三穗鸭IP3R3基因SNP位点鉴定及其对蛋壳品质的影响
2020-11-09李杰章谭光辉吴磊覃媛钰张依裕
李杰章 谭光辉 吴磊 覃媛钰 张依裕
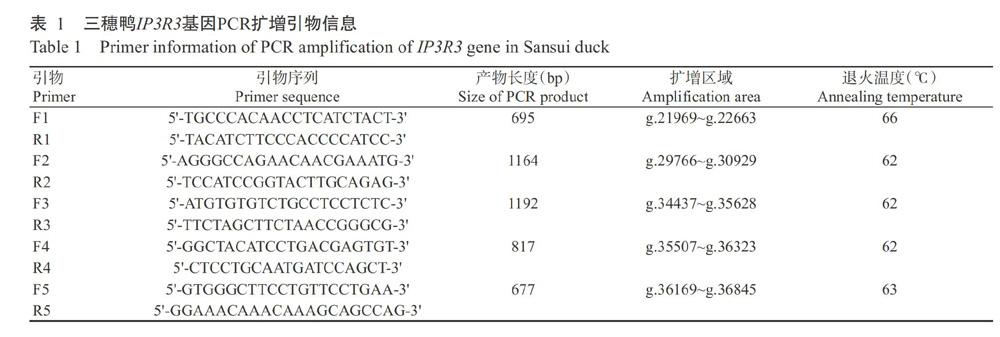
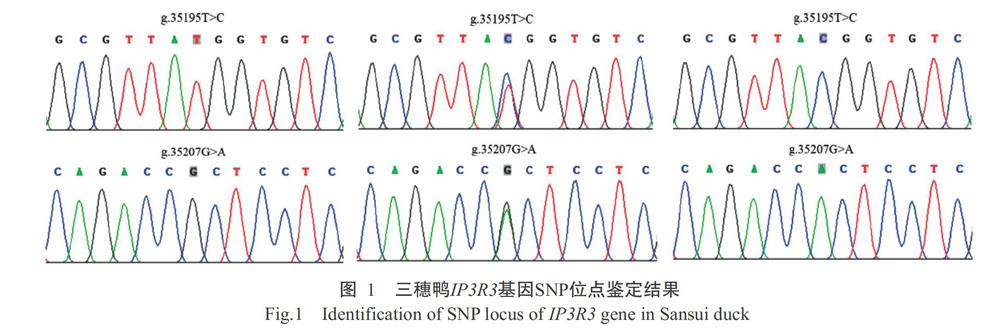
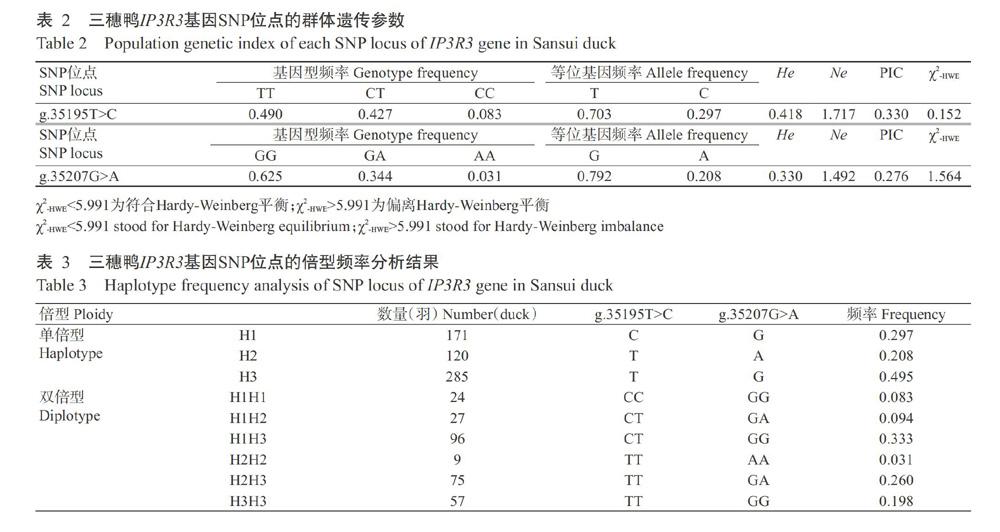
摘要:【目的】明確IP3R3对蛋壳性状的效应机制及其在蛋壳品质改良中的应用价值,为开展蛋壳品质改良分子标记育种提供参考依据,也为深入研究IP3R3基因的生物学功能打下基础。【方法】以288羽三穗鸭为研究对象,收集45周龄母鸭生产的鸭蛋,测定蛋重、蛋形指数、蛋壳厚度、蛋壳强度和蛋壳重;采用DNA直接测序法对三穗鸭IP3R3基因SNP位点进行鉴定,使用SHEsis进行单倍型及连锁不平衡分析,通过RNAfold预测不同单倍型的mRNA二级结构和自由能,并以SPSS 18.0中的一般线性模型(GLM)对三穗鸭IP3R3基因SNP位点与蛋壳品质进行关联分析。【结果】在三穗鸭IP3R3基因外显子上发现2个SNPs位点,分别位于第49外显子的g.35195T>C和g.35207G>A,均属于同义突变,且均存在3种基因型,对应的多肽信息含量(PIC)分别为0.330和0.276,属于中度多态位点(0.25
关键词: 三穗鸭;IP3R3基因;SNP位点;蛋壳品质;关联性
中图分类号: S834.89 文献标志码: A 文章编号:2095-1191(2020)07-1729-08
Abstract:【Objective】To clarify the effect mechanism of IP3R3 on eggshell traits and its application value in eggshell quality improvement, provide reference basis for molecular marker breeding of eggshell quality improvement and lay a foundation for further research on the biological function of IP3R3 gene. 【Method】Taking 288 Sansui ducks as the research object, duck eggs produced by 45-week old female ducks were collected, and egg weight, egg shape index, eggshell thickness, eggshell strength and eggshell weight were measured. The SNP sites of Sansui duck IP3R3 gene were identified by DNA direct sequencing, and haplotype and linkage disequilibrium analysis were performed by SHEsis. The secondary structure and free energy of different haplotypes mRNA were predicted by RNAfold, and the correlation analysis between SNP sites of Sansui duck IP3R3 gene and eggshell quality was conducted by general linear model(GLM) in SPSS 18.0. 