纬度低海拔地区的种类,对过度开发和环境破坏等人为干扰相当敏感,资源一旦遭受破坏,其恢复将较为困难,甚至当种群数量降低到一定限度时可能会灭绝(赵利华等,1975;殷名称,1995;陈毅峰等,2002)。但目前有关裂腹鱼类生物学的报道很少,而且大部分集中在年龄、生长或繁殖方面(马宝珊等,2011),关于西藏裂腹鱼多样性分布格局的研究更是缺乏。
在当前鱼类自然资源持续下降、外来物种入侵和人类活动加剧的背景下,对西藏裂腹鱼类的环境影响因素和分布格局进行研究,不仅可为西藏的鱼类学研究和裂腹鱼类种群资源恢复积累基础资料,还可为西藏地区的生物多样性保护和渔业资源管理提供科学指导和理论支撑。
1 研究区域与研究方法
1.1 研究区域
研究区域为怒江干流和雅鲁藏布江干流西藏段,怒江干流共进行5次野外调查,分别为2017年5月、9月和2018年4月、7月、10月。雅鲁藏布江干流进行4次野外调查,分别为2017年4—5月、11月、2018年4—5月、10—11月。基于西藏历史资料和近来已有调查资料,根据生境尺度的形态特征、支流汇入情况、交通便利性、人类干扰程度、宗教信仰与生活习俗等因素设置站点,选择典型河段断面、典型河段样区,同时兼顾空间距离的合理性。怒江干流设那曲、比如、边坝、马利镇、八宿怒江大桥、察瓦龙6个调查站位,支流玉曲设左贡调查站位。雅鲁藏布江干流设桑木张、霍尔巴、拉让、仲巴、萨嘎、拉孜、谢通门、日喀则、仁布、贡嘎、桑日、加查、郎县、米林、派镇、墨脱共计16个调查站位。每个站位调查范围约5 km江段,根据各站位环境状况设置2—3个采样点(图1)。
1.2 研究方法
通过历史资料查阅、标本采集、走访调查等方式,对怒江和雅鲁藏布江鱼类种类组成开展研究,对采集到的标本进行种类鉴定,探明调查江段的鱼类种类组成现状。在各个采样点采用GPS、溶氧仪、流速仪、测距仪、多功能水质分析仪等仪器设备,收集调查水域的非生物环境特征,如经纬度、海拔(m)、水温(℃)、溶氧(mg·L-1)、流速(m·s-1)、电导(μS·cm-1)、pH、叶绿素 a质量浓度(mg·L-1)等参数数据。
同时,在各调查站点采用流刺网和脉冲捕捞方法开展鱼类定量采集,根据各采样点环境特征,选定一定单位的河段采用流刺网(固定)、脉冲捕捞(流动)收集鱼类样本,并结合GPS定位确定雅鲁藏布江鱼类的时空分布特征。聘请渔民借助橡皮艇进行专业捕捞,对采集到的鱼类依据其外部形态特征进行鉴定并记录种类组成。对于形态特征差异极为细微、鉴定结果容易出错的种类采用分子生物学方法鉴定,主要步骤为:采取新鲜状态下的鱼类鳍条样品,用无水乙醇固定保存,带回实验室提取DNA,选择扩增线粒体DNA(mitochondrial DNA)3个基因/片段:线粒体细胞色素 c氧化酶 I基因(cytochromecoxidase I)、细胞色素 b基因(cytochromeb)和控制区(Displacement loop region)进行分类鉴定。
