翠云草叶绿素代谢途径及其关键酶
2020-11-19杨露露李林
杨露露 李林


摘 要 通过比较蓝色叶翠云草、红色叶翠云草和绿色叶小翠云三者叶绿素合成途径中11种产物的含量变化,分析翠云草叶绿素代谢途径及其关键酶,为深入研究翠云草叶色变化内在机制奠定基础。结果显示,“原卟IX→镁原卟啉IX”“原叶绿酸酯→叶绿酸酯a”这两个环节是翠云草叶绿素代谢途径的关键环节,镁离子螯合酶(CHLH)、光依赖的原叶绿酸酯氧化还原酶(LPOR)是相应的两个关键酶。基于对翠云草叶绿素代谢途径的分析,推测翠云草中叶绿素a转化为叶绿素b的效率大于其反向转化的效率,导致叶绿素a/b值小于2,以适应翠云草生长的荫蔽环境,满足其叶绿素需求。
关键词 翠云草;叶绿素;代谢途径;关键酶
中图分类号:S68 文献标志码:A DOI:10.19415/j.cnki.1673-890x.2020.25.001
彩叶植物尤其是蓝色叶植物在园林景观应用中较为少见。卷柏科(Selaginellaceae)卷柏属(Selaginella)的翠云草(Selaginella uncinata)是一种蓝色叶的蕨类植物,极具观赏价值[1]。其叶片如羽毛般柔软,在荫蔽条件下呈现蓝色,在全光照下变为暗红色,叶色变化呈现的景观效果很独特,具有广阔的园林应用前景。
迄今为止,国内外对翠云草的研究主要集中在基因组学、栽培繁殖、化工及药理学方面。Tsuji等[2]、Smith[3]、Oldenkott等[4]对翠云草进行了基因组学方面的研究。H Bant等研究了卷柏属的藤卷柏(S. willdenovii)和翠云草,利用透射电子显微镜观察其叶片横切面的组织构造,认为翠云草的蓝色叶与其叶片上层表皮的外侧细胞壁内两层薄膜干涉滤器引发光的薄膜干涉有关[5]。郑俊霞等[6]、邱宏聪等[7]、赖红芳等[8]对翠云草的化学组成、提取工艺、药理作用等进行了研究。黄莉洁[9]、张旭乐等[10]、杨俊杰[11]总结了翠云草的引种人工栽培和繁殖技术。李林等[12]、LI L等[13]、张水木等[14]从翠云草叶色变化及其分子机理等方面探讨了翠云草呈色变化机制。
本研究通过比较三种植物材料(蓝色叶翠云草、红色叶翠云草和绿色叶小翠云)叶绿素合成途径中11种中间产物含量的变化,探讨翠云草叶绿素代谢途径的特点及关键酶,为深入研究翠云草叶色变化的内在机制,最终实现人工调控叶色奠定基础。
1 材料与方法
1.1 試验材料
试验基地位于广西大学林学院苗圃(22°50′N,108°17′E)。盆栽(株龄均为6个月)翠云草和同属的小翠云(S. kraussiana)购自广西花鸟市场,参照李林等[12]的方法对所用植物材料进行预处理,得到全光照下栽培的红色叶翠云草(图1B),用于和放在三层遮荫条件下栽培的蓝色叶翠云草(图1A)和绿色叶小翠云(图1C)进行对比试验。
1.2 试验方法
随机采集植株各个方向正常生长的成熟叶片,测定叶绿素生物合成各产物的含量。
1.2.1 叶绿素合成前体物质的测定
利用全自动波长酶标仪测定叶绿素合成前体物质的吸光值,其中δ-氨基酮戊酸(ALA)、胆色素原(PBG)、尿卟啉原Ⅲ(Urogen Ⅲ)、粪卟啉Ⅲ(Coprogen Ⅲ)、原卟啉IX(Proto IX)、镁原卟啉IX(Mg-Proto IX)和原叶绿酸酯(Pchlide)含量的测定参照宗学凤[15]、赵明辉等[16]、王平荣等[17]、杨海芸[18]的方法,并适当调整,单位均采用nmol·g-1。