【Result】Two SNPs sites were found on the IP3R3 exon of Sansui duck, respectively located at g.35195T>C and g.35207G>A of exon 49, both of which were synonymous mutations, and there were three genotypes. The corresponding polypeptide information contents(PIC) were 0.330 and 0.276, respectively, belonging to moderate polymorphism sites (0.25
Key words: Sansui duck; IP3R3 gene; SNP locus; eggshell quality; correlation
Foundation item: National Natural Science Foundation of China(31760663); Guizhou Outstanding Young Scientific and Technological Talents Subsidy Project(QKHPTRC〔2017〕5788); Guizhou Province Thousand-level Innovative Ta-lents Project(701030174401)
0 引言
【研究意义】我国是典型的农业大国,畜禽养殖在国民经济中占据重要地位,而家禽养殖业又是畜牧产业的重要组成部分。禽蛋是人类重要的食品之一,由蛋壳、蛋黄、蛋白和蛋系带等部分组成,其中蛋壳是抵御外界物理损伤和微生物侵染的重要屏障;但在养殖过程中常出现大量的软壳蛋,严重影响禽蛋品质,且在运输和保存过程中易造成较大经济损失,直接影响养殖经济效益(张佳兰等,2008;Dunn et al.,2009)。因此,如何有效提高蛋壳品质已成为当前家禽育种的焦点,通常在育种方案中将分子标记育种与蛋壳品质选择相结合,以期进一步改善蛋壳品质。【前人研究进展】1,4,5-三磷酸肌醇(Inositol 1,4,5-trisphosphate,IP3)受体(IP3R)是四聚体细胞内钙离子(Ca2+)释放通道家族,几乎位于所有哺乳动物细胞类型的肌质网膜上,包括平滑肌细胞(Smooth muscle cells,SMC),其主要定位于内质网(Endoplasmic reticulum,ER),在细胞内的Ca2+释放通道中普遍表达,且通过开放响应IP3和Ca2+结合以调控Ca2+从细胞器中释放出来(Soulsby and Wojcikiewicz,2005;Narayanan et al.,2012)。在鸟类和哺乳动物中存在3种IP3R亚型(IP3R1、IP3R2和IP3R3),三者在其一级序列中具有高度的相似性,同源性达70%~80%(Chaloux et al.,2007)。尽管3种IP3R亚型的结构高度相似性,但其编码基因(IP3R1、IP3R2和IP3R3)的表达模式存在明显差异(Yamamoto-Hino et al.,1994;Wojcikiewicz,1995;Taylor et al.,1999),且在各种细胞类型中的表达程度也不同。IP3R1基因在神经元中高度表达;IP3R2基因主要在心肌和肝细胞中高度表达;IP3R3基因主要在快速增殖的细胞中表达,如上皮细胞(Kuchay et al.,2017)。IP3R3基因表达多种肿瘤坏死因子并调节肿瘤细胞的侵袭和增殖(Sakakura et al.,2003;Kang et al.,2017),可通过促进IP3R3和Ca2+介导的细胞凋亡,而抑制肿瘤生长(Kang et al.,2010);也可通过调节Ca2+以降低乳腺癌细胞的迁移能力(Mound et al.,2017)。