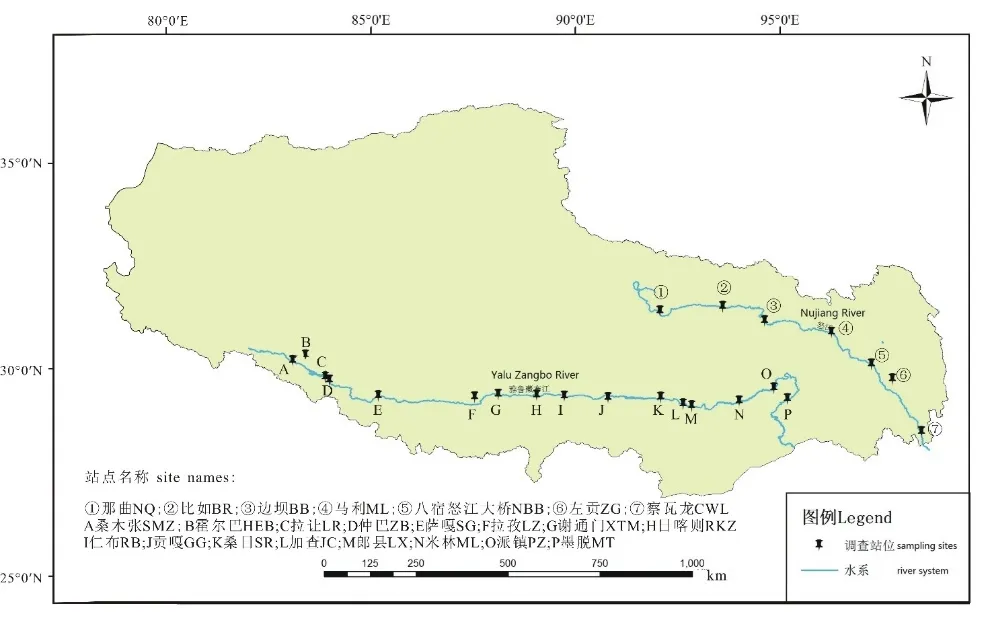
图1 调查站位分布图Fig.1 Distribution map of survey stations
1.3 数据分析
1.3.1 裂腹鱼类分布与环境因子之间的关系
为了了解鱼类物种丰度与环境因子之间的关系,将水温等理化指标作为环境变量,不同鱼类物种的丰度作为物种数据来进行分析。为了优化分析,对物种数据进行lg(x+1)转化,以消除物种出现差异的影响。首先采用除趋势对应分析法(DCA)对物种数据进行分析,以判断采用哪种模型(线性或单峰)更为合适:若length of gradient>4,采用单峰模型,即采用典型对应分析方法(CCA)分析更为合适;若length of gradient<3,则采用线性模型,即采用冗余分析方法(RDA)更合适;如果介于3和4之间,则单峰模型和线性模型均可(Leps et al.,2003)。本研究中怒江和雅鲁藏布江裂腹鱼类群落的 DCA分析结果发现 length of gradient<3,因此,均采用RDA进行分析。利用 Monte Carlo test检验(999 permutations,P<0.05)环境因子对裂腹鱼类空间分布的解释力显著水平。该最少变量组用于最终的RDA分析。数据分析和排序图输出均在R软件中完成,其中DCA分析采用vegan软件包中的decorana函数,RDA分析采用的是vegan程序包中的rda函数。
1.3.2 物种多样性指数
采用以下指数进行鱼类群落的物种多样性分析:
物种丰富度(S)即为群落中的物种数目(王太等,2015)。
Shannon-Wiener多样性指数(H′)(Shannon,1948):

Simpson集中性概率指数(Simpson,1949):

其中,Pi代表第i种个体占总个体数的比例。
1.3.3 物种海拔分布格局分析
在分析物种丰富度随海拔梯度的变化时,以200 m为单位划分海拔带。统计每个海拔带内各裂腹鱼类物种总数。在物种多样性的海拔格局分析中,将每个样点定义为1个群体单元,然后计算物种多样性指数,并对多样性结果进行线性回归分析,以探究多样性指数与海拔的关系。数据分析和排序图输出均在R软件中完成。
2 结果与分析
