智能抗逆微生物细胞工厂与绿色生物制造
2020-11-18许可王靖楠李春
许可,王靖楠,李春,
(1 清华大学化学工程系,工业生物催化教育部重点实验室,北京 100084; 2 北京理工大学化学与化工学院,生物化工研究所,北京 100081)
当前,全球81%以上的能源消耗依赖于化石燃料,大气中的CO2浓度近几十年来从(2~2.8)×10-4μL/L 急剧上升到4×10-4μL/L,导致严重的温室效应。如果持续目前的碳使用模式,预计到21 世纪中叶,大气中CO2水平将上升到5×10-4μL/L,全球24%的动植物将面临灭绝[1]。全球现有的发展模式过于依赖化石资源,以破坏环境为代价,必须寻求可持续发展之路[2]。利用可再生和低CO2排放的生物原料取代化石原料成为全球关注的热点。将生物元素引入到现有的、传统的化工制造,发展绿色、可再生、安全可控的生物制造,解决目前人类面临的环境问题、资源危机和能源可持续发展问题势在必行[1]。鉴于此,科学家提出了全新的绿色生物制造体系。绿色生物制造以工业生物技术为核心,结合化学工程技术,从生物质等可再生原料出发生产化学品、能源、材料、医药和食品等,可从源头上降低碳排放,实现绿色清洁的生产工艺,改善化工产业“高能耗、高排放”和依赖化石资源的现有模式[3]。
随着科技创新和技术进步,绿色生物制造的产品空间进一步拓展,发展前景更加广阔,但也面临一些问题,如在生产过程中,生产菌株经常会受到培养环境扰动或自身代谢产物影响,对自身生长或代谢造成不利,称为逆境胁迫[4-5]。包括高温、酸碱扰动、有机溶剂、毒性副产物及机械损伤等[6-9],在这些逆境胁迫下进行生产,会导致生产菌株活力下降、产能降低、生产过程能耗大幅升高等,严重制约着绿色生物制造产业的发展[2,10-11]。随着生物技术与合成生物学的迅猛发展,产业界对能自主响应发酵环境变化而又自行稳定生产性能的优良、智能和高效菌株的需求非常迫切。本文作者将从工业菌株的环境胁迫因子、微生物抗逆分子机制、智能抗逆基因线路设计等方面开展智能微生物细胞工厂的设计构建与绿色生物制造的论述。
1 绿色生物制造中微生物细胞工厂面临的环境胁迫因子
生物发酵过程是绿色生物制造的核心。微生物利用生物质作为原料,广泛生产燃料、氨基酸、表面活性剂、洗涤剂等产品,生物经济已从“粮食生物质”步入“非粮生物质”时代。然而,由于生物发酵过程具有产热、酸碱度变化、酶催化影响、原料产物影响等因素,使高成本和高能耗逐渐成为产业特征。
生物乙醇作为绿色生物制造产业的典型产品,是目前全球最主要的液体可再生燃料,也是公认的汽油最佳替代燃料[12],作为生物乙醇发酵的“核心”,酿酒酵母的性能决定了生产过程的经济性和绿色指数。早在2004 年,Ingledew 等[13]对影响酵母生长和发酵各因子的“警戒水平”进行了研究[图1(a)]。他们发现,单一胁迫因子超出“警戒水平”会对酵母的生长及生产性能产生较大的影响,且各胁迫因子共同作用会产生负协同效应。基于此,分析生物乙醇现行发酵工艺面临的环境胁迫因子如下。
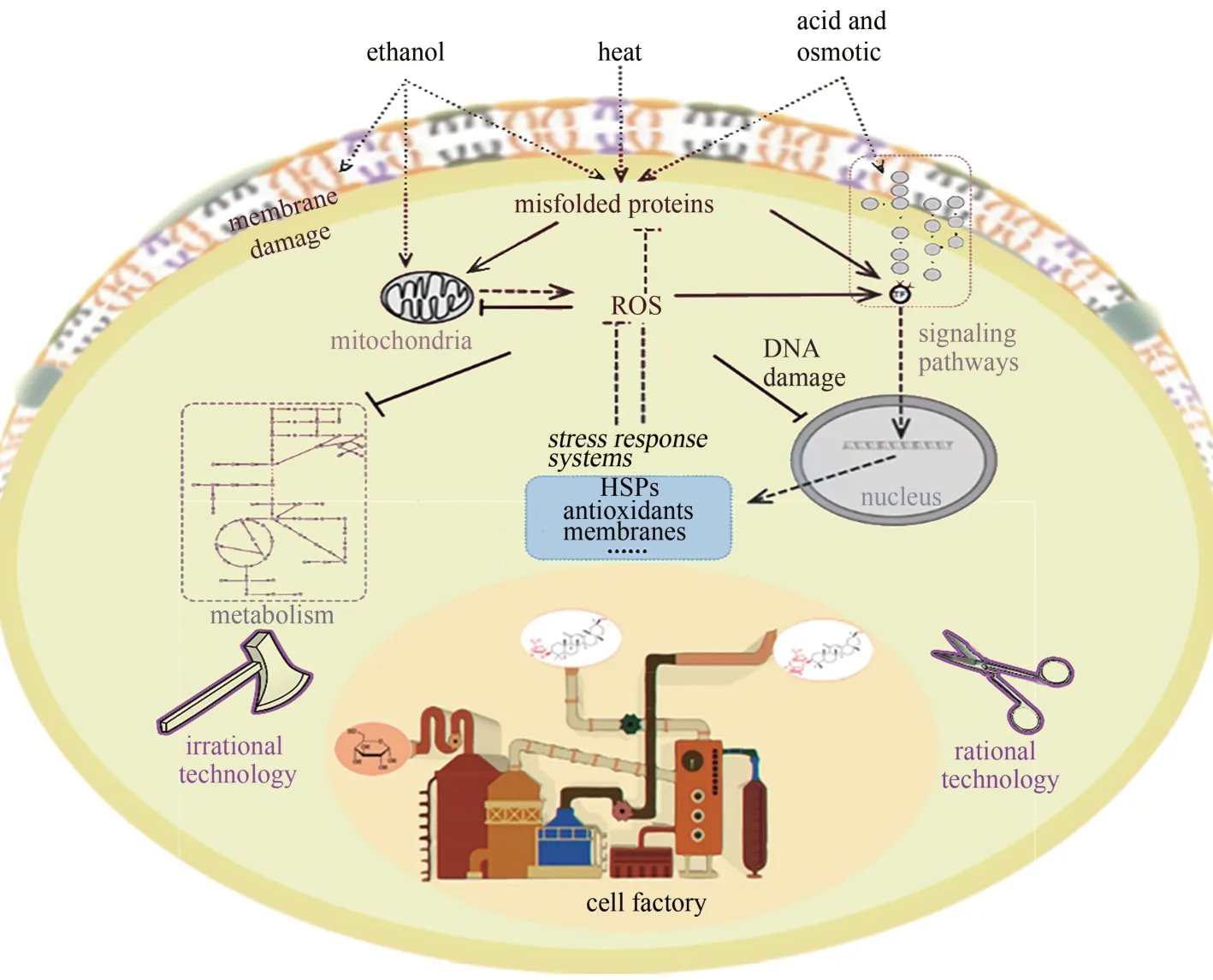