此外,IP3R3基因表达与细胞的圆度指数呈负相关,其沉默可诱导细胞骨架重组,导致细胞周径变小,细胞黏附性降低;IP3R3基因沉默还显示出振荡的Ca2+浓度特征,在细胞骨架早期修复中发生主要的振荡特征(Furukawa et al.,2003;Morikawa et al.,2008;Tsai et al.,2015)。已有研究证实,鸡子宫上皮细胞中有IP3R3基因表达(Jonchère et al.,2010;Brionne et al.,2014),其子宫内膜钙化可增加IP3R3基因的表达水平,且明显高于同样具有活性钙代谢的脑膜和十二指肠,揭示IP3R3对子宫内膜细胞中的Ca2+转运具有重要调节作用,具体表现为促进蛋壳钙化(Rodríquez-Navarro et al.,2015)。生物矿化是一种钙化,是通过Ca2+与碳酸根离子(CO32?)的共沉淀形成蛋壳。蛋壳矿化的先决条件是通过跨细胞运输在有限的细胞外环境中供应大量Ca2+,因此需要离子通道、离子泵和离子交换剂的存在及活化(Jonchére et al.,2012;Nys and Roy,2018)。子宫是为蛋壳形成提供最快矿化的主要部位,Ca是蛋壳钙化过程中的必需元素,且不断从血液供应到子宫液中以抵抗浓度梯度(Nys et al.,2004)。Ca2+转移到子宫液中涉及IP3R3或其他Ca2+通道进入蛋壳腺细胞,其中,钙结合蛋白-D28k(CALB1)作用于细胞内转运,而Na+/Ca2+或Ca2+/H+交换确保Ca2+输出(Bahadoran et al.,2018)。【本研究切入点】目前,国内外针对IP3R3的研究主要集中在信号通路及其对Ca2+促细胞凋亡转移抗癌等方面,IP3R主要参与Ca2+释放,尤其是IP3R3对动物机体Ca2+的转运具有重要调控作用,但至今在家禽上鲜有研究报道。【拟解决的关键问题】以三穗鸭为研究对象,采用DNA直接测序法对IP3R3基因的SNP位点进行鉴定,并分析其与蛋壳品质的关联性,明确IP3R3对蛋壳性状的效应机制及其在蛋壳品质改良中的应用价值,为开展蛋壳品质改良分子标记育种提供参考依据,也为深入研究IP3R3基因的生物学功能打下基础。
1 材料与方法
1. 1 试验材料
随机选择饲养于贵州大学家禽研究所且同日出雏、健康无病、相同饲养管理条件下的三穗鸭288羽,于90日龄时翅静脉采血0.2~0.5 mL,采用血液/组织/细胞基因组提取试剂盒[DP304,天根生化科技(北京)有限公司]提取基因組DNA,以1.2%琼脂糖凝胶电泳和NanoDrop2000 DNA浓度测定仪(美国Thermo Scientific公司)联合评估DNA提取质量,稀释成100 ng/μL后保存备用。收集45周龄母鸭生产的鸭蛋,测定蛋重、蛋形指数、蛋壳厚度、蛋壳强度和蛋壳重共5个指标。
1. 2 引物设计及PCR扩增
根据GenBank收录的鸭基因组(NC_040072.1)序列信息,利用Primer 3.0(http://primer3.ut.ee/)设计5对特异性扩增引物,引物信息见表1。PCR反应体系20.0 μL:DNA模板1.0 μL,2×Es Taq MasterMix 10.0 μL,上、下游引物(10 pmol/μL)各1.0 μL,ddH2O 7.0 μL。扩增程序:95 ℃预变性8 min;95 ℃ 50 s,退火40 s,72 ℃ 50 s,进行35个循环;72 ℃延伸6 min。
1. 3 三穗鸭IP3R3基因SNP位点鉴定
对288羽三穗鸭的IP3R3基因扩增产物进行纯化,然后送至生工生物工程(上海)股份有限公司直接测序,分别采用DNAMAN和Chromas 2.4.1进行SNP位点筛查及鉴定。
1. 4 统计分析