2.1 裂腹鱼类的物种组成与分布
2017—2018年,在怒江和雅鲁藏布江共发现裂腹鱼亚科鱼类13种,分属裂腹鱼属(Schizothorax)、叶须鱼属(Ptychobarbus)、裸裂尻鱼属(Schizopygopsis)、尖裸鲤属(Osygymnocypris)、裸鲤属(Gymmocypris)。其中怒江发现4种,雅鲁藏布江发现9种(表1),且两条河流中没有共同分布的裂腹鱼种类。统计了13种裂腹鱼类的分布范围(表2),在怒江水域,怒江裂腹鱼(Schigothoraxnujiangensis)除了那曲江段及左贡(支流玉曲)外在其他调查站位均有分布;裸腹叶须鱼(Ptychobarbuskaznzkovi)除了察瓦龙江段外,在其他调查站位均采集到大量个体;热裸裂尻鱼(Schizopygopsisthermalis)主要分布在左贡县、八宿县、马利乡、比如县和那曲县江段;光唇裂腹鱼(Schigothoraxlissolabiatus)仅在察瓦龙江段偶尔可见。
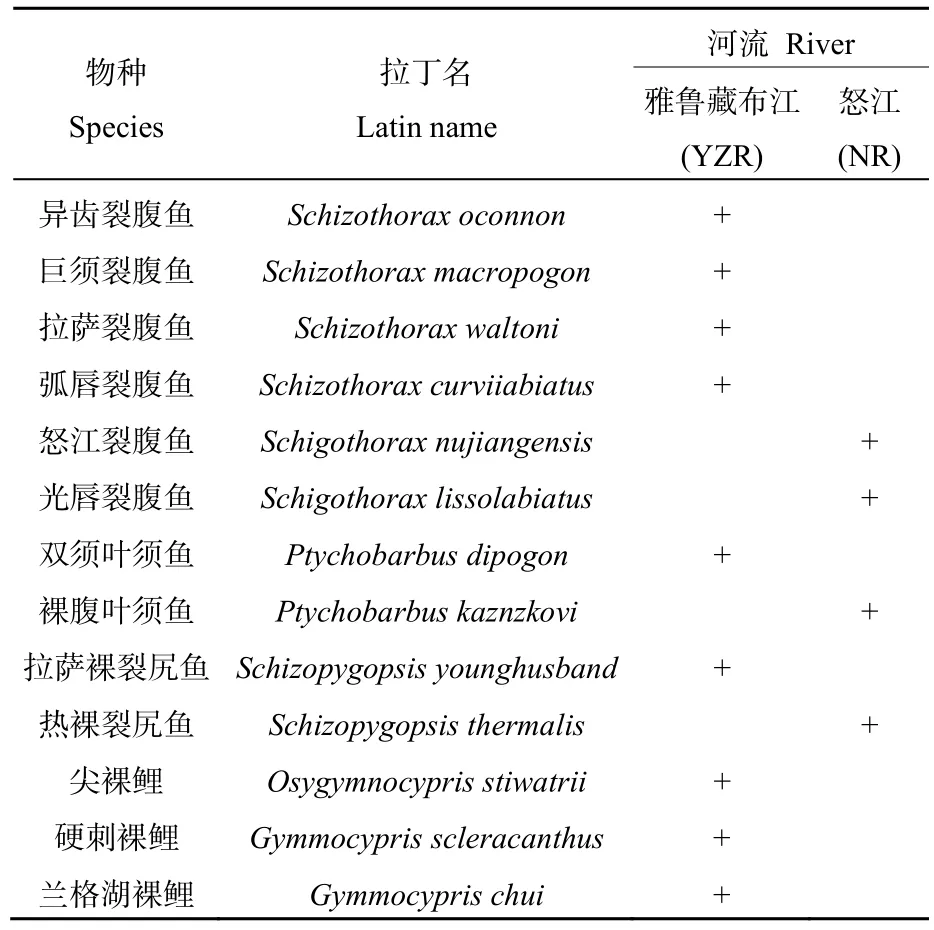
表1 怒江和雅鲁藏布江采集到的裂腹鱼类名录Table 1 A list of Schizothoracinae fishes were collected in the Nujiang River and the Yalu Zangbo River
雅鲁藏布江发现的 10种裂腹鱼类中,异齿裂腹鱼(Schizothoraxoconnon)、拉萨裸裂尻鱼(Schizopygopsisyounghusband)在除墨脱之外的所有调查站位均广泛分布;拉萨裂腹鱼(Schizothorax waltoni)在除墨脱、米林、贡嘎和桑木张4个江段之外的站位都有出现;双须叶须鱼(Ptychobarbus dipogon)在除了墨脱、贡嘎和加查3个江段之外的站位都有出现;巨须裂腹鱼(Schizothorax macropogon)在除了墨脱、贡嘎和霍尔巴江段之外的调查站位均有出现;尖裸鲤(Osygymnocypris stiwatrii)仅分布在仁布以上江段;而弧唇裂腹鱼(Schizothoraxcurviiabiatus)仅出现在墨脱江段;兰格湖裸鲤(Gymmocyprischui)和硬刺裸鲤(Gymmocyprisscleracanthus)仅在2017年调查时在霍尔巴和桑木张江段偶有少量出现。
2.2 影响裂腹鱼类分布的环境变量
