菊芋作为饲料原料的品种选育、营养价值及其饲用效果研究进展
2020-11-15晏国生熊本海
汪 悦, 刘 君, 晏国生, 熊本海*
(1.中国农业科学院北京畜牧兽医研究所,北京100193;2.廊坊市农林科学院,河北廊坊065000;3.廊坊市思科农业技术有限公司,河北廊坊 065000)
菊芋(Helianthus tuberosus)俗称洋姜,属植物分类的菊科向日葵属的多年生草本植物。具有抗盐碱、耐瘠薄、耐旱、耐寒、抗病性强等生长习性(卢秉钧,2004)。其枝叶翠绿、花簇金黄的外观特点,被国人誉为“懒省事”的新兴绿化观赏植物。据对比试验,在同等肥力的耕地上,种植苜蓿可亩产粗蛋白质 138 kg,而菊芋则高达200~250 kg(杨在宾等,2018)。由此可以预测到,菊芋是一种高产优质的新型粗饲料资源。研究表明,菊芋茎秆中含有大量的蛋白质、可溶性碳水化合物、Ca、磷(P)及铁(Fe)等多种营养素。此外,新鲜菊芋块茎中高含量的菊糖可为双歧杆菌和韦氏杆菌等肠道益生菌群提供大量优质的能量底物,优化肠道菌群结构,进而提高动物免疫能力(陈建军等,2001)。因此,菊芋既可作为家畜的优良粗饲料,也是潜在减抗的有效产品。目前,菊芋已成为国际公认的药食同源保健品。
为系统地研究菊芋的饲用价值,有不少研究针对菊芋品系(种)选育以及不同品种、不同年份、不同茬次及不同生产期菊芋的营养价值进行全面分析。结果表明,部分品种的菊芋秸秆在适当的生产期采收,具有较高的CP含量,且Ca、P含量丰富(晏国生,2012)。若用作粗饲料,可部分替代优质苜蓿饲料(巴合提·加布克拜等,2001),且具有明显的价格优势。因此,2019年11月,中国饲料库情报网中心发布的第30版《中国饲料成分及营养价值表》中,首次将菊芋秸秆作为粗饲料纳入中国饲料数据库(表1)。
1 菊芋饲料品种(系)选育
为培育满足市场需求的牧草型菊芋新品种,刘君等 (2019)在定量化育种理论与方法的指导下,运用灰色关联度和同异关系分析原理与方法,制定牧草型菊芋育种目标(表2);采用单株灰色选择和同异选择原理与方法,确定优良单株 (表3);采用品种灰色多维综合评估和同异比较原理与方法,筛选比对品种综合性状显著的牧草型菊芋新品种(系)(晏国生,2016)。
通过系统选育,成功培育出了廊芋19、20、21、22、23和24号,6个廊芋系列牧草型菊芋新品种,使菊芋新品种选育工作在定量化、信息化和科学化的道路上迈出了重要的一步,为菊芋这一新型饲料资源,提供了适于规模种植的品种资源。

表2 菊芋不同品种(系)产量与农艺性状观察值
值得注意的是,与通常相关分析不同,在灰色关联度分析中,一种性状与另一种性状之间的关联度并不具有对称性,即rij≠rji(李玉辉等,2005)。这是因为灰色关联度分析具有整体性,它不仅考虑某性状与另一性状之间的关系,同时也考虑某一性状与其他性状之间的关系,因而其分析结果更符合客观实际(郭瑞林,2011)。
