外源蛋白合成抑制剂对杂交水稻种子耐贮藏能力的影响
2020-11-10杨天姝高家东戴彰言贝锦龙张琪陈中健刘军付华陈光辉
杨天姝,高家东,戴彰言,贝锦龙,张琪,陈中健,刘军,付华,陈光辉
外源蛋白合成抑制剂对杂交水稻种子耐贮藏能力的影响
杨天姝1,2,3,高家东2,3#,戴彰言2,3#,贝锦龙2,3,张琪2,3,陈中健2,3,刘军2,3,付华4*,陈光辉1*
(1.湖南农业大学农学院,湖南 长沙 410128;2.广东省农作物种质资源保存与利用重点实验室广东 广州 510640;3.广东省农业科学院农业生物基因研究中心,广东 广州 510640;4.广东省农业科学院水稻研究所,广东 广州 510640)
以不耐贮藏杂交水稻II优998种子及博优998(对照)种子为材料,采用iTRAQ定量蛋白质组学的方法,鉴定2种不同耐贮藏能力种子贮藏前后的差异蛋白,共鉴定出5 210个蛋白,其中差异显著的蛋白100个;II优998种子在贮藏期间的差异蛋白数量显著高于对照种子的差异蛋白数;通过Pathway显著性富集的方法,确定差异蛋白参与的主要代谢途径是Ribosome途径;II优998种子在自然条件贮藏期间有大量的核糖体蛋白变化差异,表明蛋白质生物合成可能与种子耐贮藏能力有关。灌浆期利用3种外源蛋白合成抑制剂(农用链霉素、春雷霉素和广谱性农药咪鲜胺)喷施种子,结果表明,喷施农用链霉素可以提高杂交水稻种子自然条件下的耐贮藏能力。
水稻;种子;耐贮藏能力;核糖体蛋白;外源蛋白合成抑制剂;发芽率
种子是遗传物质的载体,也是植物种质资源长期保存和物种多样性保存的重要材料。由于种子生产的不确定性,常需要对种子进行短期贮藏,但水稻种子在南方高温高湿条件下容易劣变而丧失活力,对农业生产造成巨大损失[1-2]。
种子耐贮藏能力受遗传、种子发育期间的环境条件及贮藏条件等因素影响[1-6]。长期以来,国内外主要通过工程技术途径(如低温低湿技术)解决种子安全贮藏问题,但即使使用低温冷库,也无法从根本上解决种子老化劣变引起的种子活力下降与寿命丧失问题[7];因此,了解相应的调控机制,寻找更有效的调控方法,提升种子的耐贮藏能力已成为研究的热点。
本研究中,采用iTRAQ定量蛋白质组学技术获得差异蛋白,探索蛋白质的生物合成与杂交稻种子耐贮藏能力的关系,旨在筛选能够有效延长杂交稻种子耐贮藏能力的外源调控产品。
1 材料与方法
1.1 材料
以不耐贮藏的杂交稻II优998种子为材料,以生产中大面积应用的相对耐贮藏的博优998种子为对照。2种材料均来自广东省金稻种业有限公司。
1.2 方法
1.2.1种子胚iTRAQ定量蛋白质组学分析
首先参照GAO等[2]的方法提取种子胚蛋白质,然后开展蛋白质酶解和质谱检测。
将保存于冰箱的蛋白质干粉溶于适量裂解缓冲液(7 mol/L尿素、2 mol/L硫脲、4%CHAPS、40 mmol/L Tris-HCl,pH 8.5),200 W超声15 min。4 ℃ 30 000离心20 min,得上清液;用Bradford试剂盒测量蛋白浓度。加入DTT,使其浓度为10 mmol/L,于37 ℃保持4 h;冷却至室温,加入0.55 mol/L的碘乙酰胺,使其浓度为55 mmol/L,避光反应1 h;再加入5倍体积的冷丙酮,于-20 ℃过夜,沉降蛋白,4 ℃ 8 000离心20 min;收集沉淀,干燥后,加入约50 µL TEAB溶解蛋白,超声15 min;加入胰蛋白酶Trypsin Gold(酶与蛋白的质量比为1∶20),37 ℃酶解,过夜。
参照YANG等[8]的方法进行iTRAQ定量蛋白质组学分析。