冗余分析(RDA)结果表明,怒江与裂腹鱼类分布有关的环境变量主要为水温(P<0.05,R2=0.1981)和流速(P<0.01,R2=0.3636)(图 2),RDA排序轴中,第一、二轴共解释了裂腹鱼类物种丰度数据86.81%的变化,第一排序轴的特征值为0.2608,方差贡献率为62.19%;第二排序轴的特征值为0.1032,方差贡献率为24.61%。从怒江裂腹鱼类与环境变量的 RDA排序图可以看出:怒江裂腹鱼的出现主要与水温呈正相关,与流速呈一定程度
的正相关;裸腹叶须鱼的出现主要与流速呈正相关;热裸裂尻鱼的出现与水温呈负相关。
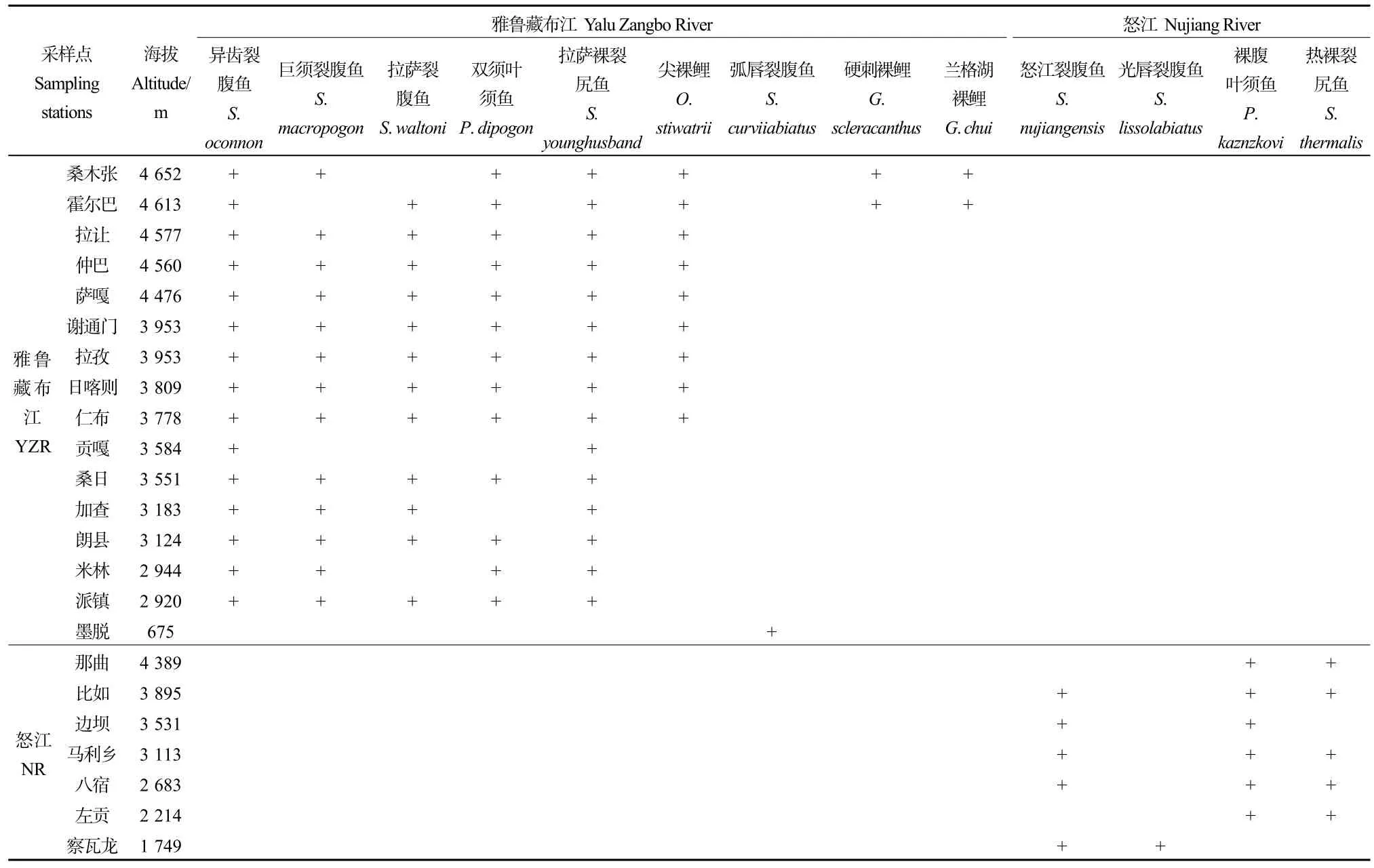
表2 雅鲁藏布江和怒江2017—2018年裂腹鱼类分布范围Table 2 Distribution of Schizothoracinae fishes were collected in the Nujiang River and the Yalu Zangbo River from 2017 to 2018
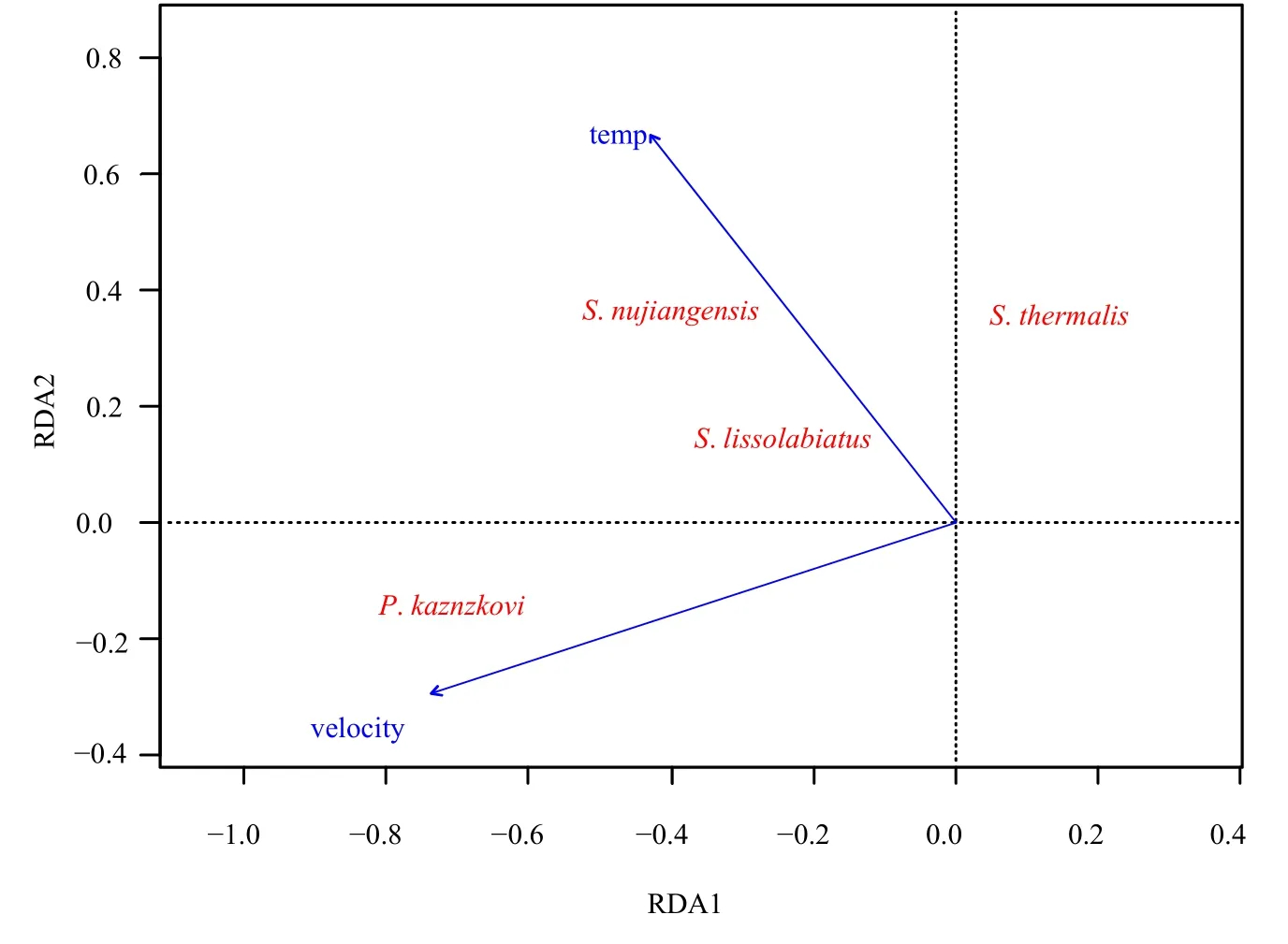
图2 怒江裂腹鱼类分布与环境变量之间关系的RDA排序图Fig.2 Redundancy analysis (RDA) ordination diagrams of the presence of Schizothoracinae fishes and environmental variables in Nujiang River
雅鲁藏布江与裂腹鱼类分布有关的环境变量主要为海拔(P<0.01,R2=0.7605)、水温(P<0.01,R2=0.2746)、Chla浓度(P<0.05,R2=0.2365),电导与裂腹鱼类分布之间的关系不显著(P<0.1,R2=0.1713)(图3)。RDA排序轴中,第一、二轴共解释了裂腹鱼类物种丰度数据88.60%的变化,第一排序轴的特征值为3.8441,方差贡献率为65.93%;第二排序轴的特征之为 1.3220,方差贡献率为22.67%。从雅鲁藏布江裂腹鱼类与环境变量的RDA排序图可以看出:异齿裂腹鱼、拉萨裂腹鱼和巨须裂腹鱼的出现与水温呈正相关;双须叶须鱼、拉萨裸裂尻和尖裸鲤的出现与海拔和叶绿素 a呈正相关,与水温呈负相关;弧唇裂腹鱼的出现与海拔呈负相关;硬刺裸鲤的出现与叶绿素a和海拔呈一定的正相关,兰格湖裸鲤以及尖裸鲤和拉萨裸裂尻鱼杂交种的出现与环境变量之间的相关性不大。
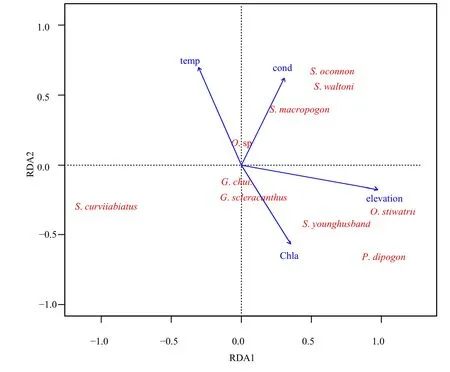