1.2.2 叶绿素及其衍生物含量的测定
利用高效液相色谱法(WATERS E2695 HPLC系统)对叶绿素及其衍生物含量进行测定,根据标准曲线,利用保留时间和峰面积确定样品中的物质种类和含量。标准品叶绿素a(Chl a)和叶绿素b(Chl b)购于Sigama公司。标准品叶绿酸酯a(Chlide a)和叶绿酸酯b(Chlide b)利用翠云草叶片中叶绿素酶(Chlase,参照Costa[19]的方法提取)分别以标准品Chl a和Chl b为底物发生反应得到,所得的Chlide a和Chlide b仅可进行定性,无法测定标准样中Chlide a和Chlide b的具体含量,因此在结果分析中以样品中检测出的峰面积相对比较三种植物材料叶片中Chlide a和Chlide b含量情况。
叶绿素及其衍生物的HPLC分析参照程红艳等[20]的方法,略加改进。取0.15 g植物叶片迅速加入5 mL甲醇溶液充分浸泡,并用超声波机器震荡15 min至无色,过滤待测。检测条件为柱型C18(4.6 mm×250 mm,德国Nucleodur色谱柱),检测波长430 nm,流动相为A(甲醇、水的体积比为80∶20)和B(乙酸乙酯)。梯度洗脱为:0~20 min,A 100%~50%,B 0%~50%;20~40 min,A=50%,B=50%。流速设置为0.8 mL·min-1,进样量为10 μL。
1.3 数据整理
利用Word 2010、Excel 2010进行数据处理、制图、制表,利用SPSS 20.0进行方差分析和相关性分析。
2 结果与分析
2.1 叶绿素合成前体物质含量分析
从表1可以看出,除Coprogen Ⅲ外,其余各产物含量在三种叶色中都具有显著性差异,但其大小的变化趋势在三种叶色中不一致。
2.2 叶绿素及其衍生物分析
2.2.1 叶绿素及其衍生物HPLC图谱
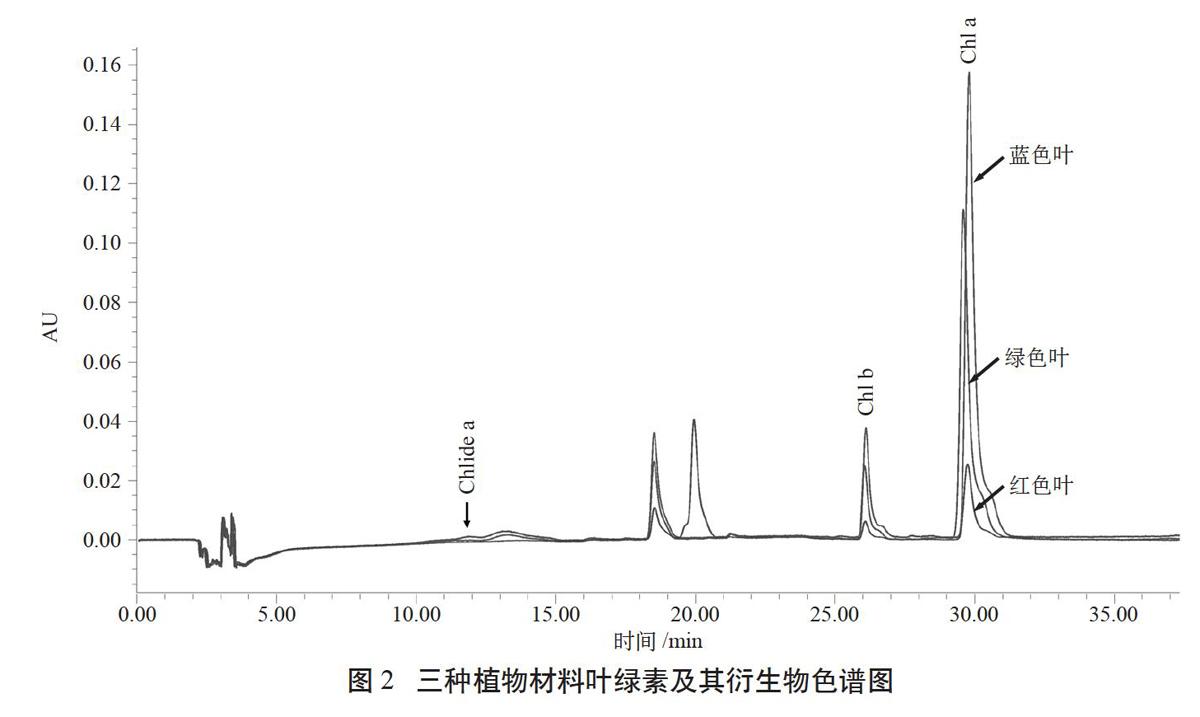
图2为三种植物材料在430 nm处的叶绿素及其衍生物HPLC色谱图。各主要物质的出峰时间分别为:Chl a,29.846 min;Chl b,26.169 min;Chlide a,11.788 min。除Chlide b以外,另外3种物质在三种植物材料中均可以检测到。此外,三种叶色都在17.669 min处检测到脱镁叶绿素a(Pheide a)。
2.2.2 叶绿素及其衍生物含量分析
表2显示,除叶绿素a/b外,三种植物材料中Chl a、Chl b、Chl a+b和Chlide a都存在显著差异。Chl a、Chl b、Chl a+b的大小变化趋势相同,都是蓝色叶>绿色叶>红色叶,且3种叶色中都是Chl a>Chl b。Chlide a的大小变化则是绿色叶>蓝色叶>红色叶。叶绿素a/b值均小于2,大小依次为红色叶>绿色叶>蓝色叶。
2.3 叶绿素合成产物相对含量比较
为了便于更直观地分析和描述,将蓝色叶中各物质含量设定为100,与红色叶和绿色叶进行比较,且分别将叶绿素合成过程ALA→PBG→Urogen III→Coprogen Ⅲ→Proto IX→Mg-Proto IX→Pchlide→Chlide a→Chl a→Chl b命名为阶段①~阶段⑨,结果见图3(参考杨露露[21]并做了一些改动)。