(1)高温胁迫—酵母菌最适生长温度通常在30℃左右,但是在生物乙醇发酵过程中,由于受到外部因素和酵母细胞自身代谢热的影响,发酵罐内温度一般维持在32~37 ℃[14],出现生长最适温度和真实发酵温度不匹配的情况[15]。发酵温度的升高导致酵母细胞蛋白质、染色体、细胞膜、细胞器等结构的破坏,且进一步引起细胞内活性氧自由基(ROS) 的升高, 产生氧化毒性[图1(b)][16]。
(2)高浓度乙醇胁迫—发酵中后期,高浓度乙醇对酵母造成有机溶剂胁迫,亦称为产物抑制[17]。有研究发现,当乙醇体积分数<3.0%,其抑制作用可忽略,当乙醇体积分数>4.0%,对酵母菌株生长明显抑制,当乙醇体积分数进一步提升到>12.0%时,细胞膜的完整性遭到破坏,且严重影响蛋白质的折叠和染色体的正常功能,酵母生长和发酵停止甚至迅速死亡[图1(c)][18-19]。
(3)高糖醪导致的渗透压胁迫—发酵前期,高浓度糖醪液造成发酵环境渗透压升高,致使酵母脱水、导致细胞变形[20]。有研究表明,酵母细胞的最适渗透压与2.6%的葡萄糖溶液相当[21]。当底物中葡萄糖浓度>5%时,即使有足够的氧存在,酵母细胞的生长速率也会明显下降[22]。
(4)酸胁迫—发酵过程中pH 的扰动会对宿主细胞产生极大影响。燃料乙醇单个发酵罐容积均在上千立方米,从节省碱消耗费用及减少酸碱中和产生盐的角度,工业生产中均不控制发酵过程pH,由于杂菌污染及乙醇发酵过程产生副产物有机酸和CO2等,发酵初期pH 逐渐降低至2.5 左右,发酵中期随着碳源的逐渐消耗,发酵过程副产物有机酸部分被吸收,pH 可逐渐回升[23]。酸胁迫导致细胞内环境的持续酸化,破坏对酸敏感的DNA,降低部分中性糖酵解酶的酶活,严重情况下会导致胞内蛋白质变性,最终影响细胞的生长[24][图1(d)]。
胁迫因子几乎存在于所有微生物发酵体系中,并且通常表现为多因子同时存在,产生胁迫效应的叠加,严重制约了生物制造产业的发展[14]。因此,构建在多重胁迫条件下有良好智能表现的高版本微生物细胞工厂,为绿色生物制造迎来了新的机遇,也提出了新的挑战。
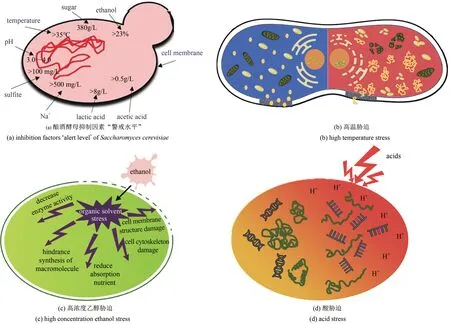
图1 酿酒酵母发酵生产乙醇过程中面临的胁迫因子[13-24]Fig.1 Stress factors faced in ethanol production by Saccharomyces cerevisiae[13-24]
2 微生物细胞抵抗环境胁迫的分子机制
微生物细胞工厂在胁迫环境下,其自身的基因表达、蛋白质和代谢物会发生一系列的动态变化,调节内在胁迫防御系统以维持细胞正常功能[25]。
2.1 单一胁迫因子在酵母细胞的传导途径