不同品种来源、生境条件及生长阶段的菊芋,营养成分及含量存在较大差异。吕世奇等(2018)将野生菊芋种质LZJ040作为母本,通过对其自然结实种子的收获,结合组培扩繁和田间产量表现,筛选出抗旱高产的兰芋1号。其DM含量为27.85%,菊糖含量为639.5 g/kg,田间抗根腐病能力强,块茎产量高,一般亩产可达3000~3300 kg。孔涛等 (2013)对红菊芋、白红菊芋、球白菊芋、长白菊芋、白菊芋 5种菊芋的块茎及茎叶的营养成分进行分析比较,结果表明长白菊芋块茎菊糖和CP含量最高(分别为 77.5%和8.72%);红菊芋茎叶的水分和EE含量最高 (77.3%和6.14%)、CF含量最低 (5.21%)、水溶性成分(65.88%)和适口性指标(CP/CF>1)皆最高,最适宜作为家畜粗饲料。Seiler等(2004)对德克萨斯州的9个野生菊芋种群在2年开花期间秸秆中的N、P、Ca、Mg、K 和 Ca/P 比率进行了评估, 结果表明用于反刍动物饲料的菊芋秸秆在开花期N、Ca、Mg和K充足,但P不足,导致Ca/P比过高,范围约为4.3~34.3:1,如果使用菊芋作为主要饲料,必须添加P补充剂或含有高浓度P的其他饲料,以降低代谢紊乱的风险。此外,9个种群的N、K、P、Ca、Mg和Ca/P比值均存在基因型差异,表明有通过杂交和选育改善的可能性;而P和Mg的群体差异很小,通过选育难以改善这些差异。

表4 牧草型菊芋品种营养分析研究结果(干基)%
2 菊芋营养成分分析
2.1 菊芋块茎 2018年,中国农业科学院北京畜牧兽医研究所与廊坊菊芋研究所联合攻关,对采集的150个菊芋样品进行块茎养分检测(表5)。分析结果表明,菊芋块茎中脂肪含量极低(或不含),主要成分为蛋白质 (9.81 g/100 g)、糖以及Mg(56.5 mg/100 g)、K(2.21×103mg/100 g)和 Fe(1.84 mg/100 g)等微量元素,氨基酸种类较为齐全(16种),其中精氨酸(1.45 g/100 g)、谷氨酸(0.99 g/100 g)和天门冬氨酸(0.62 g/100 g)含量较高,此外还含有少量黄酮。Annika等(2006)研究表明,菊芋块茎中存在的少量脂肪中仅有微量的单不饱和脂肪酸和多不饱和脂肪酸,主要是亚油酸(C18:2n-6)和亚麻酸(C18:3n-3),但没有饱和脂肪酸。在整个生长阶段,块茎中的蛋白质和氮水平保持相对恒定 (Jantaharn等,2018)。 Somda等(1999)研究了菊芋(品种名为 Sunchoke)从种植到贮藏块茎中营养元素分配,结果表明在快速生长阶段,块茎中的碳和韧皮部移动性较大的元素水平显著增加。到收获时,在成熟的块茎中发现了高水平的K、P和Ca。

表5 菊芋块茎全粉营养成分及含量
