烟草野生种eeIIFF44EE11--SS 同源基因的多样性与马铃薯Y病毒抗性分析
2021-08-04黄昌军陈学军于海芹袁诚刘勇
黄昌军,陈学军,于海芹,袁诚,刘勇
云南省烟草农业科学研究院/烟草行业烟草生物技术育种重点实验室/国家烟草基因工程研究中心,昆明,650021
马铃薯Y病毒病(Potato virus Y,PVY)是我国烟草上的重要病害,每年造成的损失位列十大烟草侵染性病害名单前列[1-2]。种植抗病品种是防控PVY最根本和最经济有效的措施,但我国烤烟主栽品种云烟87、K326、红花大金元、云烟97和中烟100等均不抗PVY[3]。优良抗源是选育高抗且丰产优质品种的关键之一。目前烟草育种利用的PVY抗源主要来源于普通烟草(Nicotiana tabacum)Virgin A mutant(VAM),是由Virginia A 经X-射线辐射诱变而来[4]。诱变导致包含eIF4E1-S基因的染色体片段缺失(以va表示)。普通烟草eIF4E1-S突变也能获得PVY抗性[5]。普通烟草为异源四倍体,具有N.sylvestris和N.tomentosiformis两套基因组,其中eIF4E1-S(感病基因)来源于N.sylvestris,可被PVY利用,完成病毒的侵染循环;同源基因eIF4E1H-T(非感病基因)来源于N.tomentosiformis,不能被PVY利用[6]。刘勇等根据eIF4E1-S基因及其家族基因的序列信息,开发出分子标记CF2GR11,利用该标记明确普通烟草常见抗源缺失eIF4E1-S基因[7]。普通烟草中PVY抗源稀少[8-9],而烟草野生种中PVY抗源较多。Sievert(1972)[10]采用一个PVY分离物接种62个野生种、14份可育的种间杂交种和近1000份TI编号的美国外引种质,筛选出11个野生种和TI 1406抗PVY,其他材料均感PVY。Burk(1982)[11]采用3个PVY株系PVY MSMR、MSNRNSNR接种,从40个野生种、99个烟草品系中,筛选出5个野生种对3个株系免疫、1个品系(VY32)具有对PVY的抗性。Dijk and Cuperus(1989)[12]采用3个PVY株系PVYO、PVYN、PVYC的3个分离物鉴定了60个烟草野生种的抗性。提出安第斯山脉中部(PVY病毒可能起源地)的圆锥烟草组(Paniculatae)的野生种对PVY的抗性较强。Doroszewska and Depta(2011)[13]采用3个PVY株系PVYNW、PVYNZ、PVYNTN(分别包含可克服和不能克服va抗性分离物)鉴定了包含68个野生种的96份资源的抗性。其中N. raimondii,N. knightiana,N.glauca N. benavidesii,N.wigandioides,N. noctiflora和N.africana抗所有实验株系。由于PVY分离物不同和种质的“同名异物”现象,不同研究者对野生种的PVY抗性评价结果可能存在一些差异,但普遍认为野生种中存在较多PVY抗源。发掘利用上述野生种中的PVY抗性,可丰富栽培烟草抗PVY基因资源。