以酿酒酵母为例,研究人员针对不同的单一胁迫因子在酵母细胞的传导方面进行了相关研究。作为对热胁迫的反应,酿酒酵母通常表现为下列基因的转录变化:代谢相关酶(如己糖激酶、甘油醛-3-磷酸脱氢酶、6-磷酸脱氢酶等),抗氧化酶(如硫氧还蛋白3、硫氧还蛋白卟啉),热激蛋白及其辅因子(如hsp104、hsp82、hsp60、hsp30、cpr1、sti1和zpr1)和全局转录因子(如hsf1、msn2/4 和yap1)等[26];在高渗胁迫下,细胞生长和应激反应的信号网络是相互协同的,RAS-PKA 途径、细胞壁完整性途径和SVG 途径与胁迫信号传导关系紧密,关键基因BCY1和Ras2的敲除可以增加RAS-PKA 通路的活性,提高对高渗的抵抗[27];乙醇胁迫影响酵母细胞中包括葡萄糖和氨基酸的代谢,降低糖酵解途径酶活。液泡和膜上的H+-ATPase 系统似乎在乙醇胁迫的恢复过程中起着重要作用[28];ROS 的积累和小热激蛋白的表达波动则出现在多种胁迫因子的传导过程[29][图2(a)]。
2.2 热激应答系统(heat shock response system)
热激应答系统由热激转录因子(HSF)进行调控,相关基因涉及蛋白质合成、碳源代谢及细胞结构等[30]。HSF 通常状态下以单体或二聚体状态存在,在胁迫条件下,无活性的HSF 形成多聚体,造成构象的改变,使得活性区域暴露出来,与热激元件的启动子上长度为5个碱基的保守序列发生特异性结合,进而转录相关基因并翻译形成热激蛋白(HSP)。HSP 除主要参与维持蛋白质天然构象外,同时参与细胞的多种生理调节,对其抗胁迫提供帮助[31][图2(b)]。

图2 微生物细胞抵抗环境胁迫的分子机制[29-33,39]Fig.2 Molecular mechanism underlying microbial cells'response to environmental stresses[29-33,39]
2.3 抗氧化防御系统(antioxidant defense system)
抗氧化防御系统由酶促防御体系和非酶促防御体系组成。胁迫环境会导致细胞内大量活性氧自由基积累,细胞面临氧化胁迫,酶促防御体系启动,清除胞内活性氧自由基和过氧化氢。超氧化物歧化酶(SOD)通过清除机体内·O2-来保护细胞;过氧化氢酶可以促使H2O2分解,清除细胞内的H2O2;过氧化物酶行使多种保护功能,包括抗氧化、分子伴侣功能和调节信号转导等。非酶防御体系由小分子物质组成,包括谷胱甘肽、硫氧还蛋白、维生素C等,作为还原剂清除自由基的小分子[17,32-33][图2(c)]。
2.4 蛋白质质量控制系统(protein quality control system)
蛋白质质量控制系统通过维持胞内蛋白质平衡提高细胞的生存能力[34]。该系统具有两部分功能:变性蛋白重折叠与新合成蛋白的折叠组装;变性失活蛋白的降解。以酵母细胞为例,其蛋白质质量控制系统主要包括自噬系统和26S泛素蛋白酶体系统。自噬系统是一种介于蛋白质水平与细胞水平的蛋白质质量控制系统,可以清除细胞中的问题蛋白,从而减少细胞因变性蛋白质聚集而产生的毒害,在维持各种自由氨基、脂肪酸和有机物的循环利用、保持细胞内平衡中起到重要作用。26S 泛素蛋白酶体系统由泛素、泛素启动酶系统以及蛋白酶体组成,泛素、泛素启动酶系统负责变性蛋白的泛素化,蛋白酶体被称为“细胞内的再生与回收中心”,泛素化的蛋白在此被分解为短肽和游离氨基酸,而泛素被回收再利用[35-36]。