晏国生(2012)对30种菊芋样品进行块茎中糖成分及含量的检测(表6)。结果表明,菊芋块茎中,葡萄糖、蔗糖、果糖等还原糖含量较低(约占总糖的25%~32%DM),而总糖中70%以上是菊糖。 据报道,菊芋和菊苣是天然菊糖最主要的来源,并且与大多数将淀粉作为碳储存的作物不同,菊芋的主要碳储存是菊糖 (Khusenov等,2016),块茎中菊糖含量占块茎鲜重的8%~21%(约为干重的 70% ~ 80%)(Wilkins等,2008)。作为一种植物多糖,菊糖聚合度(dp)是影响其生理活性的重要因素。李琬聪(2015)研究了菊芋块茎整个生长过程中菊糖dp对生物活性的影响及变化规律。通过对比7种特定dp菊糖的益生活性发现,二聚(dp 2)菊糖活性最差,而三聚(dp 3)菊糖和四聚(dp 4)菊糖益生活性最佳,五聚(dp 5)之后菊糖益生活性减弱,但dp 5~ dp 8菊糖间的益生活性变化不明显。

表6 菊芋块茎糖成分及含量%
2.2 菊芋茎叶 本实验室在前期研究过程中对30个菊芋样品茎杆进行了概略养分分析 (表7)。研究表明,不同品种菊芋茎秆间的养分存在一定差异,CP含量最高可达15.22%DM,CF含量为9.49% ~12.73%DM,EE水平为 1.27% ~2.74%DM。茎中的菊糖从顶部向底部逐渐累积,具有较高dp的菊糖多集中在茎中部,而低dp菊糖更常见于茎的基部(Silva 等,2015)。 Kyu-Ho(2013)在成熟的茎中分别分离出含有4、6、8和12个dp的菊糖。茎叶中主要矿物质是 K、Na、Ca、Mg和 P(Hay等,2006)。最研究发现,茎叶中的 Ca、P 和 K含量分别约为块茎中的7.5、5.2和4.4倍(Mattonai等,2018)。
菊芋叶片的CP含量约是块茎的5倍,约是茎秆的3倍(Rawate等,2005)。叶片中水解氨基酸的总含量可达叶片干重的12.45%,其中脯氨酸、天门冬氨酸、谷氨酸和亮氨酸为菊芋叶片中氨基酸的主要组成成分,而脯氨酸含量最高(Peksa等,2016)。叶片中的氮含量从幼叶中的35%逐渐降至老叶中的12%,CP含量从17.3%DM减少到13.3%DM(Seiler,2004)。叶片 β-胡萝卜素(371 mg/kg)和维生素C(1662 mg/kg)的水平相对较高,是块茎的约4~8倍。茎比叶子含有更多的纤维素和半纤维素。除了果糖之外,叶子和茎中发现的糖主要是葡萄糖,还有一些蔗糖、木糖、半乳糖、甘露糖、阿拉伯糖和鼠李糖(Malmberg等,2006)。
建立菊芋作为饲料的主要概略养分数据库,为牧草型菊芋品种进入国家数据库奠定了科学基础,大大丰富了《中国饲料数据库》饲料类型及数据资源,为菊芋新型饲料行业的发展提供技术支撑。
3 菊芋营养价值突出优势
与牛、羊常用粗饲料紫花苜蓿、玉米秸秆和黑麦草典型养分(干基)对比,菊芋茎叶主要具有以下五大突出优势:
3.1 CP含量高 尽管菊芋秸秆CP(7%)低于盛花期苜蓿草(16%)的含量,但明显高于其他2种粗饲料,分别比玉米秸秆和黑麦秸秆高2%和3%,比玉米芯高4%。其中,菊芋叶粉CP为19%,相当于初花期苜蓿干草CP含量;菊芋全株CP含量为10%,比成熟期苜蓿干草CP含量低3%,但与苜蓿茎(11%)及黑麦干草(10%)CP含量基本相同(熊本海等,2018)。
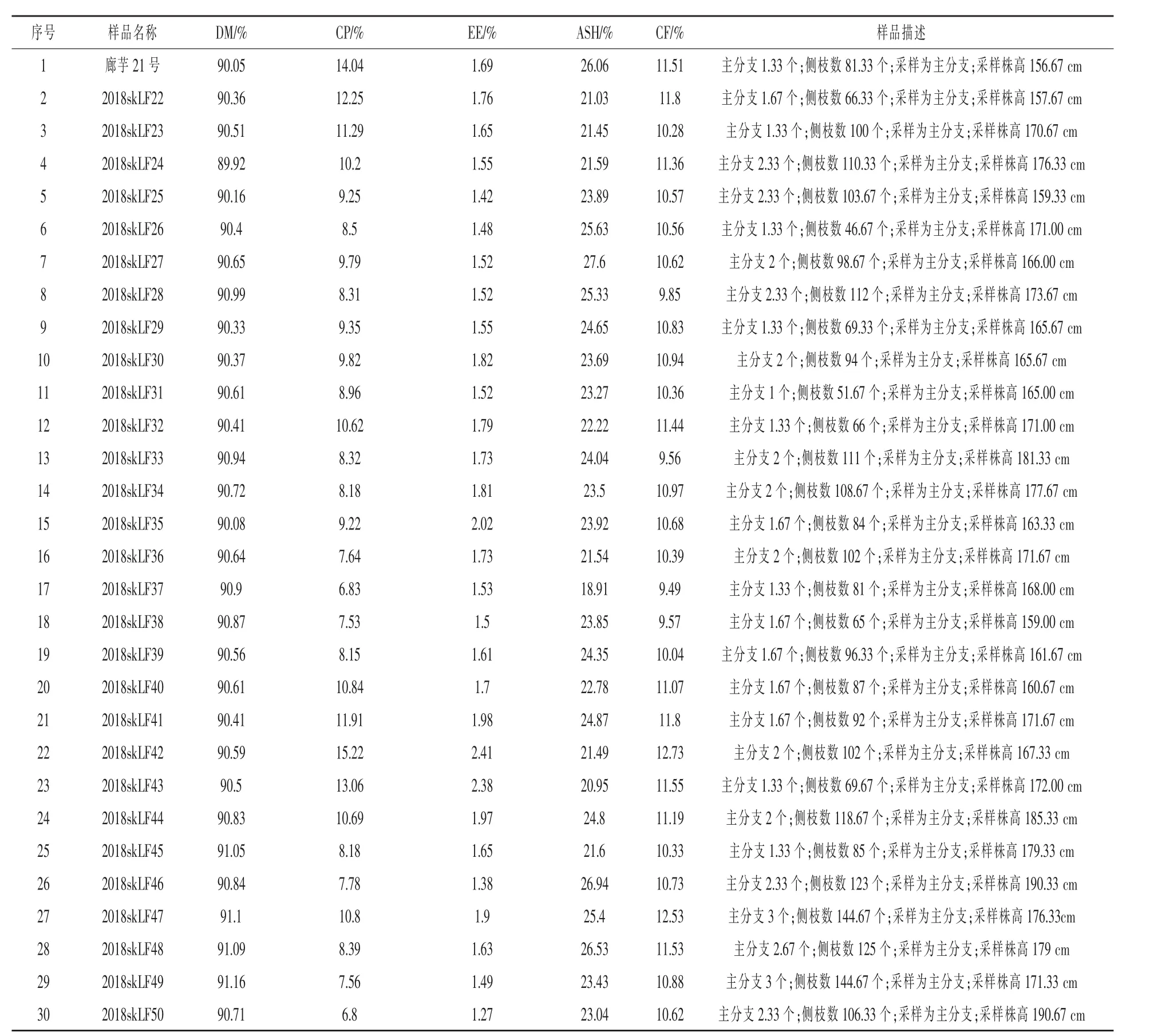