采用Applied Biosystems的4-plex iTRAQ分别标记酶解后的肽段,将少量标记样品混合,用质谱仪(Thermo Orbitrap Fusion)测定肽段标记的效率。标记检测无异常后,再将全部的标记样品等量混合。参照说明书,通过C18SPE柱(C18Cartridge Solid Phase Extraction)去除已标记肽段混合液中的盐离子。利用Thermo RSLC 3000仪器结合XterraMs C18柱(3.5 µm, 2.1 mm×150 mm, Waters)对混合后的肽段进行预分离,分级为12个组分。对每个组分进行液相串联质谱(LC−MS/MS)分析,基本条件如下:高效液相色谱仪,Thermo Scientific RSLC Nano 3000;富集柱,ThermoPepMap u-precolumn(5 µm, 300 µm×5 mm);分析柱,PepMap C18反相纳升柱(3 µm, 75 µm×150 mm, Dionex);主要有机相梯度,4%~90%乙腈(含0.1%甲酸)洗脱135 min;流速,300 nL/min;上样量,1 µg;质谱仪,ThermoOrbitrap Fusion;喷雾电压,1.9 kV;离子传输管温度,275 ℃;扫描模式,正离子;碰撞能量,40% HCD;分辨率设置,一级120 000(/200),二级30 000(/200);母离子扫描范围:/350~1550;子离子扫描范围,自动。对质谱下机的原始文件进行峰识别,得到峰列表。利用蛋白质数据库检索软件MASCOT进行肽段及蛋白质的鉴定,比较各蛋白在各样品之间相对含量的关系,从而获得一些重要的蛋白。
样品编号:贮藏前博优998种子用S1表示,贮藏后博优998种子用S2表示;贮藏前Ⅱ优998种子用S3表示,贮藏后Ⅱ优998种子用S4表示。每个样本3次重复。
1.2.2田间制种及种子寿命调控剂处理
参照吴玉坤等[9]的方法进行田间制种,灌浆期适时喷施多效唑。在种子收获前12 d或喷施“九二〇”后7 d,分别喷施外源蛋白质合成抑制剂农用链霉素、春雷霉素和广谱性农药咪鲜胺等调控剂。以喷施清水为对照。
1.2.3种子人工老化处理
参照刘军等[10]和CHEN等[11]的方法进行种子人工老化处理。
老化处理前准备3个密封的干燥器,其中1个加入过饱和MgCl2溶液,置于20 ℃人工气候箱或者烘箱,其余2个各加入过饱和的KCl溶液,分别置于25 ℃和40 ℃人工气候箱或者烘箱。将收获的水稻种子分别装于网袋,放于密封干燥器(加入过饱和的KCl溶液,25 ℃,相对湿度85%),预处理3 d;转移至已经平衡好的干燥器中(加入过饱和的KCl溶液,40 ℃,相对湿度83%)处理8 d;再置于干燥器(加入过饱和MgCl2溶液,20 ℃,相对湿度33%)干燥3 d,密封,贮藏于-20 ℃的冰箱,备用。
1.2.4种子自然老化处理
参照GAO等[2]的方法,将收获的水稻种子分别用网袋装好,于自然条件贮藏2年后取样,测定发芽率。
1.2.5种子发芽率的测定
称取干燥后的种子,每个处理15 g,充分吸水12 h。倒掉多余水分后,将种子置于底部垫有2层发芽纸的发芽皿中进行萌发(每盒发芽皿100粒)。萌发时采用15 000 lx光照培养箱(12 h光照,12 h黑暗),保持发芽纸湿润且无积水。统计萌发7 d的发芽率。3次重复。
1.3 数据处理
采用Perseus软件进行差异蛋白数据处理与统计分析[12]。根据蛋白丰度差异显著性筛选出不同类型的差异蛋白,当蛋白丰度差异倍数达到2倍以上,且<0.05时,视为差异蛋白。
运用KOBAS 2.0进行KEGG(Kyoto Encyclopedia of Genes and Genomes)通路(pathway)富集分析[13]。以所有鉴定到的蛋白为背景,对不同类型的差异蛋白进行KEGG富集度分析,采用KOBAS的富集模块计算不同KEGG通路的富集情况。
采用Excel 2007进行数据处理与分析;运用Sigma Plot绘图。
2 结果与分析
2.1 不同种子贮藏前后的差异蛋白质鉴定
