水温和雌雄配比对罗氏沼虾能量代谢的影响
2020-11-09张俊功戴习林丁福江
张俊功 戴习林 丁福江
摘要:【目的】探索水溫和雌雄配比对罗氏沼虾(Macrobrachium rosenbergii)能量代谢和生长的影响,为进一步分析与解决罗氏沼虾生长缓慢的问题提供科学依据。【方法】根据能量收支方程[摄食能(Ce)=生长能(Ge)+排粪能(Fe)+排泄能(Ue)+蜕壳能(Ee)+呼吸能(Re)]及凯式定氮法,分别计算罗氏沼虾各部分的代谢能值,分析4种水温(20、22、26和30 ℃)及3种雌雄配比(1∶1、1∶2和1∶3)对罗氏沼虾能量代谢和生长的影响。【结果】雌、雄罗氏沼虾的绝对摄食能均随水温上升而升高,其中雌虾在各水温处理组间差异显著(P<0.05,下同),雄虾则表现为非相邻水温处理组间差异显著;雄虾在26 ℃下的绝对生长能显著高于其他3个水温处理组,雌虾在26 ℃下的绝对生长能显著高于20和22 ℃水温处理组;雌、雄罗氏沼虾在20 ℃下均未蜕壳,在22~30 ℃下其绝对蜕壳能随水温上升而升高,在30 ℃时达最大值(雌虾1371±272 J/g,雄虾1035±598 J/g);雌、雄罗氏沼虾的特定生长率在26 ℃下显著高于其他3个水温处理组;罗氏沼虾的绝对呼吸能表现为高温处理组(26和30 ℃)高于低温处理组(20和22 ℃)。单性饲养下,性成熟雄虾的绝对摄食能、绝对生长能和特定生长率随数量上升而降低,且每个水槽放养1尾雄虾显著高于每个水槽放养2~3尾雄虾;性成熟雄虾绝对呼吸能则随数量上升而显著升高。不同雌雄配比下,性成熟罗氏沼虾的绝对生长能随雄虾数量上升而显著降低,雌雄配比1∶1的特定生长率显著高于雌雄配比1∶2和1∶3;雌雄配比1∶1的绝对排泄能显著低于雌雄配比1∶2和1∶3,其占摄食能的百分比随雄虾数量上升而上升。【结论】高温更适合罗氏沼虾的生长发育,且以26 ℃最适宜,在此水温条件下罗氏沼虾新陈代谢更旺盛。在性成熟罗氏沼虾生长过程中,其能量积累可能受打斗、占区和交配等生物学行为影响,且相同密度下雄虾比例增加有可能加剧影响。
关键词: 罗氏沼虾;水温;雌雄配比;能量代谢;生长
中图分类号: S966.121 文献标志码: A 文章编号:2095-1191(2020)07-1721-08
Abstract:【Objective】This article aimed to explore the effects of temperature and female-male mating ratio on energy metabolism and growth of Macrobrachium rosenbergii in order to provide scientific basis for further analysis and solution of the slow growth of M. rosenbergii. 【Method】According to the energy budget equation[the consumption energy(Ce)= growth energy(Ge)+defecation energy(Fe)+excretion energy(Ue)+molting energy(Ee)+respiratory energy(Re)]and the Kaiser method, the energy values of each part was calculated. the effects of four temperatures(20,22,26, 30 ℃) and three male-female ratios(1∶1, 1∶2, 1∶3) on energy metabolism and growth of M. rosenbergii were analyzed. 【Result】The absolute level of Ce of female and male M. rosenbergii increased with the increase of temperature, and was significantly different between groups of female M. rosenbergii, as well as groups of non-adjacent temperature levels of male M. rosenbergii(P<0.05, the same below). The absolute level of Ge of the male was significantly higher than that of other groups at 26 ℃, and that of the female was significantly higher than that of 20 and 22 ℃. The female and male did not molt at 20 ℃, and the absolute levels of Ee increased with the increase of temperature from 22 to 30 ℃, reaching the maximum at 30 ℃(female 1371±272 J/g,and male 1035±598 J/g). The specific growth rates(SGR) of female and male M. rosenbergii were significantly higher at 26 ℃ than that of the other three groups; the absolute level of Re of female M. rosenbergii was significantly higher in higher temperature groups(26 and 30 ℃) than that in lower temperature groups (20 and 22 ℃). In the test of sexually mature female and male shrimp raised separately, the absolute levels of Ce, Ge and SGRs were decreasing by the increasing population of male shrimp, that of one male shrimp in the sink significantly higher than that of two and three male shrimps in the sink; and the absolute level of Re of mature male shrimps increased significantly with the increase of population. In the test of different male-female mating ratios of M. rosenbergii, and there was a negative growth at the female-male ratio of 1∶3; the SGR of the female-male ratio at 1∶1 level was significantly higher than that at 1∶2 and 1∶3 levels, and the absolute level of Ue of the female-male ratios at 1∶1 level was significantly lower than that at 1∶2 and 1∶3 levels, and its share of Ce also increased by the increasing population of male shrimp. 【Conclusions】Relative high temperature is more suitable for the growth and development of M. rosenbergii, and 26 ℃ is the most suitable temperature, in which the metabolism of M. rosenbergii is more vigorous. It is concluded that the energy accumulation during the growth of sexually mature M. rosenbergii may be affected by its biological behavior such as fighting, occupying area and mating, and the increase of male shrimp proportion may aggravate the effect.
