野亚麻(L.stelleroides Planch.)的染色体核型分析
2020-11-20苗红梅王玉祥马建富李爱荣乔海明
刘 栋,苗红梅,马 琴,王玉祥,马建富,郭 娜,李爱荣,乔海明
(1.张家口市农业科学院,河北 张家口 075000; 2.河南省农业科学院芝麻研究中心,郑州 450000)
根据用途和形态特征亚麻可分为纤维用型、油用型和油纤兼用型亚麻。油用型亚麻(胡麻)是我国西北及华北地区的重要油料作物之一,种植历史悠久。亚麻油富含亚麻酸和亚油酸等不饱和脂肪酸;亚麻籽中含有木酚素等多种功能活性成分。亚麻油及相关加工产品深受消费者喜爱,因亚麻抗旱和耐瘠性突出,亚麻产业在高效旱作农业以及种植业结构调整中起着至关重要的作用。
但亚麻作为小宗作物,研发经费投入不足,相关基础研究落后,特别是在染色体核型方面的研究较少。染色体核型分析是细胞遗传学研究的基本方法,是研究物种演化、分类以及染色体结构、形态与功能之间的关系不可缺少的重要手段[1]。国外亚麻属的核型分析始于半个多世纪前,到目前为止有多个亚麻种的核型已经明确[2-4]。相较于国外,国内进行亚麻属染色体核型分析的研究起步较晚,且进展较慢。仅有少量研究针对亚麻的染色体数量和核型特征进行了观察和分析[5-10],且所用材料多为栽培种。因此,开展野亚麻核型分析研究,既可以为亚麻野生种的鉴定、分类、遗传和演化研究提供丰富的细胞学信息,又可为今后利用种间杂交技术创制新种质提供选择依据。
1 材料与方法
1.1 材 料
试验于2019年1月在河南省农业科学院芝麻研究中心实验室进行。野亚麻(L.stelleroidesPlanch.)种子于2018年9月采自河北省张家口市崇礼区高家营镇把图湾村S 242道路南边山坡上,地理坐标为115°11′6″E,40°54′8″N,海拔高度1 180~1 300 m,野亚麻形态特征见图1。

注:1为原生境;2为开花植株;3为花瓣;4为根;5为萼片边缘有黑色球形腺体;6为蒴果和种子。 图1 野亚麻原生境地及形态特征
1.2 材料处理及制片
采用酶解去壁低渗法进行染色体制片,在陈瑞阳等[11]的研究方法基础上进行了优化,具体步骤为: 1) 将亚麻种子放入50 mL离心管中,加入20 mL蒸馏水,离心管平放,固定在摇床上,28 ℃条件下恒温培养,待根尖长至1~2 cm时,选健壮根尖取下备用;2) 预处理。将根尖直接浸泡在0.002 mol·L-18-羟基喹啉溶液中,4 ℃条件下过夜处理; 3) 固定。将根尖转移到固定液中,固定2~3 h; 4) 前低渗。切取发白的根尖分生区组织,用水冲洗30 min; 5) 酶解。