图3 雅鲁藏布江裂腹鱼类分布与环境变量之间关系的RDA排序图Fig.3 Redundancy analysis (RDA) ordination diagrams of the presence of Schizothoracinae fishes and environmental variables in Yalu Zangbo River
2.3 物种多样性指数
对怒江和雅鲁藏布江裂腹鱼类的 Shannon-Wiener指数和Simpson指数分析结果显示:雅鲁藏布江和怒江裂腹鱼类的 Shannon-Wiener指数分别为 (1.06±0.49)、(0.51±0.31),Simpson 指数分别为(0.45±0.23)、(0.69±0.20)。雅鲁藏布江裂腹鱼类的多样性明显高于怒江。
2.4 裂腹鱼类物种多样性的海拔分布格局
怒江和雅鲁藏布江所有调查站位的海拔范围在675—4652 m之间,按照200 m的间隔划分海拔带,共可分成21个海拔区间。从裂腹鱼类物种数随海拔梯度的变化图(图 4)可以看出:随着海拔的上升,裂腹鱼类物种丰富度呈现先缓慢增加后减少的变化状态,在3700—3900 m海拔范围内裂腹鱼种类最多。不同属的物种在西藏的分布海拔有一定差异,如裂腹鱼属(弧唇裂腹鱼除外)、叶须鱼属、裸裂尻属鱼类主要分布在 1700—4650 m海拔范围;尖裸鲤属分布在3700—4650 m海拔范围;裸鲤属分布在4600—4650 m范围(表2)。
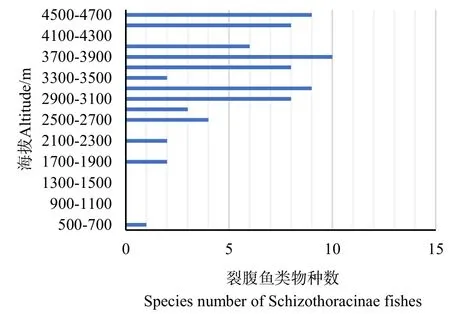
图4 裂腹鱼类物种数随海拔梯度的变化Fig.4 Variation of richness of Schizothoracinae fishes with altitude gradient
Shannon-Wiener指数与海拔的回归结果(图5A)显示,雅鲁藏布江(y=0.00034x-0.18,R2=0.4639,P<0.0001)以及雅鲁藏布江与怒江整体(y=0.00024x-0.041,R2=0.1964,P<0.001)裂腹鱼类的Shannon-Wiener指数随海拔的升高呈上升趋势。
Simpson指数与海拔的回归结果显示(图5B),雅鲁藏布江(y= -0.00016x+1.026,R2=0.4513,P<0.0001)以及雅鲁藏布江与怒江整体(y=-0.0001x+0.93,R2=0.1512,P<0.01)裂腹鱼类的Simpson指数随海拔的升高呈下降趋势。
综合上述结果可知,西藏东南部河流中裂腹鱼类的物种多样性随海拔的上升呈先上升后下降的趋势,裂腹鱼类的物种丰富度在3700—3900 m海拔区间内达到最高。
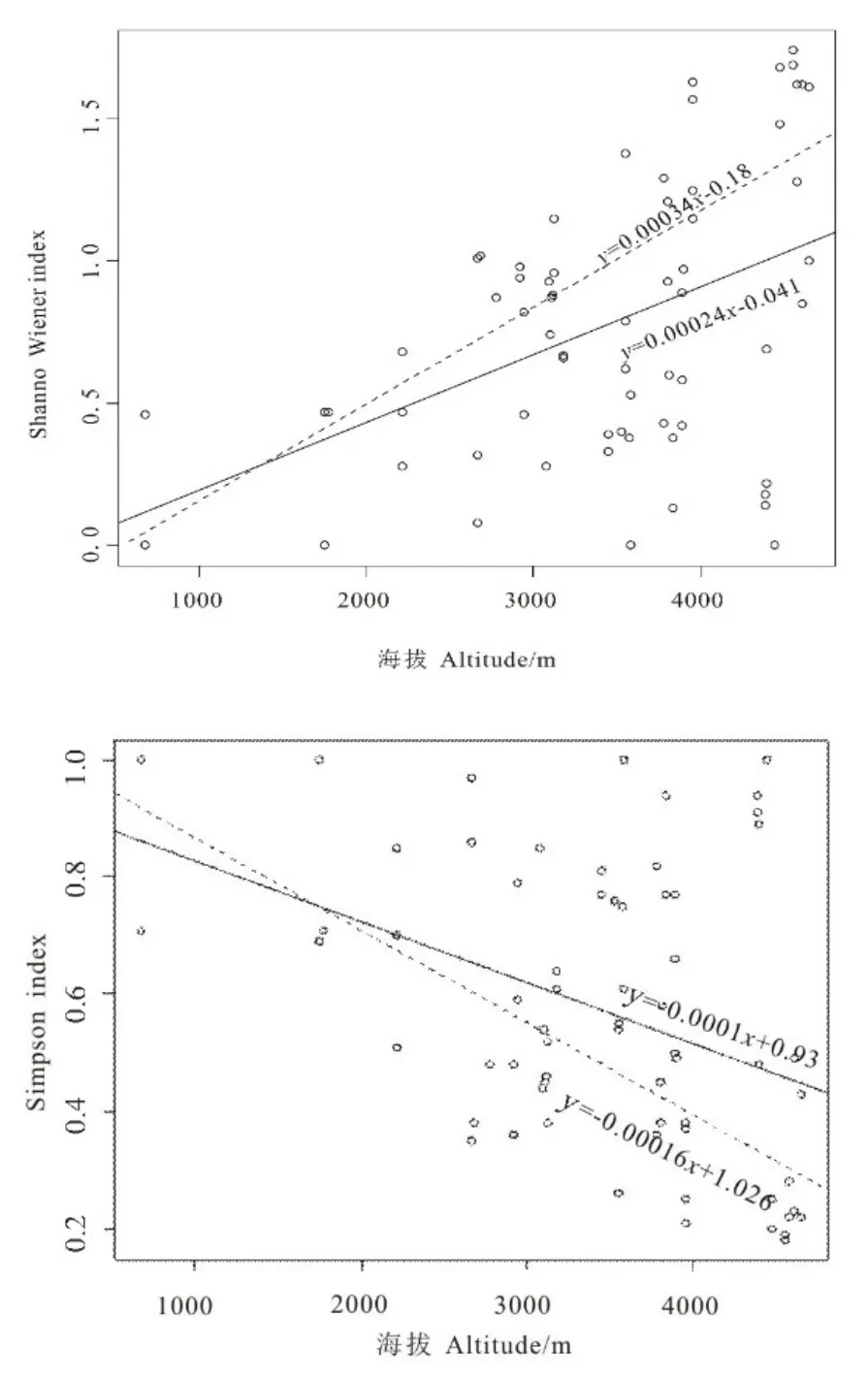