Key words: Macrobrachium rosenbergii; water temperature; female-male mating ratios,energy metabolism; growth
Foundation item:Shanghai Shrimp Industrial Technology System Construction Project(Hunongkechanzi〔2014〕No.5)
0 引言
【研究意义】罗氏沼虾(Macrobrachium rosenbergii)俗称马来西亚大虾、淡水长脚大虾,为个体最大的淡水虾(陈文静等,2000;董学洪和陈正兴,2015),是我国当前重要的养殖虾类品种之一。随着罗氏沼虾在印度及马来西亚等东南亚国家和地区的兴起,其全球产量与日俱增(Chettri et al.,2007;Karaket et al.,2011),但近年来罗氏沼虾养殖生产不稳定现象日趋严重,尤其是以摄食量减少、生长减缓和性早熟为主要特征的铁壳虾和老头虾,已成为制约其养殖业健康发展的瓶颈(董学洪和陈正兴,2015;袁锐等,2017;徐卫国和姚振海,2018)。此外,性成熟雄虾具有打斗和残杀的习性,以及雌虾性腺发育、抱卵和产卵等行为均会消耗体内积累的物质能量,而导致其生长发育缓慢。因此,开展对罗氏沼虾能量代谢研究对解决其生长缓慢的问题具有重要意义。【前人研究进展】种质退化是引起动物机体生长缓慢现象的主要原因之一(史建华等,2001)。袁锐等(2017)研究证实,遗传和环境因素同步影响罗氏沼虾的生长和发育,其中种质遗传因素是主要原因。单位面积养殖密度也影响罗氏沼虾的生长性能,一般以放养20000~60000尾/ha较合理(杨国梁,2008;贾若鲁,2018)。还有学者认为,由于亲虾感染某些病毒、致病菌或寄生虫(胡婷,2012;徐洋等,2012;朱凝瑜等,2015;程东远,2017;叶键等,2018)而导致养殖过程中出现性早熟现象,性早熟则降低生长过程中的能量积累水平。此外,苗期和成长期营养缺失及抗生素过量使用不仅降低沼虾后期存活率,还影响其生长发育(安振华等,2014;朱光来等,2019)。在罗氏沼虾的养殖过程中,除了受养殖水体盐度和离子含量水平等因素影响外(王顺昌等,2003;李树国等,2007),水温也是影响罗氏沼虾存活率和能量代谢水平的因素之一。当水温低于20 ℃时,罗氏沼虾仔虾存活率很低,而水温超过30 ℃时,罗氏沼虾亲虾易死亡(陈文静等,2000);水温高于22 ℃时罗氏沼虾性腺开始发育(杨国梁,2008),而25~30 ℃是罗氏沼虾的适宜养殖水温(张进等,2017);雌、雄性成熟罗氏沼虾交配和抱卵的适宜水温在26~28 ℃(杨国梁,2008)。