由于野生种与普通烟草的染色体倍性和数量存在较大差异,野生种与普通烟草之间普遍存在生殖隔离,将野生种的抗性转育至普通烟草中难度大、工作量大。因此,首先确定野生种中是否存在抗PVY新基因,再对包含抗PVY新基因的野生种进行抗性转育,可显著提高育种效率。烟草野生种PVY抗性包括三种可能:感病基因eIF4E1-S缺失/突变、存在新抗病基因、二者共存。普通烟草eIF4E1-S蛋白能被PVY利用而完成病毒侵染循环(eIF4E1-S可当成感病基因),eIF4E1H-T蛋白不能被PVY利用(eIF4E1H-T可当成“非感病基因”),植物感马铃薯Y病毒属病毒(Potyviruses)的eIF4E蛋白存在保守的关键结构域(loop1和loop2)[5,14]。可通过比较烟草野生种loop1和loop2氨基酸序列来判断是否含有感病基因。本文采用普通烟草eIF4E1-S和eIF4E1H-T基因cDNA特异引物扩增测序,分析了34个野生种eIF4E1同源基因的多样性、感马铃薯Y病毒属病毒关键结构域(loop1和loop2)氨基酸差异;鉴定了PVY抗性。提出假说“在存在eIF4E1-S感病基因的情况下,抗PVY野生种携带抗PVY新基因的可能性高;在不存在eIF4E1-S感病基因(包括存在eIF4E1H-T)的情况下,抗PVY野生种携带抗PVY新基因的可能性较低”。根据假说预测了抗PVY野生种含有抗PVY新基因的可能性。
1 材料与方法
1.1 试验材料
34个烟草野生种由云南省烟草农业科学研究院保存(见表1)。野生种采用漂浮盘育苗,3~4片叶时,移栽到装填育苗基质的直径10 cm的塑料花盆中。4-5片叶时,采集幼叶提取核酸。
1.2 抗性鉴定
在防虫塑料大棚内进行。按常规方法漂浮育苗,在15孔的苗盘中,每份资源定植15株苗,苗龄60 d左右,采用高压喷枪摩擦接种PVY坏死株系(ZT-5)。方法如下:采集经免疫检测试纸条检测为PVY的新鲜病叶,在无菌研钵中,加接种缓冲液(0.01 mol/L pH 7.0磷酸盐缓冲液)研磨,双层纱布过滤,调整接种液的浓度至1:100重量体积比,在病叶汁液中加入1%过孔径0.075 mm筛孔的化学纯二氧化硅。高压喷枪喷射压力为1 kg/cm2,喷枪口距离接种叶20 cm左右,每株苗喷射接种时间约为0.5 s。接种后立即用喷壶喷少量自来水,提高接种效率。烟苗接种后14 d开始调查发病情况,每7 d一次,连续调查3次,计算发病率。最后一次调查发病率小于或等于35%判断为抗病;发病率大于或等于50%判断为感病。发病率在35%~50%之间的野生种判断为中抗,本文剔除了中抗的野生种。部分野生种的PVY抗性引自Doroszewska(2011)[13],R表示抗PVYNW(-)分离物和/或PVYNTNPol(-)分离物,S表示感PVYNW(-)分离物且感PVYNTNPol(-)分离物。
1.3 核酸提取与cDNA合成
每份野生种挑选3个单株,分别取幼叶提取DNA和RNA。DNA提取采用CTAB法或者Qiagen DNA 提取试剂盒。RNA提取采用Trizol方法。采用Nanodrop测定DNA浓度和质量。将质量和浓度合格的DNA稀释至100 ng/μL备用。采用TAKARA公司的反转录试剂盒,进行cDNA合成。
1.4 eIF4E1引物设计与PCR扩增
eIF4E1-ScDNA(GenBank序列号KF155696.1,MN896999) 全 长660 bp。eIF4E1H-T的cDNA(GenBank序 列 号KM202068;MN897010)全 长657 bp。通过比对eIF4E1-S和eIF4E1H-T的cDNA序列,设计可特异扩增eIF4E1-ScDNA全长的引物CF3/CR3,上游引物CF3位于eIF4E1-ScDNA起始密码子ATG前第15-37个碱基, 下游引物CR3位于eIF4E1-ScDNA起始密码子后第654-674个碱基。同样设计扩增eIF4E1H-T同源基因cDNA部分序列的引物CF4/CR4:上游引物CF4位于eIF4E1H-TcDNA起始密码子ATG前第22-67个碱基,下游引物CR4根据eIF4E1H-T的cDNA序列起始密码子ATG后第608-629位碱基设计。CF3/CR3可特异扩增出eIF4E1-S,CF4CR4可同时扩增出大部分供试野生种eIF4E1同源基因。对野生种首先采用CF3/CR3扩增,若CF3/CR3扩增阴性则采用CF4/CR4扩增。剔除CF3/CR3和CF4/CR4都无扩增产物的野生种。