图5 Shannon-Wiener指数和Simpson指数与海拔的关系Fig.5 Relationships between diversity index and altitude
3 讨论
3.1 裂腹鱼类分布特征变化
目前,国内关于裂腹鱼类分布特征的研究报道较少。仅有四川裂腹鱼(Schizothoraxkozlovi)产卵场的分布特征(陈永祥等,1997),影响裂腹鱼类自然分布的河流水力学特征(王玉蓉等,2010)以及雅鲁藏布江中游拉萨裂腹鱼(Schizothoraxwaltoni)、拉萨裸裂尻鱼(Schizothoraxyounghusbandi)等种类分布和栖息地特征(朱挺兵等,2017)研究等少数报道。本研究区域主要为高寒气候,且区域内宽、峡谷相间,河道形态复杂多变,生境高度多样化,各调查江段中均有裂腹鱼类分布。根据研究报道,左贡县江段是怒江裂腹鱼的主要分布区域,边坝县江段是热裸裂尻鱼的主要分布区域(王龙涛,2015)。但本研究中,2017—2018年度5个季度的调查中,怒江裂腹鱼在左贡县江段都没有出现,边坝县江段也均未采集到热裸裂尻鱼。推测这两种裂腹鱼类分布区域的变化可能与近几年来人类活动造成的干扰因素有关。
怒江上游人烟稀少,工农业不发达,除了水温、流速、溶氧等的变化以及人口相对密集的村、镇生活污水的排入导致的水质变化等对裂腹鱼类的分布造成不利影响外(杨汉运等,2010),裂腹鱼类还容易受怒江上游水电开发导致的河道地形、地貌、水文、泥沙等条件的改变带来的不利影响(杨剑等,2010)。也有研究发现,在怒江支流建有小型水电站的河流,鱼类受水电站建设的影响很大:特别是半洄游性的裂腹鱼类数量减少,渔业资源量下降(王龙涛,2015)。本研究中有关人类活动对西藏东南部水体裂腹鱼类生境影响的研究有限,但可以预见的是,随着经济的发展和人类活动开发强度的不断增加,研究区域内裂腹鱼类可能会面临栖息地丧失、物种难以延续的局面。因此,未来有关西藏裂腹鱼类的研究有必要加强对人类活动影响评估的研究,以便为有关管理者制定裂腹鱼类保护策略提供科学指导。
3.2 影响裂腹鱼类分布的环境因素
有关环境因素对鱼类生存影响的研究是国际淡水群落生态学领域的焦点之一(Matthews,1982),国内外已有的对鱼类分布特征与环境因子之间关系的研究表明,水质状况(Perna et al.,2005;Wexler et al.,2011)、水文条件(Vinagre et al.,2009)、栖息地特征(Wright et al.,2004)等自然环境因素对鱼类的分布状况具有显著影响。