gD糖蛋白基因表达质粒pcDNA3Kan-gD的构建及其在Hela细胞中的表达检测
2020-11-12何卓晶周寒鹏
何卓晶,陶 薇,傅 婷,周寒鹏,洪 艳
(杭州医学院 生物工程研究所,浙江 杭州 310013)
2型单纯疱疹病毒(herpes simplex virus type 2, HSV-2)感染可引起皮肤水泡、溃疡性病变及神经系统和泌尿系统的并发症,易传播给易感的性伴侣、胎儿和新生儿[1],可增加感染人类免疫缺陷病毒的危险[2-3]。HSV-2感染导致的生殖器疱疹是引起生殖器区域溃疡的原因之一,可引起相关的公共健康问题[4-5]。HSV-2初次感染期间,病毒即在骶神经节中终身潜伏下来,并能再次激活导致复发。目前,生殖器疱疹的标准治疗和预防药物为鸟苷类似物,如阿昔洛韦,伐昔洛韦,喷昔洛韦和泛昔洛韦。鸟苷类似物药物能通过干扰HSV的DNA聚合酶而抑制病毒复制,但该类药物需每日给药以维持其抑制病毒复制的作用,且很多HSV毒株对该类抗病毒药物产生了抗性[6]。疫苗是控制生殖器疱疹的另一有效策略,HSV-2病毒衣壳上的糖蛋白D(glycoprotein D, gD)具有较高的免疫原性,近年来逐渐成为单纯疱疹病毒亚单位疫苗的研究热点。本研究构建gD糖蛋白基因表达质粒pcDNA3Kan-gD,并观察其在Hela细胞中的表达情况,旨在为单纯疱疹病毒亚单位疫苗的研究提供基础。
1 材料与方法
1.1 主要材料与试剂 质粒pcDNA3Kan为本实验室构建的含卡那霉素抗性的真核表达质粒载体,pcDNA3Kan-gD为本实验室保存的含有HSV-2糖蛋白gD基因的质粒。Hela细胞、BALB/c小鼠抗HSV-2血清均为本单位生物工程所制备及保存。RNA抽提试剂盒购自杭州诺扬生物技术有限公司,限制性内切酶、T4DNA连接酶、高保真DNA聚合酶均购自Promage公司,质粒转染SuperFectinTMⅡ in Vrio DNA transfection Reagent试剂盒购自上海普飞生物技术有限公司,高纯度质粒小提试剂盒、Goat Anti-Mouse IgG DylightTM448及Hochest 33258购自康为世纪生物科技有限公司。所有引物由上海生工生物工程技术服务有限公司合成。
1.2 真核重组表达质粒 pcDNA3Kan-gD的构建根据HSV-2gD基因的编码序列以及质粒载体pcDNA3kan的酶切位点设计引物,在上下游引物的两端分别添加EcoRI和XhoI酶切位点,上游引物P1:5’ATCGAATTCAACCACTAGTCGCCG 3’;下游引物P2:5’CGCTCGAGACTCCCTTTATGC 3’。从pcDNA3Kan-gD上通过PCR扩增获取gD全长序列基因片断,大小为420 bp。然后用EcoRI和XhoI分别对真核表达质粒载体pcDNA3Kan及PCR扩增得到的gD基因片断进行双酶切。将酶切产物用T4 DNA连接酶进行连接,经转化、克隆,获得真核重组表达质粒pcDNA3Kan-gD。
1.3 质粒的提取与纯化 将含真核重组表达质粒pcDNA3Kan-gD的E.coliBL21菌株和含pcDNA3Kan的E.coliBL21菌株分别于含卡那霉素(25 μg/mL)的LB培养基中培养过夜,次日使用高纯度质粒小提试剂盒抽提并纯化pcDNA3Kan质粒和真核重组表达质粒pcDNA3Kan-gD,核酸蛋白测定仪测定其浓度和A260/A280。
1.4 细胞培养 Hela细胞在含10%胎牛血清的高糖DMEM培养基中,于37℃ 5% CO2条件下培养。