使用Excel 2016计算三穗鸭IP3R3基因各SNP位点的基因型频率、等位基因频率、观测杂合度(Ho)、有效等位基因数(Ne)、期望杂合度(He)和多态信息含量(PIC),并进行Hardy-Weinberg平衡定律检验;采用SHEsis(http://analysis.bio-x.cn/)进行单倍型及连锁不平衡分析;利用ORF Finder进行氨基酸翻译,通过RNAfold(http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi)预测不同单倍型的mRNA二级结构和自由能;以SPSS 18.0中的一般线性模型(GLM)完成三穗鸭IP3R3基因SNP位点基因型与蛋壳表型数据的关联分析。
2 结果与分析
2. 1 SNP位点筛选及鉴定结果
利用设计的5对特异性引物对三穗鸭IP3R3基因进行PCR扩增,每个样品均单独直接测序和序列比对分析,结果如图1所示。在三穗鸭IP3R3基因外显子上发现2个SNPs位点,分别位于第49外显子的g.35195T>C和g.35207G>A,且均未引起编码氨基酸发生改变,属于同义突变。
2. 2 SNP位点群体遗传信息分析结果
通过对三穗鸭IP3R3基因的2个SNPs位点进行群体遗传信息分析,结果(表2)发现2个SNPs位点在三穗鸭群体中均存在3种基因型。g.35195T>C位点的优势基因型为TT,其频率为0.490;g.35207G>A位点的优势基因型为GG,其频率为0.625。g.35195T>C和g.35207G>A位点的优势等位基因分别为T和G,其频率分别是0.703和0.792,对应的PIC分别为0.330和0.276,均属于中度多态位点(0.25
2. 3 三穗鸭IP3R3基因SNP位点连锁不平衡及倍型分析结果
利用SHEsis分析三穗鸭IP3R3基因2个SNPs位点的连锁不平衡,结果显示,g.35195T>C位点与g.35207G>A位点间的D?和γ2分别为1.000和0.111,表明2个SNPs位点间不存在强的连锁不平衡(D>0.750,γ2<0.330)。2个SNPs位点联合共产生3种单倍型(H1、H2和H3),其中,优势单倍型H3(TG)的频率为0.495,劣势单倍型H2(TA)的频率为0.208(表3)。对3种单倍型进行组合共得到6种双倍型(H1H1、H1H2、H1H3、H2H2、H2H3和H3H3),其中,优势双倍型H1H3(CTGG)的频率为0.333,劣势双倍型H2H2(TTAA)的频率为0.031(表3)。
2. 4 三穗鸭IP3R3基因SNP位点对mRNA二级结构的影响
采用RNAfold对所构建的3种单倍型mRNA二级结构进行预测分析,结果(图2)显示,3种单倍型的mRNA二级结构存在一定差异(圆圈部分),对应的mRNA二级结构最小自由能也不同,其中,单倍型H1的mRNA二级结构最小自由能为-475.50 kJ/mol,单倍型H2的为-476.50 kJ/mol,单倍型H3的为-476.10 kJ/mol,说明在三穗鸭IP3R3基因外显子上发现的2个SNPs位点突变能引起基因mRNA二级结构和自由能改变。
2. 5 三穗鸭IP3R3基因SNP位点基因型与蛋壳品质的关联分析结果
利用SPSS 18.0分析三穗鸭IP3R3基因SNP位点各基因型与蛋壳品质的关联性,结果(表4)显示,g.35195T>C和g.35207G>A位点对三穗鸭蛋壳厚度、蛋壳强度、蛋形指数、蛋重和蛋壳重的影响均未达显著水平(P>0.05,下同),说明IP3R3基因单个SNP位点对三穗鸭蛋壳品质无显著影响。
2. 6 三穗鸭IP3R3基因SNP位点双倍型与蛋壳品质的关联分析结果
三穗鸭IP3R3基因SNP位点双倍型与三穗鸭蛋壳品质的关联分析结果(表5)表明,双倍型H1H1个体的蛋壳强度显著高于其他双倍型个体(P<0.05,下同),双倍型H2H2个体的蛋形指数显著低于其他双倍型个体,其他双倍型个体间均无显著差异。
3 讨论
IP3R是细胞内Ca2+释放通道家族,近年来人们对该受体的研究主要集中在调节信号通路上,而鲜见IP3R基因多态性对禽类生产性能影响的研究报道。本研究通过对PCR扩增产物直接测序,发现在三穗鸭IP3R3基因外显子上存在2个SNPs位点,其群体遗传信息分析结果显示,g.35195T>C和g.35207G>A位点均处于中度多态(0.25