扩增eIF4E1基因cDNA全长序列的引物对为CF3:5′-TAAAATCTATAACTAAGTACATA-3′,CR3:5′- CCATTGTAGCAAGAAAACTAT-3′,扩增片段大小约为715 bp,退火温度48℃。扩增eIF4E1H-T基因cDNA部分序列的引物对为CF4:5′-GAAAACACACGAAAATGGCAGAG-3′,CR4:5′-CTGCCGAGCCTCATTGAGTCGT-3′,扩增片段大小约为645 bp,退火温度48℃。扩增烟草内参基因Actin的特异引物为Actin-F:5′-AAGGGATGCGAGGATGGA-3′,Actin-R 5′-CAAGGAAATCACCGCTTTGG-3′,扩 增 片 段 大小为160 bp,退火温度58℃。PCR 反应体系总体积均为20 μL,其中30~50 ng/μL cDNA 样品2.5 μL、10×PCR buffer 2.0 μL, dNTPs 1.2 μL,引 物 各1.5 μL,rTaq DNA酶0.3 μL,ddH2O 12.6 μL。所用试剂购自宝生物公司。PCR反应在Ependorff梯度扩增仪上(Master Cycler)上进行。扩增的程序为:1)94℃预变性5 min;94℃变性30 s,退火温度30 s,72℃延伸1 min,共28个循环,72℃延伸10 min,4℃保存;采用2%的琼脂糖凝胶进行检测。
1.5 测序与序列分析
采用PCR产物直接测序或者克隆测序。每份野生种取3个单株叶片样品作为3个生物学重复样品,分别提取总RNA,随机引物反转录成cDNA并进行PCR扩增。每个样品PCR胶回收产物或者一个克隆双向测序。采用MEGA6比对序列,手工校正3次重复测序中个别差异碱基,确定3次测序结果一致的一条序列,代表该野生种的eIF4E1同源基因的cDNA序列。采用MEGA6软件UPGMA方法构建cDNA或推导氨基酸的系统进化树。
2 结果与分析
2.1 野生种eIF4E1-S同源基因扩增与PVY抗性
eIF4E1-S基因的特异引物CF3/CR3可特异扩增普通烟和N.sylvestris的eIF4E1-S基因cDNA全长。CF4/CR4引物可同时扩增出N.tomentosiformis eIF4E1H-T基因和N.sylvestris的eIF4E1-S基因cDNA部分序列。34个烟草野生种可扩增出目标条带如表1所示。其中19个野生种至少抗一个PVY分离物,11个野生种同时感供试的6个PVY分离物,4个野生种的PVY抗性不明。感PVY的野生种能扩增出eIF4E1-S基因,而抗PVY的野生种有的扩增出eIF4E1-S基因、有的扩增出其同源基因。
2.2 抗PVY野生种eIF4E1-S同源基因的多态性与含有抗PVY新基因的可能性