表7 菊芋茎秆概略养分分析
3.2 富含Ca、P等矿物元素,且二者比例适宜大量研究表明,紫花苜蓿是奶牛等草食动物所需Ca的良好来源。而现在数据证明,菊芋秸秆Ca含量明显高于苜蓿、玉米秸秆和黑麦草。菊芋叶粉、全株和秸秆的Ca含量分别为17%、12%和10%,均显著高于苜蓿(8% ~10%)、玉米秸秆(5% ~7%)及黑麦草(6%~8%)等粗饲料。菊芋全株的P含量为0.45%,苜蓿草为0.18%~0.30%,玉米秸秆为0.19%~0.28%,黑麦为0.04%~0.30%。菊芋全株的 Ca、P比例约为 2.1:1 (熊本海等,2018)。
3.3 CF含量适中 牛羊等反刍动物可充分利用粗饲料,通过瘤胃发酵提供70%~80%的能量需要,而其对粗饲料利用效果取决于粗饲料中CF的含量及结构(李娜等,2016)。日粮中适当比例的粗纤维能提高奶牛的产奶量,且乳脂率也有一定程度的增加。根据奶牛的生理特点及生产需求,日粮中粗饲料比例应在40%~70%,其中CF含量应占干物质的15%~24%,才能保证牛体健康(Karen等,1996)。菊芋全株 (廊坊)的CF含量(31%DM)与中花期苜蓿干草(30%DM)和黑麦干草(33%DM)含量接近。菊芋全株(廊坊)ADF和NDF(分别为40%DM,53%DM)含量与盛花期苜蓿干草(40%DM,52%DM)极为相近;菊芋叶粉ADF含量(22%DM)与苜蓿叶粉ADF含量(25%DM)相近。另外,菊芋秸秆(廊坊)的有效NDF(eNDF)含量(100%)与苜蓿茎(100%)、黑麦秸秆(100%)及玉米秸秆(成熟期)(100%)含量相同(熊本海等,2018)。而eNDF是有效维持乳脂率稳定总能力的饲料特性(闵晓梅等,2002)。
3.4 EE含量丰富 菊芋叶粉EE含量为3.64%,与玉米芯粉(3.7%)含量接近,明显高于苜蓿叶粉(2.7%);菊芋全株的EE含量为2.1%,与中花期和盛花期苜蓿干草的EE含量 (分比为2.3%和2%)大体相当(熊本海等,2018)。
3.5 泌乳净能 (NEL)具有优势 菊芋秸秆的NEL高于苜蓿茎及黑麦草,但低于带穗玉米秸秆。其中,菊芋秸秆的NEL为1.03 Mcal/kg,带穗玉米秸秆为1.45 Mcal/kg,黑麦秸秆为0.97 Mcal/kg,苜蓿茎为 1.01 Mcal/kg(熊本海等,2018)。
4 菊芋在动物生产中的应用
4.1 在单胃动物生产中的应用 菊芋在单胃动物生产中的应用多集中于块茎粉及菊糖对动物生产性能与生理机能方面的调控。王亚锴(2013)研究表明,肉用仔鸡日粮中添加 0.9%的菊芋块茎粉,其日均采食量和平均日增重分别提高5.64%和7.07%,血清中IgA和IgG浓度分别提高14.95%和19.94%,脾指数和法氏囊指数分别提高27.18%和23.62%。此外,块茎中的菊糖对肉仔鸡干物质代谢率和表观代谢也有极显著的提高作用。Nabizadeh等(2012)报道,菊糖能显著提高肉用仔鸡盲肠产丁酸细菌的浓度或代谢活性,降低消化道 pH,提高十二指肠淀粉酶和空肠胰蛋白酶活性,有利于肉仔鸡养分的消化和代谢。