苦荞株型相关性状的遗传分析及其对产量的影响
2020-11-18李春花尹桂芳黄金亮王艳青卢文洁孙道旺王春龙郭来春魏黎明加央多拉任长忠王莉花
李春花 ,尹桂芳,黄金亮,王艳青,卢文洁,孙道旺,王春龙,郭来春,魏黎明,加央多拉,任长忠*,王莉花*
(1.云南省农业科学院生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业部西南作物基因资源与种质创制重点实验室,云南昆明 650205;2.吉林省白城市农业科学院,吉林白城 137000;3.吉林省敦化市马铃薯开发繁育中心,吉林敦化 133700)
【研究意义】苦荞(Fagopyrum tataricumGaertn.)属于蓼科(Polygonaceae)荞麦属(Fagopyrum Mill)的一年生草本植物[1]。苦荞的芦丁含量较高,芦丁是一种苷糖化合物,具有消积化滞、除湿解毒等作用,对高血压及毛细管脆弱而引起的脑出血,对糖尿病等有一定的医疗作用[2]。近年,由于苦荞的营养特性和机能特性方面的优越性,苦荞以健康食品而备受关注,因此对苦荞需要量也增加[2]。但是荞麦的低产性严重阻碍了荞麦生产,加上高产粮食作物的冲击,致使世界各国的荞麦生产呈现下降趋势[3]。作物株型不仅直接影响叶片的有效光合面积,而且影响冠层结构内部水、气、温等环境,最终影响群体的光合效率和作物产量[4]。Donald[5]在1968 年提出理想株型,即有利于植物的生长发育、光合作用和籽粒产量的最理想化的株型,能够最大限度地提高光合利用率。在高产品种育种过程中改良株型的目的是为了尽可能地提高群体光合效率和物质生产能力,使群体在较高的光合效率和籽粒生产水平上达到动态平衡,最终获得较高的产量[6-7]。因此,开展株型相关性状的遗传研究对改善植株的形态特征,培育高产优质苦荞品种选育具有重要的理论意义和实际参考价值。【前人研究进展】众多研究者在水稻[8]、大豆[9]、绿豆[10]、高粱[11]、玉米[12]、甘蔗[13]等作物的株型相关性状的遗传进行了广泛研究。近年在苦荞上也有一些株型相关性状的研究,如Li 等[14]利用苦荞品种‘北陆4 号’和‘石荞麦’的杂交分离后代F2和F3进行了开花期、株高和单株粒质量的遗传分析,结果表明3 个性状的显性方差都大于加性方差,且株高与单株粒质量具有较大的相关性;梁龙兵[15]利用小米荞和晋荞2 号的杂交后代F2、F3、F5进行了遗传分析,结果表现株高和主分支数与单株粒数呈显著正相关;陈庆富等[16]利用小米荞和米荞1 号为母本,晋荞麦2 号、黔苦5 号进行杂交获得的F2群体进行了遗传分析,结果表明株高与株粒数和株粒质量呈显著的正相关。但在苦荞株型相关性状的重要组成因素的株高、主茎节数和分枝数的遗传效应及其与产量相关性状之间的表现型相关和遗传相关性的研究则较少。