本研究通过对三穗鸭IP3R3基因的2个SNPs位点进行分析,发现均为同义突变,不影响编码氨基酸改变。编码区的同义突变会影响该基因的表达功能,进而影响个体间的表现型(Ren et al.,2014)。Komar(2007)研究报道,天然存在的SNP位点可导致编码相同氨基酸的核苷酸序列在合成蛋白产物时,其功能和结构发生改变,可能是SNP位点对可变剪接调控进而干扰其生物学功能(Kelemen et al.,2013)。也有研究表明,MDR1基因中的同义突变导致底物特异性改变(Kimchi-Sarfaty et al.,2007),而山羊POU1F1基因的同义突变与其产奶量和出生体重有关(Lan et al.,2007)。说明同义突变也是相关研究中不能忽略的突变,因此非常有必要明确IP3R3基因同义突变与蛋壳品质间的关联机制。本研究通过对三穗鸭IP3R3基因2个SNPs位点进行连锁不平衡分析,结果表明,g.35195T>C位点与g.35207G>A位点间的D为1.000、γ2为0.111,两者间不存在强的连锁不平衡。2个SNPs位点联合共构建获得3种单倍型,其中,优势单倍型H3(TG)的频率为0.495,劣势单倍型H2(TA)的频率为0.208。在疾病遗传和性状分析研究中,基因对性状的影响表现为单倍型共同作用可能比单个SNP位点的作用更大(Akey et al.,2001),说明单倍型可在选育过程中发挥重要作用。采用RNAfold对所构建的3种单倍型mRNA二级结构进行预测分析,发现单倍型H1的mRNA二级结构最小自由能最大,为-475.50 kJ/mol,即单倍型H1的mRNA二级结构稳定性最低;单倍型H2的mRNA二级结构最小自由能最小(-476.50 kJ/mol),说明其mRNA二级结构最稳定。2个SNPs位点的改变均对mRNA二级结构产生一定影响,且有可能影响各细胞内的细胞因子,通过影响IP3R3通路对Ca2+的释放而影响蛋壳形成。
蛋壳基质蛋白基因的多态性被认为与蛋壳强度和蛋壳厚度有关(Dunn et al.,2009),也有研究证实鸡子宫上皮细胞中有IP3R3存在(Jonchère et al.,2010;Brionne et al.,2014)。为此,本研究利用SPSS 18.0分析三穗鸭IP3R3基因SNP位点各基因型与蛋壳品质的关联性,结果发现g.35195T>C和g.35207G>A位点对三穗鸭蛋壳厚度、蛋壳强度、蛋形指数、蛋重和蛋壳重的影响均未达显著水平。光照强度可影响蛋壳厚度,且较长光照时间可导致较早的性成熟(Lewis et al.,1999,2010)。母鸡随着年龄的增加其产蛋率下降,且软壳蛋和破裂蛋的发生率显著增加,卵壳钙化不良似乎是由于卵壳腺的某些功能障碍所致,还可能与IP3R3基因调节的Ca2+分泌有关(Joyner et al.,1987)。光照刺激主要通过视觉和激素调节系统来发挥作用,其相关基因则可能影响鸭的性成熟或激素分泌,进一步通过IP3R3调控Ca2+对蛋壳品质的影响,本研究选取45周龄(产蛋高峰期)的三穗鸭,说明蛋壳品质与年龄无关。此外,本研究发现2个SNPs位点共同联合引起的基因结构变化强于单个SNP位点对基因结构变化的影响,其对蛋壳品质的调控作用可能更有效;同时对2种单倍型进行组合共获得6种双倍型,经关联分析发现双倍型H1H1个体的蛋壳强度显著高于其他双倍型个体,而双倍型H2H2个体的蛋形指数显著低于其他双倍型个体,其他双倍型个体间均无显著差异。通过对双倍型的关联分析可降低单个SNP位点受环境、其他SNP位点及其他相关微效基因的影响,也充分说明采用双倍型关联分析在评估品种和种群遗传改良时更准确(李天科等,2015)。蛋壳品质性状受多种基因调控,除基因遗传外,还受温度、光照和日龄等相关因素的影响(张亚男,2017;刘聪等,2019;谭光辉等,2019)。因此,今后应从蛋白组和转录组等水平进行深入挖掘,提供更有育种价值的SNP位点或单倍型,以推动家禽育种发展。