1)在阶段①,红色叶中合成的前体物质由原来蓝色叶的135.25%下降到89.09%,紧接著在阶段②又上升到148.99%,在阶段③和阶段④中,红色叶>蓝色叶的趋势不变,但到了阶段⑤,红色叶中的合成产物急剧下降为蓝色叶的32.39%,其后一直保持低于蓝色叶的趋势。根据以上蓝色叶和红色叶的比较分析,推测阶段①ALA→PBG、阶段②PBG→Urogen III和阶段⑤Proto IX→Mg-Proto IX是翠云草叶绿素合成的关键环节。
2)在阶段③,绿色叶中合成的前体物质由原来蓝色叶的37.46%上升到100.76%,紧接着在阶段④又下降到蓝色叶的70.32%,在阶段⑤和阶段⑥中,蓝色叶>绿色叶的趋势不变,但到了阶段⑦,绿色叶中的合成产物快速上升为蓝色叶的128.34%,随后阶段⑧又急剧下降为71.06%,其后保持低于蓝色叶的趋势。根据以上蓝色叶和绿色叶的比较分析,推测阶段③Urogen III→Coprogen Ⅲ、阶段④Coprogen Ⅲ→Proto IX、阶段⑦Pchlide→Chlide a和阶段⑧Chlide a→Chl a是翠云草叶绿素合成的关键环节。
2.4 翠云草叶绿素合成产物相关性分析
将翠云草叶绿素合成途径各中间产物:ALA、PBG、Urogen III、Coprogen III、Proto IX、Mg-Proto IX、Pchlide、chlide a的含量分别与叶绿素指标进行相关性分析,结果(见表3)显示,前4个中间产物ALA、PBG、Urogen III、Coprogen III的含量与叶绿素各指标均无显著相关关系,而后4个中间产物Proto IX、Mg-Proto IX、Pchlide、Chlide a的含量与叶绿素各指标(除Chl a/b外)显著相关。
3 讨论与结论
3.1 翠云草的耐阴性
植物在不同光照强度的生长环境中,为满足体内叶绿素性能的需求,可以利用叶绿素循环,自我调整Chl a和Chl b的比值[22]。一般说来,叶绿素含量高、Chl a/b比值小的植物具有较强的耐阴性。低的Chl a/b值能提高植物对远红光的吸收,因而在弱光下,具有较低的Chl a/b值及较高的叶绿素含量的植物,也具有较高的光合活性。蕨类作为典型的阴生植物,Chl a/b值通常小于3∶1[23]。据丁晓浩等报道,其所测的10种观赏蕨在3种不同光照处理下的Chl a/b值均小于3∶1,为(0.17~2.49)∶1[23]。
本研究中三种植物材料Chl a/b值均小于2,和丁晓浩等的研究结果[23]一致。其比值大小依次为红色叶翠云草>绿色叶小翠云>蓝色叶翠云草,说明翠云草比同属的小翠云耐阴,而红色叶翠云草是受到强光胁迫形成的[12],其耐阴性最差。
3.2 翠云草叶绿素合成关键环节和关键酶
根据前文(“2.3”节)蓝色叶和红色叶、绿色叶的比较分析,推测翠云草叶绿素合成的关键环节是阶段①ALA→PBG、阶段②PBG→Urogen III、阶段③Urogen III→Coprogen Ⅲ、阶段④Coprogen Ⅲ→Proto IX、阶段⑤Proto IX→Mg-Proto IX、阶段⑦Pchlide→Chlide a和阶段⑧Chlide a→Chl a这7个环节。
通过“2.4”节相关性分析可知,Proto IX、Mg-Proto IX、Pchlide 、Chlide a这4个产物与叶绿素指标显著相关。即图3中的阶段④→阶段⑦环节与叶绿素合成的关系更为密切。