【本研究切入点】株型相关性状属于复杂的数量性状,一般受多数遗传基因的控制也容易受环境条件的影响,因此很难以表现型来判断其遗传基因型。松尾[17]指出,遗传率的高低与选拔的难易有关,并且决定选拔方法和育种方式上也起到很重要的作用。在实际育种过程中目标性状不止一个,而需要多个性状同时选拔,所以只了解每个性状的遗传特性信息是难以达到理想效果。因此有必要考虑这些性状的相互关联性的基础上选拔优良单株。然而,在选拔过程中表现两个性状间的基因连锁及基因多面表达的遗传相关性的信息是很重要的。【拟解决的关键问题】本研究以苦荞主要株型性状为研究对象,旨在通过对苦荞株高、主茎节数和分枝数进行遗传规律分析及其性状间以及与单株产量间的相关性分析,以期进一步弄清苦荞株型的遗传机制,从而为苦荞高产新品种培育提供理论参考。
1 材料和方法
1.1 试验材料
生育期、株高、单株粒质量等性状有较大差异的‘云荞1 号’[19]和‘米荞’[20]分别作为母本材料和父本材料。
1.2 试验方法
试验在云南省昆明安宁市县街镇甸心村的荞麦试验基地内进行,该地区海拔为1 887 m,纬度24°45′,经度102°25′。2013 年9 月以‘云荞1 号’和‘米荞’进行杂交获得F1代。杂交是借助便携式放大镜,用镊子对开花前的母本花蕾进行去雄操作,并取父本的花药对去雄的母本花蕾进行授粉之后套袋。杂交成功与否是利用SSR分子标记(BM27)对F1植株进行判断。母本‘云荞1号’没有条带,父本‘米荞’有条带,其F1为有条带。2014年3月将F1自交获得232粒种子,并同年7月与亲本各50粒进行播种。2015年7月由232个株系组成的F3群体中,取出每株系各50粒和亲本各50粒,株间距为10 cm进行点播种植。
1.3 测定项目
2014 年调查亲本各10 个单株和F2群体232 单株。2015 年调查亲本及F3群体232 个株系各10 个单株。两年都是待籽粒70%~80%成熟时收获,并调查株高、主茎节数、分枝数和单株粒质量。
1.4 数据分析
1.4.1 亲本及F2、F3群体各农艺性状的表现值分析 田间调查的基本数据用Excel 2007 软件进行整理,再使用JMP 9.0.2统计分析软件进行农艺性状的方差分析、相关分析和变异分析等。
1.4.2 农艺性状的遗传规律测定 用各农艺性状的后代间方差进行遗传规律和遗传率的测定[18]。F2群体和F3群体的方差可以用加性方差(A)、显性方差(B)和环境方差(E1)来表示。E1是计算式(1)来算出[21]。

从式(2)和(3)求出A和D,再利用A和D,根据于博[9]的计算方式算出广义遗传率和狭义遗传率。另外,利用计算式(6)来算出固定遗传率[18]。

1.4.3 性状间相关性测定 用F3群体的农艺性状表现值进行皮尔逊的积差相关分析,将性状A 和B 的表现型值设为YA,YB,利用计算式(7)算出表现型相关系数。

另外,把各亲本的单株间方差・共方差的平均分别称为环境方差・共方差,根据计算式(8)来算出遗传相关系数[18]。

eA和eB分别是性状A和B的环境效应,Var(YA)、Var(YB)和Cov(YA,YB)分别是F3群体的性状A和B的单株间方差以及A、B之间的单株间共方差。
Var(eA)=(‘云荞1号’性状A的单株间方差+‘米荞’性状A的单株间方差)/2