说明20和30 ℃是罗氏沼虾适宜的养殖水温节点,而22和26 ℃分别是其性腺发育和交配产卵的水温起点,为罗氏沼虾养殖及亲虾培育提供了重要的参考依据。【本研究切入点】有关罗氏沼虾能量利用与代谢方面的研究,除了黄光华等(2017)探究罗氏沼虾幼体能量收支情况及朱其建等(2019)研究亚硝酸盐对罗氏沼虾能量代谢的影响外,至今鲜见其他针对罗氏沼虾成虾能量代谢的研究报道。【拟解决的关键问题】通过比较分析不同水温和雌雄配比下罗氏沼虾的能量代谢情况及其差异,并探讨相关因子对罗氏沼虾生长的影响,为进一步分析与解决罗氏沼虾生长缓慢的问题提供科学依据。
1 材料与方法
1. 1 试验用虾
供试罗氏沼虾为上海申漕特种水产开发公司池塘养殖的罗氏沼虾群体,暂养10 d后,挑选个头均匀、附肢健全、体表无损伤的个体,试验条件下驯化5 d后用于正式试验。
1. 2 试验设计
1. 2. 1 不同水温下罗氏沼虾的能量收支比较 根据罗氏沼虾生长发育对水温的要求(陈文静等,2000;杨国梁,2008;张进等,2017),设20、22、26和30 ℃ 4个水温处理,挑选规格较小且未抱卵的雌虾64尾,体质量(湿重,下同)为13.57±1.47 g,挑选同等规格雄虾64尾,体质量为17.97±1.61 g。试验前停食24 h以排空肠胃。每个水温处理下雌、雄虾分开饲养,各设4个重复,每个水槽4尾虾,每组外加1个等体积水体的水槽作为空白对照(用于开展投喂饵料的溶失试验,以矫正残饵量),试验周期15 d。
1. 2. 2 不同雌雄配比下罗氏沼虾的能量收支比较
根据赵云龙等(1995)的研究结果,挑选性腺成熟雌虾21尾(头胸甲清晰可见棕黄色卵黄),体质量为17.81±2.34 g;同时挑选性腺成熟雄虾42尾(均一规格下精荚全部饱满),体质量为24.03±3.20 g。试验前停食24 h以排空肠胃。在26 ℃的水温条件下,设3个梯度组,雄虾数量依次为1、2和3尾,雌虾各组均为1尾,每个梯度组設7个重复,外加1个等体积水体的水槽作为空白对照(用于开展投喂饵料的溶失试验,以矫正残饵量),试验周期10 d;然后组内雌、雄虾依次配对,相同条件下再饲养10 d。
1. 3 试验管理
试验用水槽均为78 cm×52 cm×48 cm的蓝色塑料箱,分别配备散气石持续充氧和500 W电加热棒控制水温,养殖水体溶解氧保持在5.5 mg/L以上。试验期间,每天8:00投喂饵料,饵料湿重约占试验虾体质量的10%。试验期间不换水,每个水槽水体因蒸发作用减少时补给蒸馏水至初始水位(40 cm)。
1. 4 样品收集及预处理
每个水槽作为1个样品处理,每天15:00时采用虹吸法吸出粪便,20:00时吸出残饵,试验结束后将所收集样品分类汇总。试验前后分别测定各水槽的水体总氮(水体充分搅拌);收集的蜕壳、粪便、残饵及虾体干重均在70 ℃下烘干至恒重,用0.1 mg的分析天平称量,记录后密封保存。