Li等根据转录组数据,对翠云草叶绿素代谢相关基因进行差异表达分析,发现镁离子螯合酶(CHLH)、光依赖的原叶绿酸酯氧化还原酶(LPOR)、叶绿素酶(CLH)(即图3中的阶段⑤、阶段⑦和阶段⑨所对应的3个酶)在蓝色叶中比红色叶中均有所上调[13]。
综上,我们认为阶段⑤Proto IX→Mg-Proto IX和阶段⑦Pchlide→Chlide a这两个环节是翠云草叶绿素代谢途径的关键环节,CHLH和LPOR是相应的两个关键酶。
3.3 翠云草叶绿素代谢途径
杨海芸对花叶矢竹(Pseudosasa japonica f. akebonosuji)进行叶色突变研究,发现引起其叶色变异的原因在于:Coprogen III→Proto IX这一中间过程(图3中的阶段④)生成的Proto IX与上一产物Coprogen III相比较,其相对含量水平急剧下降,导致花叶矢竹白色条纹叶Chl a、Chl b和总叶绿素的含量均低于绿色叶,致使叶片颜色变淡或白化[18]。孙捷音等发现油菜(Brassica napus)的叶绿素合成途径中,从PBG→Urogen III(图3中的阶段②),和野生型相比,其含量降低了一半,是Chl b的缺失造成了叶色突变[24]。董立花采用荧光光谱和紫外可见光谱分析了金心吊兰(Chlorophytum capense var. Medio-pictum)叶绿素代谢流中间产物的变化,发现其金心部分ALA、Proto IX和Pchlide的合成(图3中的起始环节和阶段④、阶段⑥)受阻,且推测ALA合成受阻可能是其形成黄色叶条纹的主要原因[25]。上述各例都是由于叶绿素合成过程中Chl a、Chl b和总叶绿素产生变化后导致叶色发生改变。Li等通过对蓝色叶翠云草和红色叶翠云草进行转录组测序,找到了翠云草叶绿素代谢途径中几乎所有的酶,且基于两个样本中的叶绿素合成酶(Chlorophyllsynthase, CS)和叶绿素a加氧酶(CAO)的表达都没有显著差异的结果,推断翠云草中两种叶绿素的合成都没有受阻[13]。
从L-谷氨酰-tRNA到Chl a,再到Chl b,整个叶绿素生物合成过程一共15步反应,需要15种酶参与[17]。叶绿素循环过程从Chlide a开始,经过CS酯化反应形成Chl a。Chl b是由Chl a转化形成的,在此过程中,Chlide a在CAO的催化下形成Chlide b,Chlide b在CS作用下形成Chl b。而Chl b在一定的条件下也可还原为Chl a。植物可以利用Chl a和b能够相互转化的反应来调整叶绿素a/b的比值以适应不同的生理条件和环境变化[26]。通常Chl a和Chl b吸收不同的波长,前者吸收红光而后者主要吸收蓝紫光,在荫蔽条件下,Chl b能够利用环境中的蓝紫漫反射光适应光照条件。本试验应用HPLC在三种叶色材料中检测到Chl a、Chl b和Chlide a,但均未检测到Chlide b,推测Chlide a在CAO催化下,产生的Chlide b迅速进入下一步反应生成Chl b,使得Chlide b在代谢流中不积累,即翠云草中Chl a转化为Chl b的效率大于其反向转化的效率,导致叶绿素a/b值小于2,以适应翠云草生长的荫蔽环境,满足其叶绿素需求。
图4为我们参考Li等[13]并做了一些改动,总结出的翠云草叶绿素代谢通路图。
参考文献:
[1] 吉醒.苍翠清新的翠云草[J].花木盆景(花卉园艺),2011(10):10-11.