2.5 微生物膜系统(microbial membrane system)
微生物膜系统作为一种生物屏障,在调节微生物的耐受性方面起着重要作用。膜损伤通常被认为是多种胁迫因子的主要作用机制之一,在胁迫环境下,微生物膜起到保护屏障的作用,将细胞质从细胞外胁迫环境中分离出来[37]。微生物细胞工厂在很大程度上依赖于膜内稳态和膜内脂蛋白相互作用,涉及诸多关键的生理功能,包括能量代谢、溶质转运、信号转导、细胞生长等[38],因此,维持膜内稳态,包括调节脂质和蛋白质组成并维持其生物物理特性,是维持细胞工厂性能行之有效的方法[39][图2(d)]。
2.6 海藻糖调节系统(trehalose regulation system)
海藻糖是一种性质稳定的非还原性二糖[40],广泛存在于低等植物和微生物中。海藻糖是大多数昆虫和真菌的主要储能物质,也是一种良好的生物保护剂,能够保护细胞中的生物大分子免受干燥、高温和高渗透压等胁迫环境的破坏[41]。近年来,科研人员对海藻糖的作用机制及其代谢调控进行了一定的研究。结果表明,海藻糖具有在胁迫条件下防止酵母体内蛋白质聚集和帮助蛋白质正确折叠的功能,是一种在生物界广泛存在的具有生物抗逆保护作用的重要物质。尽管海藻糖在不同环境胁迫下对生物的保护机制尚不完全清楚,但可以肯定的是,这些重要的生理功能是与其独特的理化性质分不开的[42]。
除以上的抗逆机制外,细胞内糖蛋白的表达也与细胞抵御外界胁迫密切相关[43],最近研究表明,钙稳态有可能是胁迫环境下真核细胞存活的关键因素[44]。
3 非理性技术对抗逆性能的提高
研究人员在充分了解微生物抗逆机制的基础上,采用非理性技术增强细胞自身防御系统,提高其抗逆性能[45]。
3.1 非理性改造结合反向代谢工程策略
结合微生物细胞的胁迫因子传导,可以采取反向代谢工程策略进行酵母细胞抗逆的研究。根据基因组、转录组、数量性状基因定位(QTL)等数据比较,挖掘潜在的关键基因,进一步通过基因敲除、过表达来验证基因功能。Nielsen 等[46]采用适应性驯化的方法,得到40 ℃下生长的耐热酵母菌株,并通过转录组研究了关键作用基因。Alper等[47]采用全局转录调控方法通过突变或过表达全局转录因子,增加了酿酒酵母对高温、乙醇及抑制物的耐受[图3(a)]。
3.2 基于分子抗逆响应实现微生物基因组的非理性改造
利用最新的基因编辑技术同样可以实现微生物基因组的非理性改造,并借助高通量筛选得到抗逆性能优秀的细胞工厂。Zhao Huimin 课题组[48]开发了一种在酿酒酵母中实现自动化多基因编辑的方法,以所需的功能基因线路构建cDNA 文库,用CRISPR-Cas 将多个片段整合到基因组,筛选到乙酸耐受和丁醇耐受的酵母菌株[图3(b)];元英进课题组[49]在人工染色体酵母细胞中使用SCRaMbLE 技术进行基因组重排,筛选到耐受42 ℃高温和耐受咖啡因的酵母菌株,并找到了关键作用基因[图3(c)];Snoek 等[50]通过靶向性基因组重排得到了3000 个突变株,进一步筛选得到8 株发酵效果优于原始菌株的突变株;Elela 团队[51]构建了酿酒酵母全部内含子的单独敲除细胞,通过对所有内含子单敲细胞的表型、转录组数据以及遗传性状进行综合分析,揭示出内含子自身独立于其编码基因的通用功能——介导细胞对饥饿的应答。