4 结论
三穗鸭IP3R3基因与蛋壳品质有一定关联性。在三穗鸭IP3R3基因外显子上发现的2个SNPs位点能引起基因mRNA二级结构和自由能改变,其构建的双倍型可作为主效候选基因或与主基因紧密连锁的分子标记用于鸭蛋壳品质改良。
参考文献:
李天科,赵娟花,裴杰,梁春年,郭宪,秦文,阎萍. 2015. 牦牛Ihh基因组织表达分析、SNP检测及其基因型组合与生产性状的关联分析[J]. 畜牧兽医学报,46(1):50-59. [Li T K,Zhao J H,Pei J,Liang C N,Guo X,Qin W,Yan P. 2015. Tissue expression,SNP detection and association of genotype combination of Ihh gene with production traits in yak[J]. Acta Veterinaria et Zootechnica Sinica,46(1):50-59.]
刘聪,李华,张正芬,華国洪,罗杰灵,于辉. 2019. 清远麻鸡快慢羽品系产蛋性能比较分析[J]. 南方农业学报,50(7):1625-1631. [Liu C,Li H,Zhang Z F,Hua G H,Luo J L,Yu H. 2019. Comparative analysis of laying performance between early and late feathering strains in Qingyuan Pa-tridge Chickens[J]. Journal of Southern Agriculture,50(7):1625-1631.]
谭光辉,平立锋,熊建民,覃媛钰,吴磊,李杰章,张依裕. 2019. 三穗鸭IP3R3基因的表达、遗传变异及对蛋壳品质效应的影响[J]. 农业生物技术学报,27(11):2033-2041. [Tan G H,Ping L F,Xiong J M,Qin Y Y,Wu L,Li J Z,Zhang Y Y. 2019. Expression,genetic variation of IP3R3 gene and its effect on eggshell quality in Sansui duck (Anas platyrhyncha domestica)[J]. Journal of Agricultu-ral Biotechnology,27(11):2033-2041.]
張佳兰,赵玉琴,高玉鹏. 2008. 蛋鸡周龄对褐壳蛋蛋壳品质的影响[J]. 西北农业学报,17(2):48-50. [Zhang J L,Zhao Y Q,Gao Y P. 2008. Effect of age of brown hen on eggshell quality[J]. Acta Agriculturae Boreali-occidentalis Sinica,17(2):48-50.]
张亚男. 2017. 饲粮锰调控鸡蛋壳品质的作用机制研究[D]. 北京:中国农业科学院. [Zhang Y N. 2017. Dietary manganese supplementation modulated eggshell quality in laying hens[D]. Beijing:Chinese Academy of Agricultu-ral Sciences.]
Akey J,Jin L,Xiong M. 2001. Haplotypes vs single marker linkage disequilibrium tests:What do we gain?[J]. European Journal of Human Genetics,9(4):291-300.