利用MASCOT共鉴定出二级谱图545 722个,特有肽段序列74 651个,5 210个蛋白质(表1)。使用Perseus软件进行统计分析,共鉴定出100个差异表达蛋白质(DEP)。其中,不同比较方案的平均差异蛋白分别为19、29.75、23和28.33种(表2)。从表2可以看出,Ⅱ优998贮藏前后(S3与S4)2倍差异以上的蛋白平均为28.33个(4个下调,24.33个上调);对照博优998贮藏前后(S1与S2)2倍差异以上的蛋白19个(平均7.67个上调,11.33个下调)。不同耐贮藏能力种子贮藏前后的差异蛋白质变化较大。贮藏前S1与S3的差异蛋白为29.75个,其中S3中丰度高的差异蛋白质仅有2.75个,而对照S1中丰度高的差异蛋白较多,为27个。贮藏后比较S2与S4,与贮藏前的结果相反,不耐贮藏的II优998老化种子中丰度高的差异蛋白为19.33个,多于S2中的。

表1 不同水稻种子贮藏前后蛋白质组iTRAQ鉴定结果

表2 差异蛋白数量统计
表中数据为3次重复的平均值。
2.2 不同种子贮藏后的差异蛋白质比较
采用KEGG通路(Pathway)显著性富集分析上述差异表达蛋白参与的最主要代谢途径和信号转导途径。在所有的比较类型中,3种比较(S1与S3、S2与S4、S3与S4)都仅有1条通路具有统计学意义,且均为Ribosome途径,而S1与S2的比较也是Ribosome途径排名第一。
表3列出了Ⅱ优998种子在贮藏前后(S3与S4)出现的代表性差异蛋白。从表3可以看出,在II优998种子中,丰度较高的蛋白绝大部分是核糖体蛋白,而这些核糖体蛋白的绝大部分(87%)是贮藏后种子 (S4)中丰度高的差异蛋白。说明Ⅱ优998种子在自然老化过程中大量的核糖体蛋白丰度上调,有较多的蛋白质合成代谢。

表3 Ⅱ优998种子在贮藏前后出现的代表性差异蛋白
2.3 灌浆期外源调控剂对人工老化种子发芽率的影响
从图1可以看出,种子入库时,不同外源调控剂处理的种子发芽率都为96%左右,处理与对照无显著差异,表明各调控剂处理对种子初始发芽率的影响不明显。
人工老化8 d后,外源蛋白质合成抑制剂春雷霉素处理的种子发芽率可达80%,显著高于对照的发芽率,比对照的高5.65%,表明春雷霉素能够提高种子人工老化后的出芽能力。与对照相比,农用链霉素处理的发芽率略有增加,咪酰胺处理的发芽率有所下降,但均与对照无显著差异。

图1 不同处理种子人工老化前后的发芽率
2.4 灌浆期外源调控剂处理对自然老化种子发芽率的影响
从图2可以看出,自然贮藏2年后,农用链霉素处理的种子的发芽率(76.25%)极显著高于对照处理的(66.79%);春雷霉素处理的种子发芽率与对照无显著差异;咪酰胺处理的种子的发芽率仅为43.50%,极显著低于对照种子的发芽率。表明农用链霉素处理可以有效提高种子的耐贮藏能力。
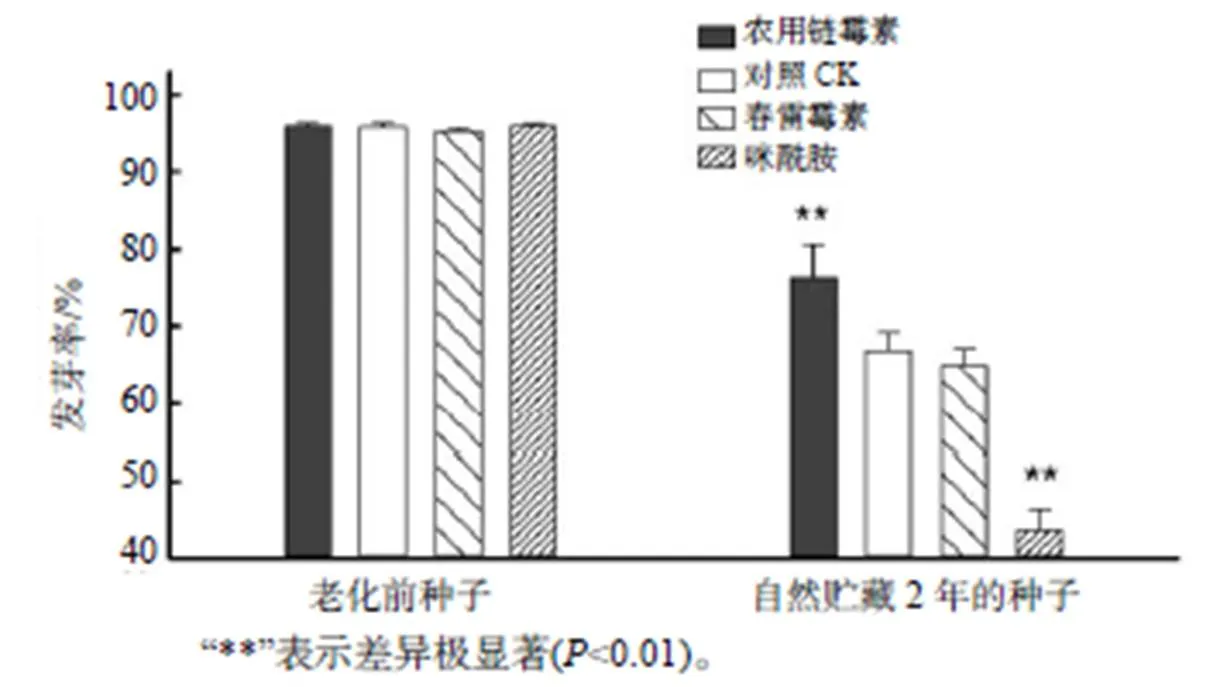
图2 不同处理种子自然老化前后的发芽率
3 结论与讨论
为探索水稻种子劣变过程中响应各种胁迫的调控机制,本课题组前期选取不同耐贮藏能力的杂交稻种子(用自然贮藏2年后种子发芽率衡量种子的耐贮藏能力),利用基于双向电泳的蛋白质组学方法对种胚的蛋白质差异进行比较,发现与自然贮藏老化过程种子耐贮藏有关的蛋白包括氧化还原调节蛋白(如谷氧还蛋白,乙二醛酶等)、DNA损伤修复蛋白以及Lea 蛋白等,且不耐贮藏种子在贮藏过程中的差异蛋白数量远远高于相对耐贮藏种子的[2]。本研究采用iTRAQ技术鉴定出5 200多个总蛋白,同样发现种子自然贮藏过程中不耐贮藏种子的差异蛋白数高于耐贮藏种子中的数量,2种方法鉴定结果基本一致。
本研究中,发现不耐贮藏种子在贮藏过程中有核糖体蛋白的剧烈变化,表明蛋白质的生物合成可能与种子贮藏寿命有关。基于蛋白质翻译合成可能与种子贮藏寿命有关的结果,推测不耐贮藏种子在老化期间可能有较多的蛋白质合成。本研究中,通过喷施外源蛋白合成抑制剂来调控水稻种子的耐贮藏能力,发现农用链霉素能够提高自然贮藏2年的种子的发芽率,即提高了种子的耐贮藏能力,但其作用机理还需进一步研究。
[1] 傅家瑞.种子生理[M].北京:科学出版社,1985. FU J R.Seed Physiology[M].Beijing:Science Press, 1985.