例如,有学者对淠河流域河源溪流影响鱼类空间分布格局的影响因素研究分析发现,影响鱼类物种数及个体数的参数包括海拔、流速、底质、水深、河宽、电导率、底质等环境因素(张晓可等,2017);此外,在澜沧江流域的调查中发现,鱼类分布主要受海拔、气候的影响(刘明典等,2011);新疆塔里木河水系鱼类随着海拔梯度变化而呈现出垂直分布特点(马燕武等,2009)。
基于冗余分析法(RDA)分析裂腹鱼与环境因素关系的结果表明,怒江和雅鲁藏布江裂腹鱼类的分布均与水温具有显著的相关关系。由于水温的变化受海拔变化的直接影响,高海拔地区水体的水温常常比低海拔地区的水体低(陈自明等,2006),因此,推断研究区域内裂腹鱼类的分布与海拔(亦即水温)之间存在一定的相关关系。这一推断在怒江和雅鲁藏布江也得到印证(图 2和图3),只是有的种类分布数量与水温出现正相关,有的却是负相关。可能与不同裂腹鱼适宜生活的水温范围不同有关,如拉萨裸裂尻鱼幼鱼、异齿裂腹鱼幼鱼和双须叶须鱼幼鱼摄食水温分别为8—27、8—25、7—26 ℃(曾本和等,2019),鱼类对水温的耐受幅度及边界值的不同会导致鱼类分布江段的不同,而最适宜生长水温将决定哪些江段鱼类数量较多,如异齿裂腹鱼生长最适宜水温范围为15.15—17.24 ℃(曾本和等,2018)。此外,裂腹鱼类与水温的正负相关性与特化程度有一定关联。本次分析中,分布于怒江的怒江裂腹鱼以及分布于雅鲁藏布江的异齿裂腹鱼、拉萨裂腹鱼和巨须裂腹鱼的数量均与水温呈正相关,这 4种属于裂腹鱼属,为原始等级类群(曹文宣等,1981);分布于怒江的热裸裂尻鱼以及分布于雅鲁藏布江的双须叶须鱼、拉萨裸裂尻鱼和尖裸鲤的数量与水温呈负相关,叶须鱼属为特化等级类群,尖裸鲤属和裸裂尻鱼属为高度特化等级类群。不同特化程度的裂腹鱼营养及空间生态位出现分化(王起等,2019),或许可以解释这一现象。科学的解释还需要进一步分析论证。
除了海拔和水温,其他因素也会影响裂腹鱼的分布。本研究在雅鲁藏布江发现的 10种裂腹鱼类物种中,异齿裂腹鱼、拉萨裸裂尻鱼在除墨脱之外的所有调查站位均广泛分布;尖裸鲤仅分布在仁布以上江段,这一结果与朱挺兵等(2017)在雅鲁藏布江中游的研究结果一致。但在环境影响因素方面,本研究与其研究存在一定差异。根据上文的分析结果,雅鲁藏布江流域与裂腹鱼类分布有关的环境变量主要为海拔、水温和Chla浓度,pH不是主要的影响因素。这一差异一方面可能来源于研究区域上的不同,本研究范围覆盖了中国境内雅鲁藏布江整个干流江段,涵盖了海拔梯度变化明显的上游和环境差异较大的中游,研究结果更容易体现出差异和梯度;另一方面可能与数据分析采用的水体理化指标体系上的差异有关。