通过测序获得野生种eIF4E1-S同源基因的cDNA序列和推导的氨基酸序列,如果同一个野生种同时扩增出eIF4E1-S和eIF4E1H-T的同源基因,选用eIF4E1-S同源基因构建系统进化树。文献报道植物eIF4E蛋白表面帽结合袋(cap binding pocket)附近的2个结构域决定其对PVY属病毒的感病性[15]。普通烟草感PVY基因eIF4E1-S编码的loop1氨基酸序列为WFDNPMAKSRQAAWGSS(第53-69位氨基酸)、loop2为 INHPSKLVVG(第89-98位氨基酸)[14]。在MEGA6软件UPGMA方法构建的系统进化树上(图1),绒毛烟草组(Tomentosae)的4个抗PVY野 生 种N.otophora、N.setchelli、N.tomentosa、N.tomentosiformis的同源基因与eIF4E1H-T聚成一组。上述4个野生种同源基因编码的loop1与eIF4E1H-T的一致、loop2与eIF4E1-S和eIF4E1H-T仅存在V97A,M,N,T差异。推测这4个野生种的eIF4E1-S同源基因进化为eIF4E1H-T,获得对PVY部分株系的抗性。且这4个野生种都不抗可克服va抗性的PVY突变株系[13],因此,这4个野生种包含抗PVY新基因的可能性较小。圆锥烟草组(Paniculatae)的4个抗PVY野生种N.benavidesii、N.raimondii、N.knightiana聚成一组、N.cordifolia在较远分支,同源基因的氨基酸序列与eIF4E1-S一致率高于eIF4E1H-T。上述4个野生种loop1和loop2氨基酸序列相互之间差异较大,与eIF4E1-S和eIF4E1H-T都不相同,这4个野生种的PVY抗性既可能来源于eIF4E1-S基因突变又可能来源于抗PVY新基因。因此,这4个野生种包含抗PVY新基因的可能性中等。
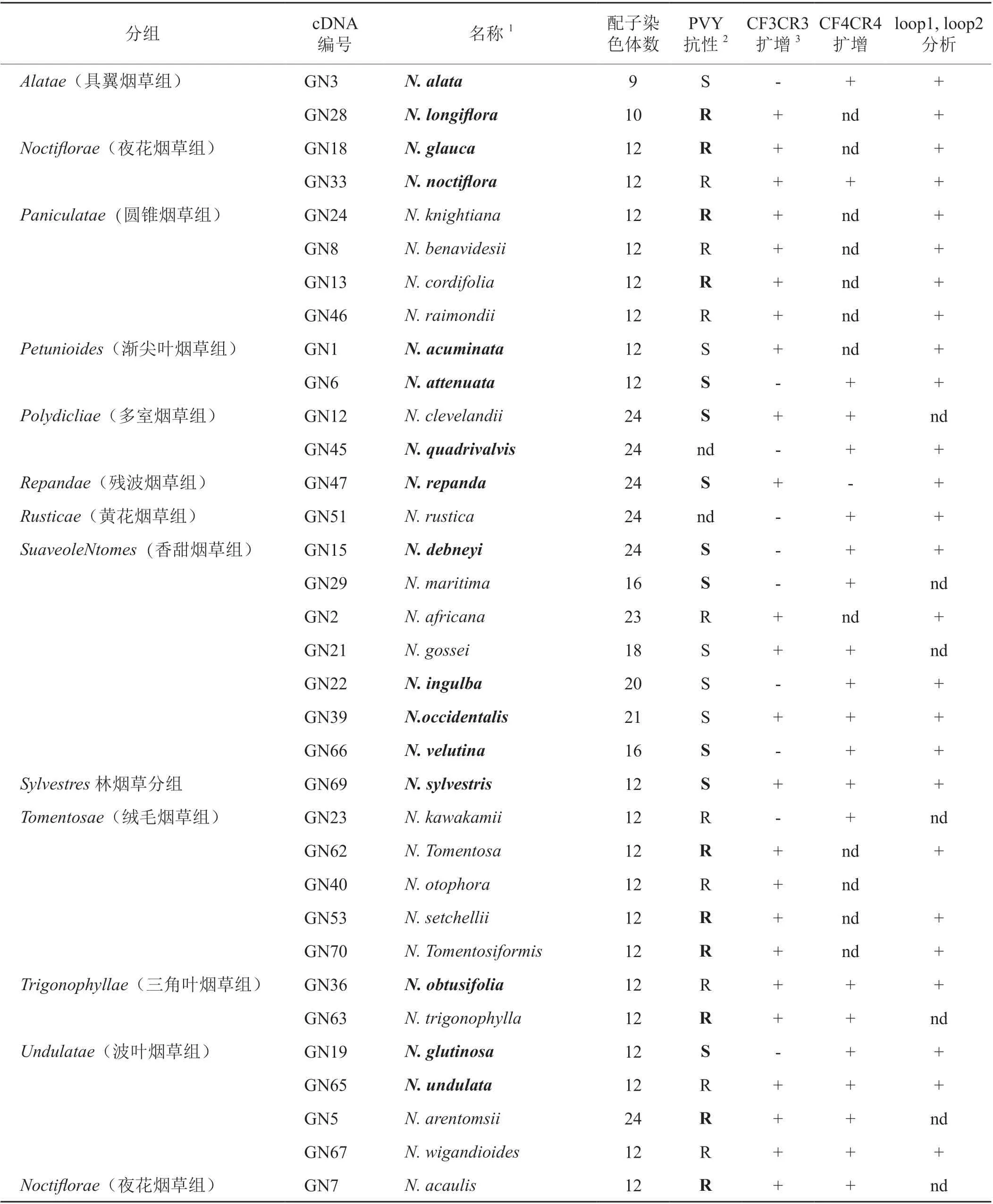
表1 烟草野生种的PVY抗性与eIF4E1-S同源基因的cDNA检测Tab. 1 PVY resistance of wild species and eIF4E1-S homologs cDNA detection
其他抗PVY和感PVY野生种聚为一个大的分支。该分支中与eIF4E1-S聚成一个小分支的N.glauca、N.noctiflore、N.africana高抗供试的5个PVY分离物(包含可克服va抗性的PVY分离物)。氨基酸序列比对结果表明N.glauca和N.noctiflore的同源基因与eIF4E1-S蛋白分别只有7个和4个氨基酸酸差异,其中loop1氨基酸序列与eIF4E1-S相同,loop2与eIF4E1-S之间只有1-2个氨基酸差异(N.glauca为K94E+V97M,N.noctiflore为V97M),N.glauca和N.noctiflore之间只有一个相同的差异位点(未列出数据)。推测N.glauca和N.noctiflore的PVY抗性与eIF4E1-S的氨基酸突变无关,存在新的抗PVY基因的可能性高。N.africana同源基因编码的loop1与eIF4E1-S和eIF4E1H-T差异较大、loop2与eIF4E1-S之间只有1个氨基酸差异(V97M)。N.africana存在抗PVY新基因的可能性稍低于N.glauca和N.noctiflore。Lewis等报道[16]N.africana存在与va不同的PVY抗性。N.longiflora的同源基因与eIF4E1-S聚成一个小分支,loop1氨基酸序列与eIF4E1-S相同,loop2只存在V97M差异,但只抗PVYNW(-)分离物。N.longiflora存在抗PVY新基因的可能性高,但育种价值较小。