[2] GAO J D,FU H,ZHOU X Q,et al.Comparative proteomic analysis of seed embryo proteins associated with seed storability in rice(L.) during natural aging[J].Plant Physiology and Biochemistry,2016,103:31-44.
[3] VENTURA L,DONÀ M,MACOVEI A,et al. Understanding the molecular pathways associated with seed vigor[J].Plant Physiology and Biochemistry,2012,60:196-206.
[4] WALTERS C,WHEELER L M,GROTENHUIS J M. Longevity of seeds stored in a genebank:species characteristics[J].Seed Science Research,2005,15(1):1-20.
[5] YAN S J,HUANG W J,GAO J D,et al.Comparative metabolomic analysis of seed metabolites associated with seed storability in rice(L.)during natural aging[J].Plant Physiology and Biochemistry,2018,127:590-598.
[6] 杨亚平, 姜孝成, 陈良碧, 等. 水稻种子老化的生理机制[J]. 湖南农业大学学报(自然科学版),2008,34(3):265-269. YANG Y P,JIANG X C,CHEN L B,et al. Study on physiological mechanism in aging of rice seeds[J]. Journal of Hunan Agricultural University(Natural Sciences),2008,34(3):265-269.
[7] 卢新雄,辛霞,尹广鹍,等.中国作物种质资源安全保存理论与实践[J].植物遗传资源学报,2019,20(1):1-10. LU X X,XIN X,YIN G K,et al.Theory and practice of the safe conservation of crop germplasm resources in China[J].Journal of Plant Genetic Resources,2019,20(1):1-10.
[8] YANG Q S,WU J H,LI C Y,et al.Quantitative proteomic analysis reveals that antioxidation mechanisms contribute to cold tolerance in plantain(L.;ABB Group) seedlings[J].Molecular & Cellular Proteomics,2012,11(12):1853-1869.
[9] 吴玉坤,陈国荣,黄健文,等.优质高产杂交稻新组合天优998制种技术[J].杂交水稻,2005,20(1):31-32. WU Y K,CHEN G R,HUANG J W,et al.Seed production techniques of Tianyou 998,a new good quality and high yield hybrid rice combination[J].Hybrid Rice,2005,20(1):31-32.