将尽量干的根尖置于2.5%果胶酶和2.5%纤维素酶等量混合液中,在杂交炉中37 ℃条件下避光处理过夜;6) 后低渗及固定。将根尖放在蒸馏水中冲洗2~3次(约30 min),于固定液中4 ℃条件下处理2 h以上; 7) 涂片。取根尖,放在清洁的载片上,加固定液,用镊子将根尖敲碎。滴加少许吉姆萨工作液,染色10~15 min,用流水冲洗载片,待干燥后即可镜检。用Leica DM 6000 B光学显微镜观察根尖细胞染色体,100倍油镜下对分裂中期细胞染色体进行拍照。
1.3 镜检及核型分析
选取30个染色体分散较好的细胞,统计各细胞染色体数目。以5个细胞各项指标平均值计算核型参数值,核型分析参照文献[12]的方法,利用photoshop CS 5对清晰的染色体中期分裂图像进行核型分析,核型分类参照Stebbins等[13]的分类标准,采用Arano[14]的方法计算核型不对称系数,按照乔永刚等[15]的方法,绘制核型模式图。
2 结果与分析
2.1 野亚麻染色体数目
对染色体分散较好的30个细胞进行观察计数,所有观察的细胞染色体数目均为2 n=2 x=20,未发现染色体数异常细胞,具体见图2。
2.2 野亚麻染色体核型分析
选取5张清晰的染色体中期图片进行核型分析,野亚麻的染色体数为20条,相对长度变幅为3.79~6.40,染色体臂比大于2的数量为0,最长染色体与最短染色体长度的比值为1.68,核型不对称系数为54.15%,属于1 A类型(见表1和图2)。20条染色体全部为中部着丝点染色体(m)。野亚麻的核型公式为2 n=2 x=20=20 m。
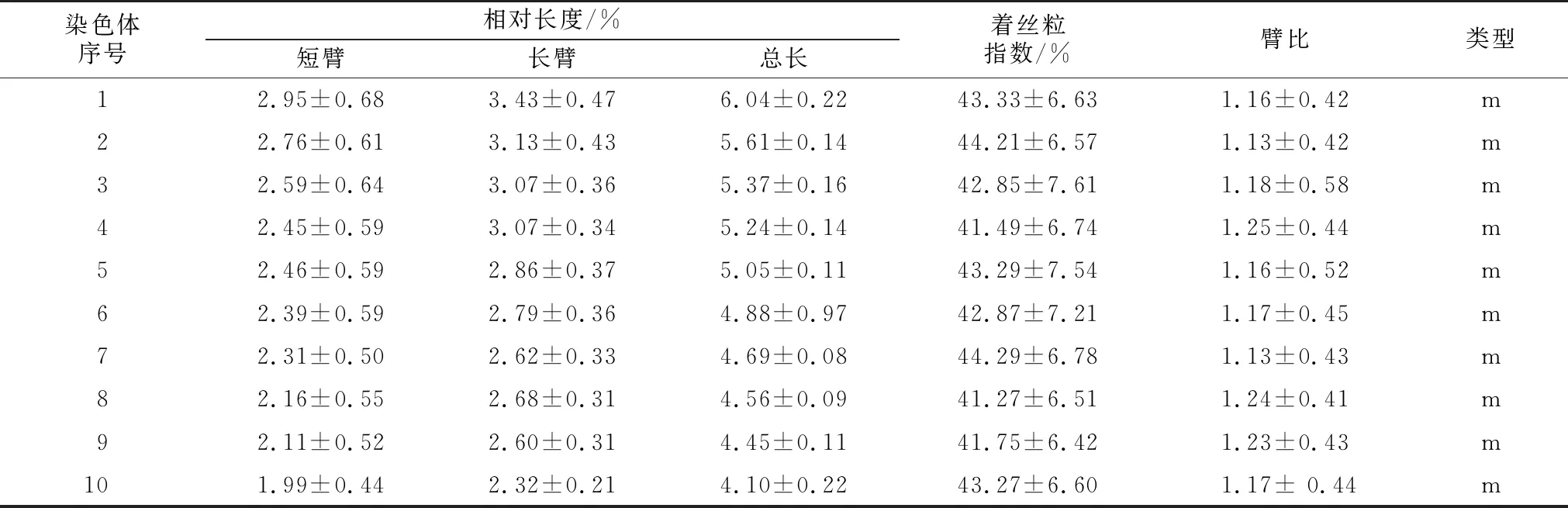
表1 野亚麻(L.stelleroides Planch.)核型参数表
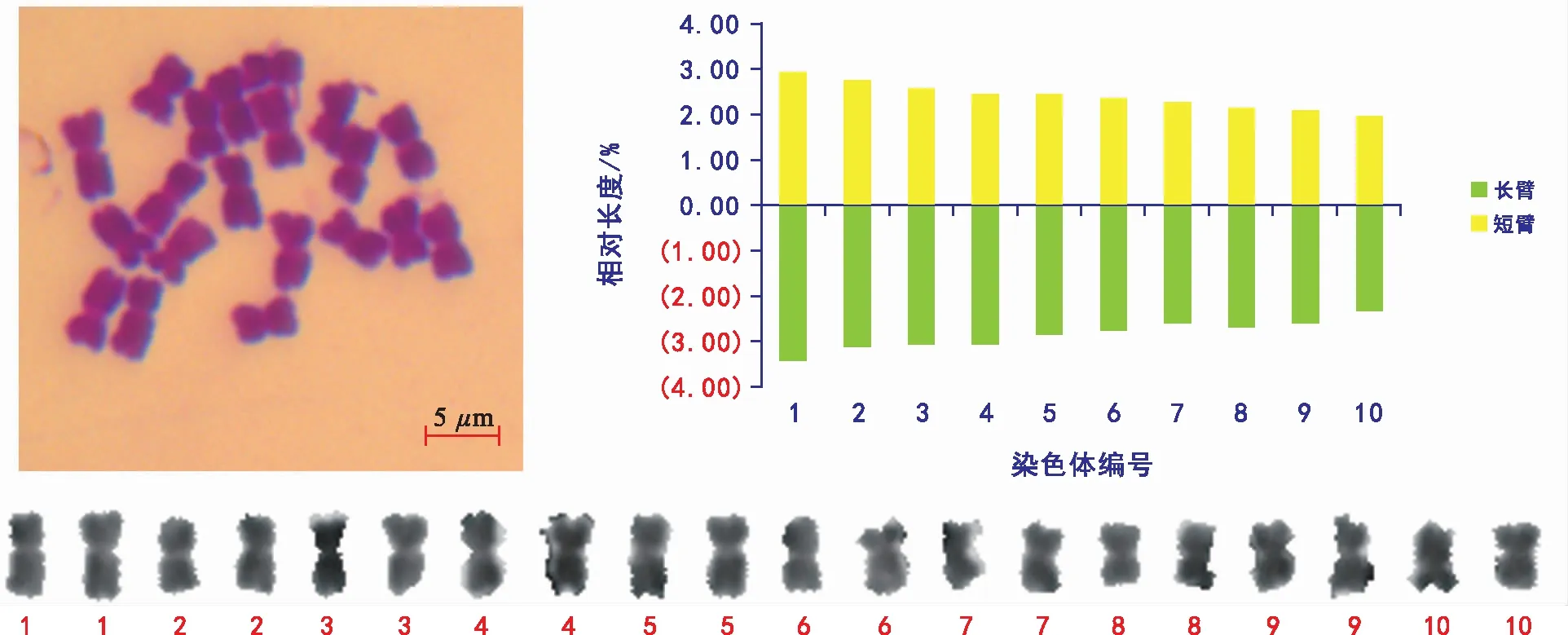
图2 野亚麻的染色体形态特征及核型模式图
3 讨 论
亚麻科(Linaceae)有22个属,其中亚麻属(Linum)约200个种,主要分布于世界的温带和亚热带[16]。据中国植物志[17]记载,亚麻属在中国约有9种,分布于西北、东北、华北和西南等地,其中包括野亚麻(L.stelleroidesPlanch.)。而据河北植物志[18]记载,亚麻属在河北有3个种,分别为栽培亚麻(L.usitatissiumL.)、宿根亚麻(L.perenneL.)和野亚麻(L.stelleroidsPlanch.)。荣冬青等[19]于2016年报道,在张家口赐儿山发现垂果亚麻(L.nutansMaxim)后,河北省的亚麻种数量增加至4种(见分种检索表),其中宿根亚麻(L.perenneL.)[20-22]和垂果亚麻(L.nutansMaxim)[19,23]经过开发并在育种中得到利用,而野亚麻(L.stelleroidsPlanch.)只在陕西七里川[24]和长白山[25]被发现并采集,在河北省内只有书面记载,尚未有其原生境介绍以及植株与种子被采集的相关报道。