图1 烟草野生种eIF4E1-S同源基因推导氨基酸序列构建的系统进化树(UPGMA)Fig.1 System Evolution tree (UPGMA) for eIF4E1-S homologs deduced amino acid sequences of tobacco wild species
2.3 感PVY野生种关键结构域高度保守
文献报道普通烟草感PVY蛋白eIF4E1-S的loop1和loop2与VPg诱饵蛋白eIF4E1H-T存在较大差异(图2)。本文11个感病野生种的loop1氨基酸序列与eIF4E1-S一致、且其中8个感病野生种的loop2只存在V97M/A差异,N.glutinosa的loop2 只存在G98N差异,N.repanda的loop2只存在H89L差异。表明感PVY野生种eIF4E-S蛋白关键结构域高度保守。

图2 抗感PVY野生种的loop1和loop2氨基酸比较Fig.2 Comparison of loop1 and loop2 amino acids of wild species with PVY resistance
3 讨论
3.1 烟草野生种PVY抗性与eIF4E1-S基因的关系
烟草育种长期利用eIF4E1-S缺失的抗源(va),田间已出现可克服va抗性的突变病毒[14,17-19],表明va品种的抗性持久性不够高;eIF4E1-S基因移码突变体或无义突变体的抗性持久性低于va,不能满足抗病育种需要。因此,有必要在野生种中发掘不同于eIF4E1-S基因缺失或者突变的新抗病基因。鉴定筛选抗PVY的野生种相对容易,采用常规的摩擦接种抗性鉴定即可。但分析抗PVY野生种是否存在新抗病基因难度很大,因为大部分野生种与普通烟草、野生种之间存在生殖隔离,几乎不可能利用常规杂交方法分析抗病基因是否等位。野生种PVY抗性包括三种可能:感病基因缺失/突变、存在新抗病基因、二者共存。本文尝试利用同源基因克隆测序方法预测野生种存在新抗病基因的可能性,并提出如下假说:在存在eIF4E1-S感病基因的情况下,抗PVY野生种携带抗PVY新基因的可能性高;在不存在eIF4E1-S感病基因(包括存在eIF4E1H-T)的情况下,抗PVY野生种携带抗PVY新基因的可能性较低。
几篇文献报道感PVY属病毒的eIF4E蛋白表面存在loop1和loop2关键氨基酸结构域[14-15,21-22],如:抗PVY辣椒种质资源的2个loop存在非同义突变、这些氨基酸突变与抗性有关[23]。感病辣椒的这两个结构域的单个或几个关键氨基酸突变可获得PVY抗性。通过比较野生种loop1和loop2氨基酸序列来判断野生种是否含有感病基因。本文再进一步推测:抗PVY野生种的loop1和loop2氨基酸序列与eIF4E1-S基因相同,则该野生种含有抗PVY新基因的可能性高;抗PVY野生种的loop1和loop2氨基酸序列与eIF4E1-S基因不同(包括与eIF4E1H-T相同),则该野生种含有抗PVY新基因的可能性较低。在排除个别野生种eIF4E1-S同源基因缺失/突变、且含有抗PVY新基因的情况下,某个野生种loop1和loop2氨基酸序列与eIF4E1-S的多样性越高,则越可能丧失PVY感病性,含有新抗病基因的可能性较低。
3.2 野生种eIF4E1-S同源基因多样性与抗PVY新基因可能性