[10] 刘军,黄上志,傅家瑞.不同活力玉米种子胚萌发期间热激蛋白的合成[J].植物学报,2000,42(3):253-257. LIU J,HUANG S Z,FU J R.Synthesis of heat shock protein in maize seed embryo with different vigor during germination[J].Acta Botanica Sinica,2000,42(3):253-257.
[11] CHEN H H,CHU P,ZHOU Y L,et al.Overexpression of AtOGG1,a DNA glycosylase/AP lyase,enhances seed longevity and abiotic stress tolerance in[J]. Journal of Experimental Botany,2012,63(11):4107- 4121.
[12] TYANOVA S,COX J.Perseus:a bioinformatics platform for integrative analysis of proteomics data in cancer research[M]//VON STECHOW L.Cancer Systems Biology.Methods in Molecular Biology.New York:Humana Press,2018:133-148.
[13] XIE C,MAO X Z,HUANG J J,et al.KOBAS 2.0:a web server for annotation and identification of enriched pathways and diseases[J].Nucleic Acids Research,2011,39(suppl 2):W316-W322.
Effects of exogenous protein synthesis inhibitors on hybrid rice seed storability
YANG Tianshu1,2,3, GAO Jiadong2,3#, DAI Zhangyan2,3#, BEI Jinglong2,3, ZHANG Qi2,3, CHEN Zhongjian2,3, LIU Jun2,3, FU Hua4*, CHEN Guanghui1*
(1.College of Agronomy, Hunan Agricultural University, Changsha, Hunan 410128, China;2.Guangdong Provincial Key Laboratory for Crop Germplasm Resources Preservation and Utilization, Guangzhou, Guangdong 510640, China; 3.Agro-biological Gene Research Center, Guangdong Academy of Agricultural Sciences, Guangzhou, Guangdong 510640, China; 4.Rice Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou, Guangdong 510640, China)
In this study, a quantitative proteomics of iTRAQ was conducted for rice seeds before and after 24-month natural storage on two hybrid rice cultivars, II You 998 with low storability and the control Boyou 998. More than 5 210 proteins were identified, of which 100 differentially expressed protein (DEP) were significant different. Interestingly, the amount of differentially expressed proteins in the storage period of the seeds with poor storability was significantly higher than that of the control seeds. Furthermore, ribosome pathway was determined to be the main metabolic pathway involved in regulation of seed storability by pathway significant enrichment. Our data showed that more ribosomal proteins were synthesized in the seeds which were not resistant to storage, suggesting that protein biosynthesis might be related to the storage tolerance of seeds. And, the germination percentages of hybrid rice seeds treated with different exogenous protein synthesis inhibitors during the filling stage significantly varied after 2 years storage under natural conditions. Our study showed that treatment of agro-streptomycin, during the filling stage could effectively enhance rice seed storability under natural storage conditions.
; seed; storability; ribosomal protein; exogenous protein synthesis inhibitors; germination percentage
S511.093
A
1007-1032(2020)05-0501-06
杨天姝,高家东,戴彰言,贝锦龙,张琪,陈中健,刘军,付华,陈光辉.外源蛋白合成抑制剂对杂交水稻种子耐贮藏能力的影响[J].湖南农业大学学报(自然科学版),2020,46(5):501-506.
YANG T S, GAO J D, DAI Z Y, BEI J L, ZHANG Q, CHEN Z J, LIU J, FU H, CHEN G H. Effects of exogenous protein synthesis inhibitors on hybrid rice seed storability[J]. Journal of Hunan Agricultural University(Natural Sciences), 2020, 46(5): 501-506.
http://xb.hunau.edu.cn

2019-11-04
2019-12-23
国家自然科学基金项目(31871716、31371715);广东省科技项目(2018B020202004、2019KJ106);广州市科技项目(201909020001、201807010114)
杨天姝(1993—),女,黑龙江哈尔滨人,硕士研究生,主要从事种子科学与技术研究,1290989730@qq.com;#共同第一作者,高家东(1977—),男,湖南永州人,助理研究员,主要从事种子生物学研究,gaogao126@139.com;#共同第一作者,戴彰言(1985—),男,广东广州人,助理研究员,主要从事生物信息学研究,daizhangyan@agrogene.ac.cn;*通信作者,付华,副研究员,主要从事水稻分子育种研究,13825164071@139.com;*通信作者,陈光辉,博士,教授,主要从事种子科学与技术研究,cgh68@163.com
责任编辑:毛友纯
英文编辑:柳正