目前有关雅鲁藏布江裂腹鱼类分布的环境影响因素研究较少,无法进一步地比较分析。对于地处青藏高原的怒江和雅鲁藏布江,其受外界因素的干扰相对较小。亚洲中部高海拔地区鱼类与其他区域显著不同,主要是适应急流和高海拔的冷水物种。有研究报道历史因素是造成裂腹鱼等高原鱼类目前分布模式的主要驱动因素(He et al.,2020),包括青藏高原的隆升、邻近造山运动以及随之而来的东亚季风气候的形成,对青藏高原鱼类分布产生了深远影响,而这些历史因素在河流环境最直接的体现就是海拔和水温的差异性。
3.3 西藏裂腹鱼类多样性的海拔格局
物种多样性随海拔梯度的变化是生物多样性保护研究的重要内容(Vetaas et al.,2002;Parmesan,2006)。根据本研究的结果,裂腹鱼类的物种丰富度存在随海拔升高而先增加后减少的趋势,在3700—3900 m海拔范围内达到最高,这一结果与Fu et al.(2004)的结果类似,说明裂腹鱼类物种丰富度随海拔梯度增加呈现单峰曲线分布趋势,在特定的海拔范围内最高。但需要注意的是,本研究中裂腹鱼类的物种丰富度峰值并不是中海拔处,而是 3700—3900 m这一相对较高的海拔范围,这可能与研究区域内调查站位分布不均衡有关。研究区域虽然跨越了较大的海拔梯度,但受实际调查工作中采样时间和经费限制,调查站位主要集中在干流江段,支流江段作为研究区域河流流域的重要部分,其裂腹鱼类的分布情况尚未可知。因此,下一步的研究工作中,有必要针对支流江段的裂腹鱼类资源状况进行调查与分析,以对本研究的结果进行补充和验证。
多样性指数的海拔回归分析结果发现,裂腹鱼类的物种多样性指数随海拔的升高呈上升趋势。这一结果与陈自明等(2006)在独龙江的研究结果存在较大差异,独龙江流域冬季鱼类多样性指数随海拔上升而下降。两者在鱼类种类和研究区域上都与本研究不同,无法进行深入比较分析。由于目前关于裂腹鱼类多样性分布格局的研究较少,本研究结果可为裂腹鱼类资源的保护和管理提供重要的基础资料和理论参考。
4 结论
西藏河流裂腹鱼类的分布区域性较强,怒江和雅鲁藏布江没有共同分布的裂腹鱼种。裂腹鱼物种丰富度随着海拔的上升呈现先增加后减少的趋势,在3700—3900 m海拔范围内裂腹鱼种类最多。不同属的物种在西藏的分布海拔有一定差异,裂腹鱼属、叶须鱼属和裸裂尻属鱼类主要分布在 1700 m以上海拔区域,尖裸鲤属分布在3700 m以上,裸鲤属分布在4600 m以上。裂腹鱼类的分布与水温存在显著的相关关系,不同特化程度的裂腹鱼与水温相关性有所不同。