同样,在对9~12周龄生长猪的试验中发现,菊糖能够降低盲肠pH并增加丁酸浓度,而丁酸有助于结肠上皮细胞增殖,刺激结肠黏膜细胞的生长,进而提高营养物质的吸收率(Sattler,2014)。在180只仔猪(7周龄)的基础饲料中添加5%菊芋块茎粉,5周后屠宰,取胃肠内容物和肠样品进行分析。试验结果显示,与对照组(仅饲喂基础饲粮)相比,试验组(基础饲粮+5%菊芋块茎粉)仔猪粪肠球菌和大肠杆菌含量显著减少(P<0.05),粪便乳杆菌显著增加(P<0.05)。此外,在试验组仔猪的空肠中观察到肠绒毛顶端分支、肠细胞中度变性和有丝分裂,且空肠细胞和迁移细胞中凋亡细胞数量明显增加。研究结果表明,用菊芋块茎饲喂可显著改善猪肠道的微生物结构,促进宿主肠细胞防御和再生过程(Valdovska,2014)。 McRorie (2017)提出,在富含碳水化合物的大鼠饮食中添加菊糖能够减少肝脏脂肪酸的从头合成。由于脂肪酸合成酶是通过修饰脂肪生成基因进行表达的,菊糖的摄入使脂肪生成酶和脂肪酸合酶mRNA的活性降低,通过减少肝脏中极低密度脂蛋白-三酰甘油(VLDL-TAG)的分泌,进而降低了菊糖的TAG效应。
菊糖不能被单胃动物自身分泌的消化酶消化,但它进入肠道后段可被肠道内定植的有益菌消化利用,起到有益菌增殖因子的作用。同时所产生的酸性物质可降低整个肠道的pH,从而抑制有害菌(如大肠杆菌、沙门氏菌等)的生长,提高动物的抗病能力(Claus等,2017)。 Kolida 等(2007)报道,双歧杆菌、消化链球菌与克雷伯杆菌可利用菊糖,而梭状芽孢杆菌、大肠杆菌和沙门氏菌等不能利用。在所测试的20种血清型沙门氏菌中,无一能在以菊糖为碳源的培养基上生长。菊糖被细菌代谢后,可提供短链脂肪酸作为黏膜细胞增殖的能源。短链脂肪酸是大肠黏膜细胞代谢的主要能源物质,大肠黏膜能源的70%来自肠细菌的发酵产物。而菊糖调节机体免疫系统主要是通过充当免疫刺激的辅助因子来提高抗体免疫应答的能力,从而提高动物机体体液及细胞免疫能力(Kelly 等,2012)。
4.2 在反刍动物生产中的应用 断奶羔羊日粮中添加15%菊芋粕,饲喂60 d,与对照组(基础日粮)相比,体质量增加30.6%,体高增加8.6%,体长增加16.3%,胸围增加12.60%。显著的生长育肥效果源于动物对饲料营养成分高效的吸收率。试验表明,小尾寒羊对菊芋粕DM消化率为62.47%,OM消化率为 65.18%,NDF消化率为48.2%,ADF消化率为46.3%,消化能达到13.22 MJ/kg(王水旺等,2014)。杨立杰等(2017)试验表明,在奶牛基础日粮中添加10%菊芋秸秆饲喂3周,可使乳脂率提高7.7%(P<0.05),血清GSHPx活性提高10.4%(P<0.05),乳汁中谷胱甘肽过氧化物酶(GSH-Px)和超氧化物歧化酶(SOD)活性分别提高18.7%和28.8%。奶牛日粮中粗料比例增加,有助于提高瘤胃pH,增加乙酸产量,进而提高乳脂率(Yves等,2007)。