尽管研究人员基于非理性育种技术获得了具有不同抗逆特征的微生物细胞工厂,但是也凸显出该方法获得的菌株智能响应环境胁迫变化的能力较弱。究其原因,细胞内部防御系统大多是胁迫环境下的应答系统,无法使微生物获得稳定的抗逆性能,且常常以牺牲微生物的生产性能为代价,表现为生产菌株在多重胁迫下生产性能大幅降低。如何实现细胞工厂智能响应发酵过程的环境变化进而实现刚柔并济是亟待解决的问题。
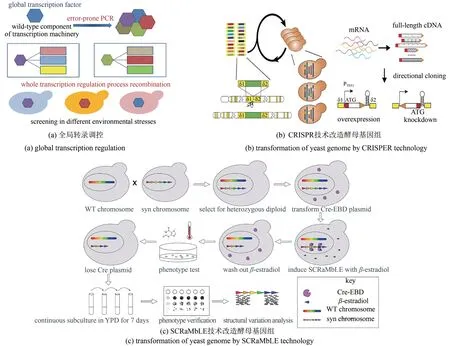
图3 非理性技术增强细胞自身防御系统提高其抗逆性能[48-49]Fig.3 Random and semirational technologies to engineer microbial cell self-defense systems for stress response[48-49]
4 抗逆基因线路设计与智能型细胞工厂的构建
合成生物学融入了工程学思想和策略,将自然存在的基因元件标准化、去耦合、模块化来设计新的生物系统或改造已有的生物系统[52-53]。随着合成生物技术的发展,基于分子层面的人工合成体系的构建得到广泛关注[54],这使得对鲁棒性和智能型细胞工厂的需求更为迫切,重编程微生物时代正在到来。
极端环境微生物代表着生命的极限,蕴藏着丰富的特殊功能基因资源,有着不可估量的生物技术开发前景[55-56]。通过研究极端环境微生物的抗逆分子机制,运用合成生物学技术将其植入到细胞工厂的改造,可以极大提升生物制造产业的能力[57-58]。
当前,抗逆基因线路设计思路主要是通过系统设计和改造,对极端环境微生物抗逆基因及调控序列进行发掘、分类和定义;克隆或人工合成相关基因和调控元件;进一步进行抗逆基因线路模块化组装,研究底盘宿主与抗逆基因线路的适配性机理等[59]。
4.1 微生物智能热量调节引擎的设计
嗜热菌可以在高温条件下生长,不但可以降低发酵过程反应系统的冷却能耗,而且不易被中温微生物污染。李春等[60]通过挖掘及分析嗜热微生物的耐热元件,发明了微生物热量智能调节引擎(IMHeRE)。IMHeRE的基因线路包括从嗜热微生物中筛选得到的HSP作为耐热功能元件,以及人工设计的RNA温度响应开关作为调控元件,利用对不同温度响应的RNA温度响应开关调控在不同温度下效果最佳的HSP 实现了工程菌株的梯级耐热。李春等[60]进一步研究了IMHeRE与底盘宿主的适配性,并对模式工业宿主大肠杆菌和酿酒酵母进行了模块化组装与适配性研究,实现了菌体耐热与细胞数量调控的智能化,拓宽了其最适生长温度的范围,使大肠杆菌在37~43 ℃、酿酒酵母在30~40 ℃正常生长,大幅度提高了底盘宿主的耐热性,IMHeRE应用于赖氨酸40 ℃高温发酵,发现赖氨酸产量较对照提高了2~5倍[图4(a)]。