1.5 质粒转染 Hela细胞按照质粒转染试剂盒的使用说明进行:于转染前一天将Hela细胞转入12孔细胞培养板中传代培养一次,待细胞长至70%~80%融合度且生长良好时进行转染。转染前更换新鲜的完全培养基并孵育30~60 min。于1.5 mL无菌离心管中准备以下溶液,溶液A:用不含血清的高糖DMEM培养基分别将0.188 μg、0.375 μg和0.750 μg(即1×、2×和4×)真核重组表达质粒pcDNA3Kan-gD稀释至38 μL;溶液B:用不含血清的高糖DMEM培养基将2.25 μL SuperFectinTMⅡ试剂稀释至38 μL;将溶液B加入到溶液A中并混匀,室温孵育15 min。将SuperFectinTMⅡ/DNA混合物加入细胞培养物,轻轻摇动培养板以混匀。5%CO2,37℃培养12 h后,更换新鲜的完全培养基。继续培养24 h后,采用半定量RT-PCR技术及免疫荧光技术检测梯度转染的pcDNA3Kan-gD重组质粒在Hela细胞中的表达情况。
1.6 半定量RT-PCR 倾去转染后Hela细胞的培养上清,用0.25%的胰蛋白酶消化后收集细胞。按RNA抽提试剂盒使用说明提取细胞总RNA,提取的RNA溶于DEPC处理水中。按cDNA第一链合成试剂盒使用说明完成逆转录反应并合成cDNA第一链后进行PCR反应。反应条件为:94 ℃预变性2 min;94 ℃变性30 s,57 ℃退火30 s,72 ℃延伸30 s,28个循环;72 ℃延伸2 min。上游引物:5’GCGTGTTTACCACATTCAGCC 3’;下游引物:5’TCCGTCCAGTCGTTTATCTTCA 3’。β-actin的扩增反应条件为:94 ℃预变性2 min;94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸30 s,28个循环;72 ℃延伸2 min。上游引物:5’ATCATGTTTGAGACCTTCAACA 3’;下游引物:5’CATCTCTTGCTCGAAGTCCA 3’。扩增产物用1%的琼脂糖凝胶电泳鉴定,并采用ImageJ软件分析条带的灰度值。
1.7 免疫荧光技术 消化梯度转染(1×和4×)pcDNA3Kan-gD重组质粒的Hela细胞进入12孔板,待细胞贴壁,用PBS清洗3次,4%多聚甲醇固定30 min,PBS清洗;于37 ℃用含0.3% Triton的PBS处理30 min,PBS清洗3次;用含1%BSA的PBS封闭30 min后,PBS清洗1次;以BALB/c小鼠抗HSV-2血清(1∶100稀释)作为一抗,4 ℃孵育过夜,次日PBS清洗;以Goat Anti-Mouse IgG DylightTM488作为荧光二抗,37 ℃孵育1 h,PBS清洗;加入Hochest 33258染液染细胞核10 min,PBS清洗。采用荧光倒置显微镜(CKX41,日本OLYMPUS)观察结果。
2 结果
2.1 真核重组表达质粒pcDNA3Kan-gD的鉴定 将构建完成的真核重组表达质粒pcDNA3Kan-gD以限制性内切酶EcoRI和XhoI双酶切,产物用1%的琼脂糖凝胶电泳鉴定,在420 bp处显示一条带,大小与预期相符,见图1。
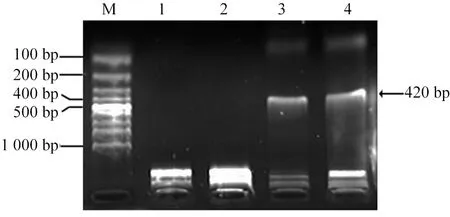
M:标准分子量DNA;1:真核重组表达质粒pcDNA3Kan-gD;2:真核表达质粒载体pcDNA3Kan;3:通过PCR扩增获取的gD全长序列基因片断;4:pcDNA3Kan-gD的EcoR I和Xho I双酶切片段。图1 真核重组表达质粒pcDNA3Kan-gD的鉴定结果
2.2 质粒提纯 将纯化的质粒用无菌蒸馏水定容至0.5 mL后,测定其浓度和纯度。纯化后的pcDNA3Kan-gD的浓度为0.16 μg/μL, A260/A280为1.703;纯化后的pcDNA3Kan的浓度为0.14 μg/μL,A260/A280为1.718。