本实验室前期研究表明,奶牛日粮中添加0.8%菊芋块茎粉,可显著增加瘤胃乙酸和丙酸浓度,降低瘤胃NH3-N浓度,微生物蛋白(MCP)含量随饲喂时间的延长,有一定的上升趋势。同时,乳脂和乳糖含量显著增加,乳中多不饱和脂肪酸:棕榈油酸甲酯(C16:1cis-9)、 油酸甘油三酯 (C18:1cis-9)、 亚麻酸C18:3、花生四烯酸甘油三酯(C20:4)含量呈线性增加 (P<0.05)。奶牛瘤胃中乙酸和丁酸是合成乳脂肪的重要前体物,而丙酸不仅是提供能量的前体物,丙酸浓度增加也有助于提高乳蛋白含量,进而改善乳品质(Wilson,2017)。日粮中的脂质、瘤胃和乳腺中脂肪酸的转化均会影响牛奶中脂肪酸含量(Annika等,2006)。此前,对菊芋块茎营养成分的分析结果表明,菊芋块茎不含饱和脂肪酸,主要的脂肪成分由单不饱和多不饱和脂肪酸构成,主要是亚油酸(C18:2n-6)和亚麻酸(C18:3n-3)。此外,乳中脂肪酸含量与瘤胃微生物氢化活动有关(Jiang等,2015),并且通常以三酰甘油的形式从乳腺细胞转移到牛奶中。有研究证明,菊糖可显著提高奶牛瘤胃假单胞菌属、拟杆菌属、瘤胃球菌属、丁酸弧菌属的丰度,也可在一定程度上促进纤维降解菌属的增加(胡丹丹等,2017)。以前的研究表明,育肥犊牛在育肥后期会出现餐后高血糖、糖尿、高胰岛素血症和明显的胰岛素抵抗(Hugi等,1997),这些症状表明代谢控制不足或葡萄糖作为能源的低效使用。然而,也有证据表明,菊糖摄入可通过降低基础肝葡萄糖产生来降低餐后葡萄糖、尿素和甘油三酯浓度。其作用机制可能是可溶性纤维的补充增加了血浆胰高血糖素样肽-I的浓度,胰高血糖素样肽-I进而刺激胰岛素分泌(Kaufhold 等,2000)。
5 总结
菊芋种植成本低、附加值高又适于规模化加工,通过定量化育种理论与灰色关联度分析方法进行系统、客观地品系(种)选育可实现牧草型菊芋种植与畜牧饲养发展相衔接。牧草型菊芋秸秆的营养价值高于一般粗饲料,块茎中丰富的菊糖不仅是人类理想的绿色营养食品,也可为畜禽提供保健、减抗等功效的优质饲料添加剂。菊芋秸秆作为新成员进入国家饲料数据库,将为缓解我国优质粗饲料资源短缺、促进畜牧养殖业的发展,提供新的饲料供给途径,有可能迈向 “小菊芋大产业”的未来。然而,值得注意的是菊芋不同品种之间营养成分存在较大的差异。不同刈割时期、不同植株部位(地上部分、地下部分)、不同状态(新鲜、风干)以及不同的贮藏方式等都会对评估菊芋的营养成分及饲用效果造成较大的影响。此外,菊芋饲料在动物生产(特别是反刍动物)中的应用仍有较大的探索空间。例如,菊芋青贮饲料的营养成分的测定及饲用价值;菊芋秸秆及块茎在瘤胃中的降解代谢以及在生产中适宜添加量的确定;菊芋中生物活性物质对动物抗病作用的分子机制研究等。综上,尽管菊芋对农业、畜牧业甚至医学都表现出多种潜在利用价值,但仍需进一步研究以更全面地了解和利用这种多功能作物,为新型饲料资源的开发利用以及为动物的健康水平提高和生产提供一定的帮助。