图4 抗逆基因线路设计及鲁棒性、智能型细胞工厂的构建[15,60-61,63]Fig.4 Designing of tolerant gene circuits to construct robust and intelligent cell factory[15,60-61,63]
4.2 蛋白质质量控制系统提高微生物细胞工厂的耐受性
生物发酵过程中,多种胁迫因子协同作用导致生产菌株的功能蛋白变性失活,造成其蛋白质内稳态失衡,严重影响细胞的生长代谢。维持胞内蛋白质平衡是提高细胞工厂多重耐受性的关键。Xu等[15]从嗜热菌、耐热菌及酿酒酵母蛋白质质量控制系统中挖掘并验证了大量基因,根据其功能相关性,组合成多功能基因线路,构建了人工蛋白质质量控制系统(APQC),并将其应用于酿酒酵母高温乙醇发酵。高温发酵培养及生理特性研究表明,APQC与单功能耐热基因线路相比,能更好地提高酵母的耐热性、细胞活力与细胞壁完整性,证明APQC可以在多重胁迫因子环境下有效调控酵母细胞内蛋白质代谢平衡[图4(b)]。
4.3 基于抗氧化防御系统提高微生物细胞工厂的耐受性
好氧生物在进行有氧代谢的过程中会产生ROS,进而造成细胞膜脂质氧化及核酸损伤等。发酵过程中,多种胁迫因子协同作用加速ROS 的积累,导致氧化胁迫。Xu等[32]基于对酿酒酵母抗氧化防御体系的分析,挖掘嗜热栖热菌和酿酒酵母内的抗氧化蛋白,初步表征和探讨抗氧化蛋白的性能,并将验证结果较好的基因组合成多功能基因线路,构建人工抗氧化防御系统并将其整合到工业酿酒酵母基因组,经发酵验证表明,工程菌株细胞生长量及乙醇产量均比对照有明显提高,ROS 含量测定和线粒体完整性检测结果显示导入人工抗氧化防御系统可以减少ROS 对宿主细胞的胁迫。
4.4 细胞膜工程化设计
膜损伤通常被认为是胁迫导致细胞毒性的主要机制。研究人员可以通过工程化设计改变磷脂分布强化微生物膜的功能。Tan 等[61]在大肠杆菌中提出了一种新的磷脂头部工程策略,通过增加磷脂酰丝氨酸合成酶(+pssA)的表达显著提高了大肠杆菌对辛酸和其他相关抑制剂如糠醛、乙酸、甲苯、乙醇和低pH 的耐受性。工程菌株中,除磷乙醇胺(PE)头部基团相对丰度增加,脂肪酸尾部组成也发生变化,导致菌株在膜完整性、表面电位、电化学电位和疏水性、细胞内酸化敏感性等方面均有显著变化。进一步研究表明,磷乙醇胺含量的增加是导致双层膜厚度增加的原因,而疏水核厚度的增加是脂肪酸头部和尾部同时改变的结果[图4(c)]。
4.5 基于核糖体开关的pH感应基因线路
Pham 等[62]认为,环境pH 是指导活细胞新陈代谢的基本信号,细胞对环境pH 的精确响应对于其日益复杂的功能化至关重要,针对这一情况,他们设计了一套基于核糖体开关的pH 感应基因线路,根据不同的环境pH 控制基因表达。基因线路中应用的核糖体开关可以精确调控pH 依赖性基因表达,其OFF-ON 输出差异最大为31 倍。为了进一步阐明pH 感应基因线路的应用,研究人员将基因线路、易错PCR 和荧光细胞标记综合应用,对大肠杆菌进行自主进化编程和耐酸表型富集,获得了耐受性显著提高的细胞工厂。
4.6 多重防御系统构建抗逆工业酵母及其应用