2.3 重组质粒转染后半定量RT-PCR结果 转染pcDNA3Kan-gD的Hela细胞总RNA经RT-PCR后扩增出大小为420 bp的gD基因条带,β-actin基因条带大小为318 bp;分析条带灰度值显示,转染质粒的相对表达量(转染质粒与β-actin表达量的比值)与转染倍数呈剂量依赖关系;见图2。

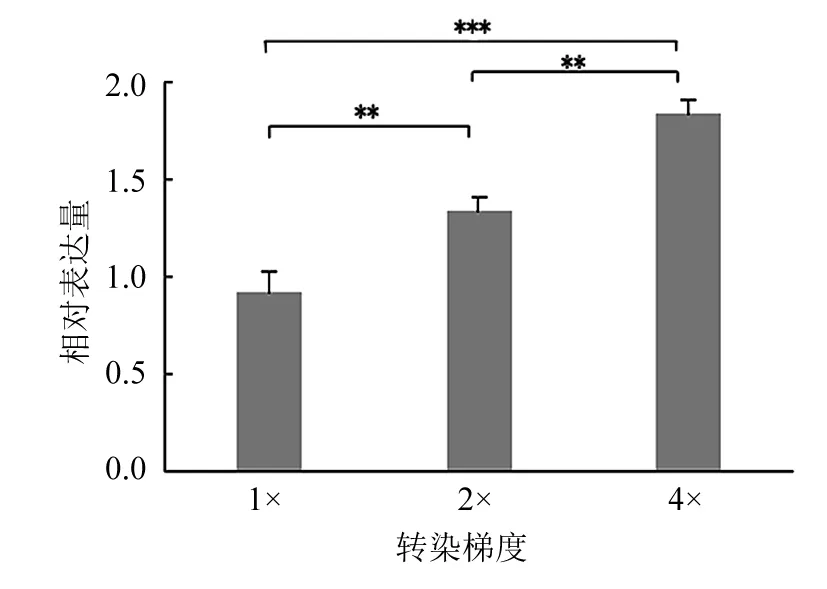
M:标准分子量DNA;1:gD基因片段(转染量为0.188 μg,1×);2:gD基因片段(转染量为0.375 μg,2×);3:gD基因片段(转染量为0.750 μg,4×);4:β-actin基因片段(转染量为0.188 μg,1×);5:β-actin基因片段(转染量为0.375 μg,2×);6:β-actin基因片段(转染量为0.750 μg,4×);**P<0.01; *** P<0.001。图2 重组质粒转染后半定量RT-PCR结果
2.4 免疫荧光技术分析结果 转染pcDNA3Kan-gD的Hela细胞呈现绿色荧光,1×与4×转染倍数的Hela细胞平均光密度比较,差异有统计学意义(P<0.001),见图3。
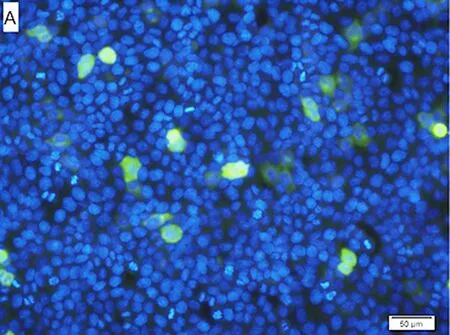

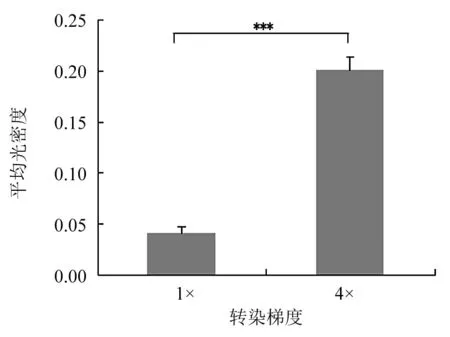
A:转染1× pcDNA3Kan-gD的Hela细胞;B: 转染4× pcDNA3Kan-gD的Hela细胞;***P<0.001。图3 转染重组质粒的Hela细胞免疫荧光分析结果
3 讨论
DNA疫苗通常由DNA质粒构建而成。当DNA质粒转染进入疫苗接种者后表达蛋白抗原,由此介导外源蛋白的内源性生产,包括其天然构造和转录后适当的修饰。早期研究[7]结果认为,向哺乳动物肌肉中直接注射携带真核基因的质粒DNA,能诱导编码蛋白的内源性表达及抗编码蛋白的特异性免疫反应,由此奠定了DNA疫苗的发展基础。这种以DNA为基础的免疫实验已经发展为保护人类和动物健康,抗击感染性疾病、癌症或过敏症极具前景的手段。DNA疫苗优点在于其安全性高(使用的质粒在真核细胞中是非复制性的),生产系统简单(大肠杆菌的扩增和纯化不复杂且相对廉价),能刺激强有力的细胞免疫反应(由转染细胞引起的MHC I介导的抗原递呈),具有组合疫苗的可能性(仅需混合不同的DNA分子[8]),无需冷冻运输装置(DNA的高度稳定性)。
早期的临床试验[9]显示,以单纯疱疹病毒gD糖蛋白为亚基的疫苗是安全的,且在人体中能诱导病毒中和抗体。Stanberr等[10]观察到适度的疫苗效力能对血清反应阴性的个体起到保护作用。gD是单纯疱疹病毒的一个基本糖蛋白,在病毒的侵入过程中有两个主要作用:绑定两个主要的细胞受体(HVEM或nectin-1);激活病毒融合装置的下游组分,gH/gL和gB[11]。在gD上至少有5个特异性的抗原表位能诱导病毒中和抗体。本研究以gD糖蛋白基因为基础,构建含有gD全长序列的真核重组表达质粒,并观察其在Hela细胞中的表达情况。
质粒载体pcDNA3具有高度的稳定性并能在大多数哺乳动物细胞中瞬时表达,而其抗性选择基因氨苄青霉素基因可能对人体存在潜在的威胁。本研究对含Amp抗性基因的真核表达质粒载体pcDNA3进行重组,以Kan抗性基因替换Amp抗性基因,构建了含Kan抗性的真核表达质粒载体pcDNA3Kan-gD,为该DNA疫苗更适应于人体奠定了基础。
综上所述,含有单纯疱疹病毒gD糖蛋白全长序列的真核表达质粒pcDNA3Kan-gD能在Hela细胞中瞬时表达gD蛋白,有可能成为抗单纯疱疹病毒DNA疫苗候选物之一。