本文分析了34个抗/感PVY野生种的eIF4E1-S同源基因的推导氨基酸序列,表明野生种eIF4E1-S同源基因之间存在较大的多样性,感PVY属病毒的eIF4E1蛋白关键结构域loop1和loop2的氨基酸序列高度保守。其中10个感PVY野生种loop1氨基酸序列与eIF4E1-S一致、其中8个野生种loop2仅在V97M, A存在差异。表明感PVY野生种loop1和loop2的保守性显著高于抗PVY野生种。根据以上假说,本文预测了野生种中存在抗PVY新基因的可能性高低。绒毛烟草组(Tomentosae)的4个 野 生 种N.otophora、N.setchelli、N.tomentosa、N.tomentosiformis的PVY抗性可能与eIF4E1进化为eIF4E1H-T有关,其育种利用价值不大。因为感PVY的普通烟中存在eIF4E1H-T。圆锥烟草组(Paniculatae)的4个抗PVY野 生 种N.benavidesii、N.raimondii、N.cordifolia、N.knightiana推导的氨基酸序列与eIF4E1-S一致率高于eIF4E1H-T,loop1 和loop2与eIF4E1-S存在较为随机的2-6个氨基酸差异,这些氨基酸变异可能导致与PVY VPg不能互作,获得PVY抗性。因此,圆锥烟草组4个抗PVY野生种包含抗PVY新基因的可能性中等。
根据loop1 和loop2的变异较小且抗多个PVY分离物,推测N.glauca、N.noctiflore存在新的高抗PVY基因的可能性高、N.africana其次。N.africana作为父本可与普通烟杂交结实,产生母本来源的单倍体和混倍体,其PVY抗性已经部分转育至普通烟中[16],可增强va抗性,对可克服va抗性的PVY突变株系也有一定的抗性。尚未见N.glauca、N.noctiflore的PVY抗性转育至普通烟草中的报道,发掘利用烟草野生种的抗PVY新基因推荐以N.glauca、N.noctiflore、N.africana为重点。
PVY抗性不明确的N.quadrivalvis的loop1和loop2氨基酸序列与eIF4E1-S一致,推测其不是感PVY就是存在新抗病基因。PVY抗性不明确的N.rustica,N.wigandioides的loop1与eIF4E1H-T一致率较高,与eIF4E1H-T存在5个氨基酸差异,如果抗PVY,则其抗性可能与同源基因进化为eIF4E1H-T有关,因此,含有抗PVY新基因的可能性中等。
3.3 感PVY同源基因克隆测序法的可行性与局限性
烟草属野生种大部分为二倍体,少数为异源四倍体。高质量的全基因组数据或者全长转录组数据,可从全局上分析野生种eIF4E1-S同源基因。但因为烟草基因组巨大和重复序列多,导致近期内难以获得30多份野生种的基因组数据或全长转录组数据。本文尝试采用同源基因克隆测序方法预测野生种存在抗PVY新基因的可能性,为目前可行的尝试。我们根据普通烟草的eIF4E1-S、eIF4E1H-T和祖先种同源基因设计引物扩增测序,本文获得了34个野生种eIF4E1同源基因的cDNA序列。其他3个野生种(N.auculis和N.sanderae,N. petuniodes)供试引物扩增不成功、7个野生种的个别氨基酸存在疑问,未采用同源基因分析方法预测抗PVY野生种是否存在新的抗PVY基因。eIF4E1-S同源基因克隆测序法的局限性在于,野生种同源基因的变异较大,难以设计可扩增所有野生种的通用引物。四倍体野生种的同源基因可能存在两个拷贝,导致克隆测序时个别氨基酸序列不一致。
4 结论
根据eIF4E1-S同源基因的多样性,将抗PVY烟草野生种存在抗PVY新基因(即不属于eIF4E1-S基因缺失或突变)的可能性划分为高、中和低三档。N.glauca、N.noctiflore、N.africana含有新的抗PVY基因可能性高于其他供试野生种,可作为重点抗源研究利用。