[2] Tsuji S, Ueda K, Nishiyama T, et al. The chloroplast genome from a lycophyte (microphyllophyte), Selaginella uncinata, has a unique inversion, transpositions and many gene losses[J]. Journal of Plant Research, 2007, 120(2): 281-290.
[3] Smith D R. Unparalleled GC content in the plastid DNA of Selaginella[J]. Plant Molecular Biology, 2009, 71(6): 627-639.
[4] Oldenkott B, Yamaguchi K, Tsuji-tsukinoki S, et al. Chloroplast RNA editing going extreme: more than 3400 events of C-to-U editing in the chloroplast transcriptome of the lycophyte Selaginella uncinata[J]. RNA, 2014, 20(10): 1499-1506.
[5] H Bant C, LEE D W. Ultrastructural Basis and Developmental Control of Blue Iridescence in Selaginella Leaves[J]. American Journal of Botany, 1984, 71(2): 216-219.
[6] 邱宏聪,陈小刚.壮药材翠云草化学成分、药理活性及质量控制的研究进展[A].中华中医药学会.中华中医药学会第七次中药分析学术交流会会议论文集[C].广州,2014.
[7] 郑俊霞,郑扬,颜秋萍,等.翠云草总黄酮的提取工艺研究[J].中药材,2011(12):133-135.
[8] 赖红芳,温晓娟.果胶酶法提取翠云草中总黄酮的工艺优选[J].中国实验方剂学杂志,2012,18(23):34-36.
[9] 黄莉洁.观赏蕨类卷柏——翠云草的引种栽培和繁殖技术[J].农业研究与应用,2011(5):86-88.
[10] 张旭乐,杨燕萍,黄建,等.蕨类植物翠云草栽培繁殖技术及园林应用价值[J].农业科技通讯,2009(6):202-203.
[11] 杨俊杰.翠云草栽培管理技术[J].农业工程技术·温室园艺,2015(4):46-48.
[12] 李林,黄洁瑜,张水木,等.不同遮荫处理对翠云草叶色变化的影响[J].分子植物育种,2015(5):193-198.
[13] LI L, WANG Q, DENG R, et al. Transcriptome profiling of blue leaf coloration in Selaginella uncinata[J]. Canadian Journal of Plant Science, 2016, 97(4): 580-593.
[14] 张水木,彭媛媛,李林.不同光质处理对翠云草叶色变化的影响[J].北方园艺,2016(12):75-79.
[15] 宗学凤.水稻叶片全生育期黄化突变体ylg3叶绿素合成特性研究[J].西南大学学报(自然科学版),2013,35(11):21-26.
[16] 赵明辉,隋阳辉,陈温福.超绿水稻叶绿素的生物合成[J].作物杂志,2010(6):33-37.
[17] 王平榮,张帆涛,高家旭,等.高等植物叶绿素生物合成的研究进展[J].西北植物学报,2009(3):207-214.
[18] 杨海芸.花叶矢竹叶色变异机理研究[D].北京:北京林业大学,2015.
[19] COSTA M L, CIVELLO P M, CHAVES A R, et al. Effect of ethephon and 6-benzylaminopurine on chlorophyll degrading enzymes and a peroxidase-linked chlorophyll bleaching during post-harvest senescence of broccoli (Brassica oleracea L.) at 20℃[J]. 2005, 35(2): 191-199.
[20] 程红艳,陈军辉,张道来,等.超声波辅助提取RP-HPLC法测定浒苔中的叶绿素a、b[J].海洋科学,2010,34(2):23-27.
[21] 杨露露.观赏蕨翠云草叶绿素代谢途径研究[D].南宁:广西大学,2019.
[22] 李祖祥,吕梅,夏爱军.遮阴对冷季型草坪草叶绿素含量和生物量的影响[J].草原与草坪,2017,37(5):69-74.
[23] 丁晓浩,何云核.10种观赏蕨类植物的耐阴性[J].江苏农业科学,2013,41(6):160-162.
[24] 孙捷音,张年辉,杜林方,等.油菜叶绿素b减少突变体Cr3529叶绿素生物合成的研究[J].西北植物学报,2007,27(10):1962-1966.
[25] 董立花.金心吊兰的叶绿素生物合成以及光胁迫的耐受性[D].雅安:四川农业大学,2015.
[26] 陈熙.低叶绿素b水稻类囊体膜的蛋白质组学研究[D].南京:南京农业大学,2009.
(责任编辑:丁志祥)