Bahadoran S,Samani A D,Hassanpour H. 2018. Effect of heat stress on the gene expression of ion transporters/channels in the uterus of laying hens during eggshell formation[J]. Stress,21(1):51-58.
Brionne A,Nys Y,Hennequet-Antier C,Gautron J. 2014. Hen uterine gene expression profiling during eggshell formation reveals putative proteins involved in the supply of minerals or in the shell mineralization process[J]. BMC Genomics,15:220. doi:10.1186/1471-2164-15-220.
Chaloux B,Caron A Z,Guillemette G. 2007. Protein kinase A increases the binding affinity and the Ca2+ release activity of the inositol 1,4,5-trisphosphate receptor type 3 in RINm5F cells[J]. Biology of the Cell,99(7):379-388.
Dunn I C,Joseph N T,Bain M,Edmond A,Wilson P W,Milona P,Nys Y,Gautron J,Schmutz M,Preisinqer R,Waddinqton D. 2009. Polymorphisms in eggshell organic matrix genes are associated with eggshell quality measurementsin pedigree Rhode Island Red hens[J]. Animal Genetics,40(1):110-114.
Furukawa R,Maselli A,Thomson S A M,Lim R W L,Stokes J V,Fechheimer M. 2003. Calcium regulation of actin crosslinking is important for function of the actin cytoskeleton in Dictyostelium[J]. Journal of Cell Science,116(Pt 1):187-196.
Goliásová E,Wolf J. 2004. Impact of the ESR gene on litter size and production traits in Czech Large White pigs[J]. Animal Genetics,35(4):293-297.
Jonchère V,Brionne A,Gautron J,Nys Y. 2012. Identification of uterine ion transporters for mineralisation precursors of the avian eggshell[J]. BMC Physiology,12:10. doi:10.1186/1472-6793-12-10.
Jonchère V,Réhault-Godbert S,Hennequet-Antier C,Cabau C,Sibut V,Cogburn L A,Nys Y,Gaytron J. 2010. Gene expression profiling to identify eggshell proteins involved in physical defense of the chicken egg[J]. BMC Genomics,11:57. doi:10.1186/1471-2164-11-57.
Joyner C J,Peddie M J,Taylor T G. 1987. The effect of age on egg production in the domestic hen[J]. General and Comparative Endocrinology,65(3):331-336.
Kang S S,Han K S,Ku B M,Lee Y K,Hong J,Shin H Y,Antoine A G,Woo D H,Brat D J,Eun M H,Yoo S H,Chung K C,Park S H,Peak S H,Roh E J,Lee S J,Park J Y,Traynelis S F,Lee C J. 2010. Inhibition of the Ca2+ release channel,IP3R subtype 3 by caffeine slows glioblastoma invasion and migration and extends survival[J]. Cancer Research,70(3):1173-1183.
Kang S,Hong J,Lee J M,Moon H E,Jeon B,Choi J,Yoon N A,Paek S H,Roh E J,Lee C J,Kang S S. 2017. Trifluoperazine,a well-known antipsychotic,inhibits glioblastoma invasion by binding to calmodulin and disinhibiting calcium release channel IP3R[J]. Molecular Cancer The-rapeutics,16(1):217-227.
Kelemen O,Convertini P,Zhang Z Y,Wen Y,Shen M L,Falaleeva M,Stamm S. 2013. Function of alternative splicing[J]. Gene,514(1):1-30.
Kimchi-Sarfaty C,Oh J M,Kim I W,Sauna Z E,Calcaqno A M,Ambudkar S V,Gottesman M M. 2007. A “silent”polymorphism in the MDR1 gene changes substrate specificity[J]. Science,315(5811):525-528.
Komar A A. 2007. Silent SNPs:Impact on gene function and phenotype[J]. Pharmacogenomics,8(8):1075-1080.
Kuchay S,Giorgi C,Simoneschi D,Paqan J,Missiroli S,Saraf A,Florens L,Washburn M P,Collazo-Lorduy A,Castillo-Martin M,Sebti S M,Pinton P,Paqano M. 2017. PTEN counteracts FBXL2 to promote IP3R3- and Ca2+-mediated apoptosis limiting tumour growth[J]. Nature,546(7659):554-558.