河北省亚麻属分种检索表[18]如下:

亚麻野生种具有抗旱、抗寒、耐盐碱、耐瘠薄等特点,是胡麻品种改良的重要基因来源[26]。对野亚麻原生境地的调查以及考种数据表明,该种不仅抗旱耐瘠薄,且产量性状优异,特别是单株蒴果数最高可达830个。此次在张家口崇礼区发现的野亚麻(L.stelleroidsPlanch.),丰富了野生种质资源库,为下一步开展抗逆育种提供了优异基因资源,同时为研究野生种和栽培种种间遗传多样性、野生资源优异基因定位和克隆以及亚麻起源与进化等奠定了良好的材料基础。
亚麻属染色体数量(2 n=16,18,20,26,28,30,32,36,42,72,84)存在广泛的变异,这说明染色体变异在亚麻种的形成过程中起着关键的作用[27]。对亚麻属染色体数量的研究常伴着对其核型特征的研究。目前为止,关于国内的8个亚麻野生种的染色体数量以及核型的研究还未见相关报道。国内只有少量针对栽培亚麻的核型特征进行的研究,结果显示,栽培种的染色体条数为30或者32,核型类型多为不对称的“2 B”类型,是一种不对称程度较高的染色体组,在系统发生上属于比较进化的类型。另外,吴广文等[28]对引自波兰的多年生野生种黄花亚麻进行了核型分析,发现该野生种亚麻染色体有28条,且核型类型为“2 A”型,而针对国内亚麻野生种的核型特征研究尚属空白。
本研究结果表明,野亚麻为二倍体,其核型公式为2 n=2 x=20=20 m,核型类型为1 A。比较本研究中野亚麻与栽培亚麻,发现该种与栽培种在染色体数量、大小和核型特征方面均具有较大差异。根据Stebbins等[13]的观点,植物系统演化上处于比较古老或原始的植物,大多具有较对称的核型,而不对称的核型则常见于衍生的或进化较高级的植物中。据此,野亚麻的核型属于对称的类型,推断该种在亚麻系统发生中处在较原始的位置。但要明确亚麻起源、演化的整个过程,需在将来的工作中尽可能多的收集国内外的亚麻野生种,在材料丰富的基础上,从染色体和基因组水平上研究不同种间的核型特征和基因序列的遗传多样性,从而推测和判断亚麻的起源与演化路径。
4 结 论
本研究报道了亚麻属野生种野亚麻(L.stelleroidesPlanch.)的染色体数量和核型特征,该结果可为研究栽培亚麻起源、演化和野生种的利用提供科学依据。