Var(eB)=(‘云荞1号’性状B的单株间方差+‘米荞’形状B的单株间方差)/2
Cov(YA,YB)=(‘云荞1 号’性状A 和B 之间的单株间共方差+‘米荞’性状A 和B 之间的单株间共方差)/2。用t检验来判断表现型相关系数的显著性。
2 结果与分析
2.1 亲本及F2、F3群体各农艺性状的表现值
由表1 可见,在株高上2014 年和2015 年母本的变异范围和平均值都小于父本,且2014 年的表现值小于2015年的表现值。在分枝数和主茎节数上父母本以及年次的差异不大,而在单株粒质量上2014年和2015 年母本的表现值大于父本的表现值,且2015 的表现值大于2014 年的表现值。变异系数来看,两年的株高、分枝数、主茎节数和单株粒质量由小到大依次为株高、主茎节数、分枝数和单株粒质量,并且都小于10%。在株高、分枝数、主茎节数和单株粒质量的F2群体和F3群体表现值范围都超过父母本的表现值,并且分枝数、主茎节数和单株粒质量的2015 年种植的F3群体表现值的平均值都大于2014 年种植的F2群体的表现值,而株高是2014年的表现值大于2015年的表现值。F2群体和F3群体的各农艺性状的变异系数来看,株高分别为20.5%和22.2%,分枝数分别为9.7%和8.2%,主茎节数分别为12.8%和10.1%,单株粒质量分别为51.0%和43.5%。其中分枝数的变异系数最小,且小于10%,单株粒质量的变异系数最大。

表1 亲本及F2和F3群体各农艺性状的表现值Tab.1 Performance values of each agronomic trait of parents and F2 and F3 groups
2.2 各农艺性状的遗传规律分析
由表2 可知,除了分枝数的加性方差大于显性方差,平均显性度小于1 以外,株高、主茎节数和单株粒质量的显性方差大于加性方差,平均显性度大于1。在广义遗传率来看,株高、分枝数、主茎节数和单株粒质量在F2代和F3代中差异并不大,且在0.62~0.78。在狭义遗传率来看,在F2代和F3代中单株粒质量分别为0.26 和0.31,最小,分枝数分别为0.65 和0.68 最大,且株高和主茎节数在其之间。另外,株高、分枝数、主茎节数和单株粒质量的固定遗传率分别为0.62、0.78、0.56和0.62。
2.3 各农艺性状间的表现型相关及遗传相关
由表3可知,在4个农艺性状间的表现型相关系数来看,单株粒质量与株高和主茎节数在1%水平上存在显著性正相关,与分枝数在5%水平上存在显著的正相关;株高与分枝数和主茎节数在1%水平长存在显著的正相关;分枝数与主茎节数在1%水平上存在显著的正相关。在4个农艺性状间的遗传相关系数来看,单株粒质量与株高、分枝数、主茎节数都存在正相关,且单株粒质量与株高的相关系数为0.592最大。另外,株高与主茎节数也存在较大的遗传相关性。

表2 各农艺性状的遗传规律分析表Tab.2 Genetic parameters of four agronomic traits