Lan X Y,Pan C Y,Chen H,Zhang C L,Li J Y,Zhao M,Lei C Z,Zhang A L,Zhang L. 2007. An AluI PCR-RFLP detecting a silent allele at the goat POU1F1 locus and its association with production traits[J]. Small Ruminant Research,73(1-3):8-12.
Leal S M. 2010. Detection of genotyping errors and pseudo-SNPs via deviations from Hardy-Weinberg equilibrium[J]. Genetic Epidemiology,29(3):204-214.
Lewis P D,Danisman R,Gous R M. 2010. Photoperiods for broiler breeder females during the laying period[J]. Poultry Science,89(1):108-114.
Lewis P D,Morris T R,Perry G C. 1999. Light intensity and age at first egg in pullets[J]. Poultry Science,78(8):1227-1231.
Morikawa K,Goto T,Tanimura A,Kobayashi S,Maki K. 2008. Distribution of inositol 1,4,5-trisphosphate receptors in rat osteoclasts[J]. Acta Histochemica et Cytoche-mica,41(2):7-13.
Mound A,Vautrin-Glabik A,Foulon A,Botia B,Haque F,Parys J B,Ouadid-Ahidouch H,Rodat-Despoix L. 2017. Downregulation of type 3 inositol(1,4,5)-trisphosphate receptor decreases breast cancer cell migration through an oscillatory Ca2+ signal[J]. Oncotarget,8(42):72324-72341.
Narayanan D,Adebiyi A,Jaggar J H. 2012. Inositol trisphosphate receptors in smooth muscle cells.[J]. American Journal of Physiology. Heart and Circulatory Physiolgoy,302(11):H2190-H2210.
Nys Y,Gautron J,Garcia-Ruiz J M,Hincke M T. 2004. Avian eggshell mineralization:Biochemical and functional cha-racterization of matrix proteins[J]. Comptes Rendus Palevol,3(6-7):560-562.
Nys Y,Roy N L. 2018. Chapter 22—Calcium homeostasis and eggshell biomineralization in female chicken[J]. Vitamin D,1:361-382.
Ren G,Huang Y Z,Wei T B,Liu J X,Lan X Y,Lei C Z,Zhang C L,Zhang Z Y,Qi X L,Chen H. 2014. Linkage disequilibrium and haplotype distribution of the bovine LHX4 gene in relation to growth[J]. Gene,538(2):354-360.
Rodríquez-Navarro A B,Marie P,Nys Y,Hincke M T,Gautron J. 2015. Amorphous calcium carbonate controls avian eggshell mineralization:A new paradigm for understan-ding rapid eggshell calcification[J]. Journal of Structural Biology,190(3):291-303.
Sakakura C,Hagiwara A,Fukuda K,Shimomura K,Takagi T,Kin S,Nakase Y,Fujiyama J,Mikoshiba K,Okazaki Y,Yamagishi H. 2003. Possible involvement of inositol 1,4,5-trisphosphate receptor type 3(IP3R3) in the peritoneal dissemination of gastric cancers[J]. Anticancer Research,23(5A):3691-3697.
Soulsby M D,Wojcikiewicz R J H. 2005. The type III inositol 1,4,5-trisphosphate receptor is phosphorylated by cAMP-dependent protein kinase at three sites[J]. The Biochemical Journal,392(Pt 3):493-497.
Taylor C W,Genazzani A A,Morris S A. 1999. Expression of inositol trisphosphate receptors[J]. Cell Calcium,26(6):237-251.
Tsai F C,Kuo G H,Chang S W,Tsai P J. 2015. Ca2+ signa-ling in cytoskeletal reorganization,cell migration,and cancer metastasis[J]. BioMed Research International,2015:409245. doi:10.1155/2015/409245.
Wojcikiewicz R J. 1995. Type I, II, and III inositol 1,4,5-trisphosphate receptors are unequally susceptible to down-regulation and are expressed in markedly different proportions in different cell types[J]. The Journal of Biolo-gical Chemistry,270(19):11678-11683.
Yamamoto-Hino M,Sugiyama T,Hikichi K,Mattei M G,Hasegawa K,Sekine S,Sakurada K,Miyawaki A,Furuichi T,Haseqawa M. 1994. Cloning and characterization of human type 2 and type 3 inositol 1,4,5-trisphosphate receptors[J]. Receptors and Channels,2(1):9-22.
(責任编辑 兰宗宝)