Li 等[63]为应对多重胁迫因子的协同作用,提出了构建微生物多重防御系统的研究思路。他们将具有不同抗逆性能的基因元件进行重排组合,采用Golden Gate Assembly 进行包含多个抗逆基因线路的多重抗逆防御系统的快速构建,并将不同组合方式的多重抗逆防御系统高效转化工业酿酒酵母,获得携带不同抗逆机制的工业酵母菌株库。采用燃料乙醇工厂工艺水模拟发酵物料配制培养基,利用高通量全自动筛选仪器平台模拟工业高温、高糖、乙醇毒害等多重压力,得到了一株在工业液化醪中高产乙醇的菌株A223。液化醪中试发酵结果表明,A223 乙醇产量比出发菌株提高6.9%,总糖残留仅为出发菌株的51%。细胞凋亡水平、细胞膜完整性、线粒体完整性、活性氧及丙二醛含量均表明工程菌株的耐受性高于出发菌株。转录组分析表明,在多重胁迫下,工程菌的线粒体氧化磷酸化、三羧酸循环和过氧化物酶体的代谢水平明显加强。显著降低了发酵过程的控温能耗,提高了糖醇转化率,发酵能耗降低45.1%,提高产值近0.4亿元[图4(d)]。
4.7 动态调控酵母胞内活性氧与乙酸含量
木质纤维素是丰富的可再生资源,可通过发酵生产各种生物燃料和化学品。酿酒酵母是目前利用木质纤维素生产生物乙醇的优良菌株。与实验室培养相比,酵母在工业木质纤维素发酵中受到多重胁迫压力,例如高温、高糖和乙酸等。部分压力会造成胞内活性氧上升,使细胞生产率受到压力的阻碍。增强木糖发酵酵母的健壮性对于改善木质纤维素乙醇的生产很重要,Qin等[64]采用胁迫驱动的启动子构建抗逆基因线路,增强了酿酒酵母中谷胱甘肽的生物合成途径和乙酸降解途径,进而增强酵母对活性氧和乙酸升高引起的胁迫的耐受性。作者从转录组中挖掘的应激驱动启动子,实现抗逆遗传线路的动态反馈调节,维持细胞内低活性氧的同时减轻代谢负担,最终提高酵母的健壮性和乙醇产量。在胁迫和非胁迫条件下,工程菌株的细胞生长、木糖利用率和乙醇产量均得到提高。该工程菌株在36°C 的实验室培养基和工业木质纤维素培养基中的乙醇生产率分别比其亲本菌株高49.5%和17.5%。使用工业木质纤维素培养基(乙二胺预处理的玉米秸秆),通过同步糖化共发酵,工程菌株96 h可生产90.7 g/L乙醇[64]。
5 结 语
综上所述,绿色生物制造是关系到未来国际竞争地位的战略性新兴产业,其发展对合成生物学技术提出了新的要求:以工程化思维、借助合成生物学技术有效改善微生物细胞工厂的抗逆性能,提高其在胁迫条件下的生存能力。目前该领域研究尚处于初始阶段,局限于应对单一胁迫因子的基因线路的设计。对多重胁迫因子的信号传导方式尚不清楚,不能有针对性地对胁迫信号传导进行阻断调控是制约其发展的重要原因。因此,在了解微生物代谢调控的基础上,从分子水平上研究其抗逆机制,阐明阻断调控细胞工厂胁迫级联传导的分子机制,有针对性地设计并集成多重抗逆防御系统,提高细胞工厂在多重压力环境中的抗逆能力,将极大提高生物制造过程的生产效率,降低发酵能耗和生产成本,实现人工生物反应体系的高效性。未来,研究人员建立新方法、获得新菌株的同时,积极推动成果应用,将有效带动全国生物制造行业技术提升,加快实现生物产业“十四五”规划目标。