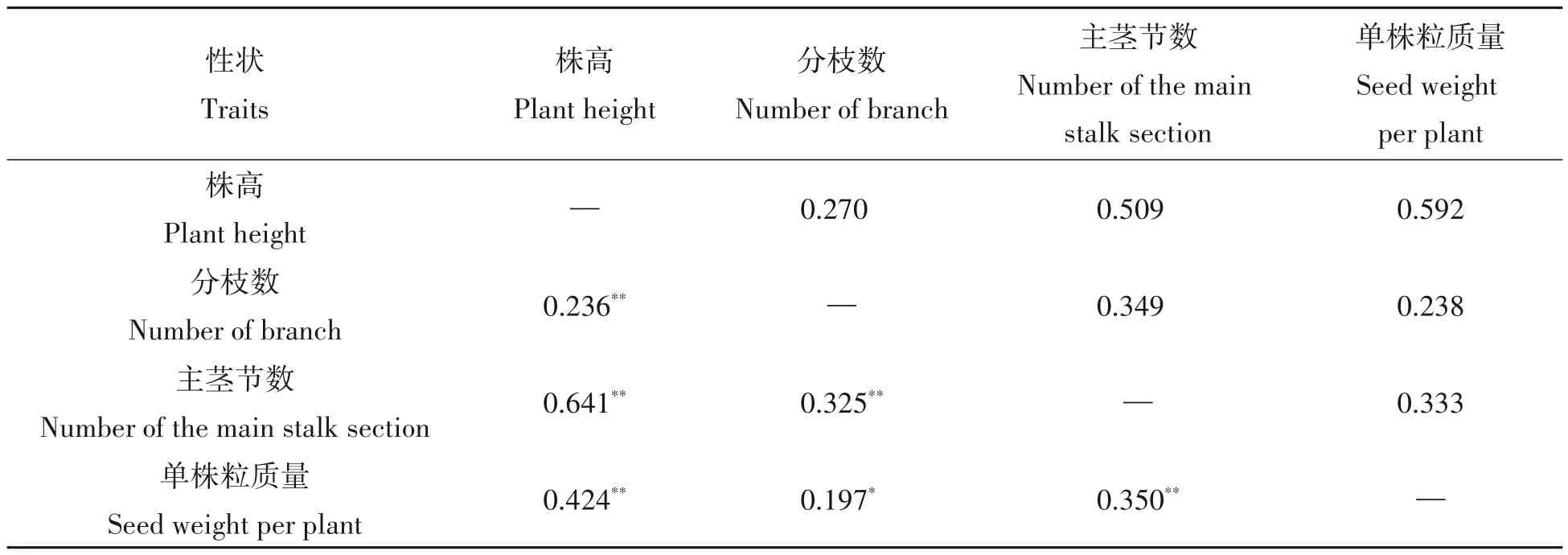
表3 各农艺性状间的表现型相关及遗传相关Tab.3 Phenotypic and genetic correlation between four agronomic traits
3 结论与讨论
产量是农作物的育种中最重要的育种目标。通过改良株型,培育具有理想株型的新品种,可进一步发挥作物增产潜力。同样,在苦荞其株型性状的遗传分析对其育种的理论、选育方法和策略非常重要。本研究通过对苦荞株型相关性状进行了遗传分析、各性状间的表现型相关和遗传相关分析,得出苦荞主要株型性状的遗传规律及相关性,以期为今后苦荞株型改良提供参考。
本研究结果表明,F2群体和F3群体的各农艺性状都出现超亲分离,表明云荞1 号和米荞的杂交后代群体中出现了与亲本不同的基因型,也说明了这两个亲本选配在其后代都会选择到超双亲性状的株系。变异系数的大小反映品种固有特征及品种间的个体差异,是性状遗传多样性的具体体现,性状变异系数越大,遗传背景越丰富,越有利于品种鉴定[22]。株高、分枝数和主茎节数的3个株型性状中株高的变异系数为20.5%,较其他性状大,表明在株型性状中株高具有最大的选择与提高潜力。分枝数的变异系数相对较小,表明比其他株型性状较稳定。
本研究结果表明,株高、主茎节数和单株粒质量的显性方差大于加性方差,并平均显性度大于1。这与在苦荞上利用同样分析方法测定的Li 等[14]的株高和单株粒质量一样的结果,表明株高和单株粒质量容易受环境因素的影响,因此在后期世代进行选拔为有效。但是这与闫玉莹等[23]在甜荞重组自交系的株高遗传率为高遗传率性状,进行早期个体选择有效的说法以及颜秀娟等[24]在大豆的杂交后代F2群体的株高遗传率高,所以低世代选择效果较好的说法有矛盾。并且本研究的结果中分枝数的遗传率比其他株型性状较大,可以在初期选择。这与苦荞[20]的一级分枝数遗传率较小,容易受环境影响,育种中不易在前期单株选拔的说法互相矛盾。这是由于作物、分析方法的不同而引起的,并且所有性状的遗传率与环境都有不同程度的互作效应,说明育种时对这些性状的选择应与适当的环境相结合才能有效地进行。另外,在本研究中各农艺性状的狭义遗传率从F2代、F3代到固定遗传率逐步增高,表明对这些杂交组合来说这些性状是繁殖到后期世代提高遗传率以后进行选拔是最有效的。
株型与产量的相关关系,在水稻、玉米、小麦等作物上已有较多报道[25-27]。了解性状间的相关关系,在选择性状中提供科学决策。研究表明,株高、分枝数和主茎节数与产量具有显著的相关性,其中株高对苦荞产量的综合作用较大。该结论与李春花等[14]研究的结论一致。另外,分枝数和主茎节数都与株高具有极显著的相关性,可以说各性状对产量除了直接作用以外还有间接作用。若想获得高产的目标材料,具有较高的遗传率的分枝数可以在早期世代进行选择,后期世代进行与产量相关系数较大的株高的选择,但是产量与株型性状是相互关联的,在选择育种中对各个性状的选择要综合考虑,才能达到最优的选择效果。