1. 5 罗氏沼虾特定生长率(SGR)及能值测定
采用凯氏定氮法测定各处理组蜕壳、粪便、残饵及虾体干重的含氮量(李杉杉,2018),并以HD-3000智能汉字量热仪测定其含热量(张硕和董双林,2002)。
罗氏沼虾特定生长率(%/d)=(lnWT - lnW0)/T×100
式中,W0是罗氏沼虾初始体质量(g),WT是罗氏沼虾终末体质量(g),T是试验天数(d)。
罗氏沼虾试验过程中能量收支方程计算式(张硕和董双林,2002):
摄食能(Ce)=生长能(Ge)+排粪能(Fe)+排泄能
(Ue)+蜕皮能(Ee)+呼吸能(Re)
摄食能(Ce)=摄食量×单位饵料能值
生长能(Ge)=终末虾总能量-初始虾总能量
终末虾总能量=终末虾体质量×终末虾体单位
能值
初始虾总能量=初始虾体质量×初始虾体单位
能值
排粪能(Fe)=粪便干重质量×单位排粪能值
蜕皮能(Ee)=蜕皮干重质量×单位蜕皮能值
排泄能(Ue)=(CN–GN–FN–EN)×2.483×104
式中,初始蝦体单位能值以取样同一群体中15尾同规格个体单位能值均值代替(J/g);各种能量的单位均为J;单位饵料能值、终末虾体单位能值、单位排粪能值和单位蜕皮能值均通过量热仪测定(J/g);CN为摄食饵料所含氮,GN为罗氏沼虾体内积累氮,FN为粪便所含氮,EN为蜕壳所含氮,其单位均为g;水体中少量氮统计在蜕壳氮中,2.483×104为每克氮的能值(J/g)。
1. 6 统计分析
能量计算结果以罗氏沼虾单位体质量为基准(J/g),采用SPSS 25.0对试验数据分别进行T 检验、单因素方差分析(One-way ANOVA)和Duncans多重比较。
2 结果与分析
2. 1 不同水温下罗氏沼虾的能量收支及生长差异情况
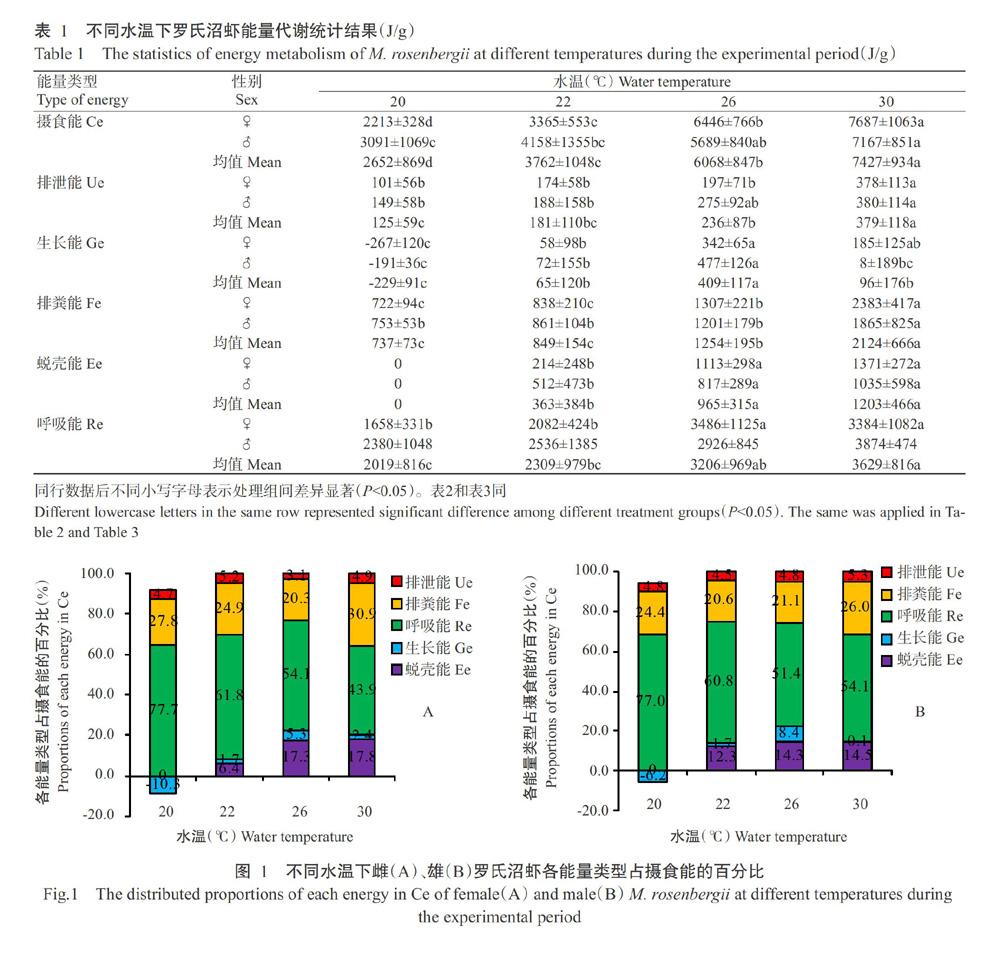
由表1可看出,雌、雄罗氏沼虾的绝对摄食能均随水温上升而升高,其中雌虾在各水温处理组间差异显著(P<0.05,下同),雄虾则表现为非相邻水温处理组间差异显著;低水温组(20和22 ℃)下雄虾的绝对摄食能高于雌虾,但高水温组(26和30 ℃)下雌虾的绝对摄食能高于雄虾。罗氏沼虾绝对排粪能和绝对摄食能的变化趋势基本一致,其中,雌虾相对排粪能(绝对排粪能占总摄食能的百分比)在20~26 ℃下呈下降趋势,而在26~30 ℃下呈上升趋势(图1-A),在26 ℃时达最低值(20.3%);雄虾相对排粪能在20~22 ℃下呈下降趋势,在22~30 ℃下呈上升趋势(图1-B),在22 ℃时达最低值(20.6%)。说明水温影响了罗氏沼虾的总摄食能和相对排粪能水平。
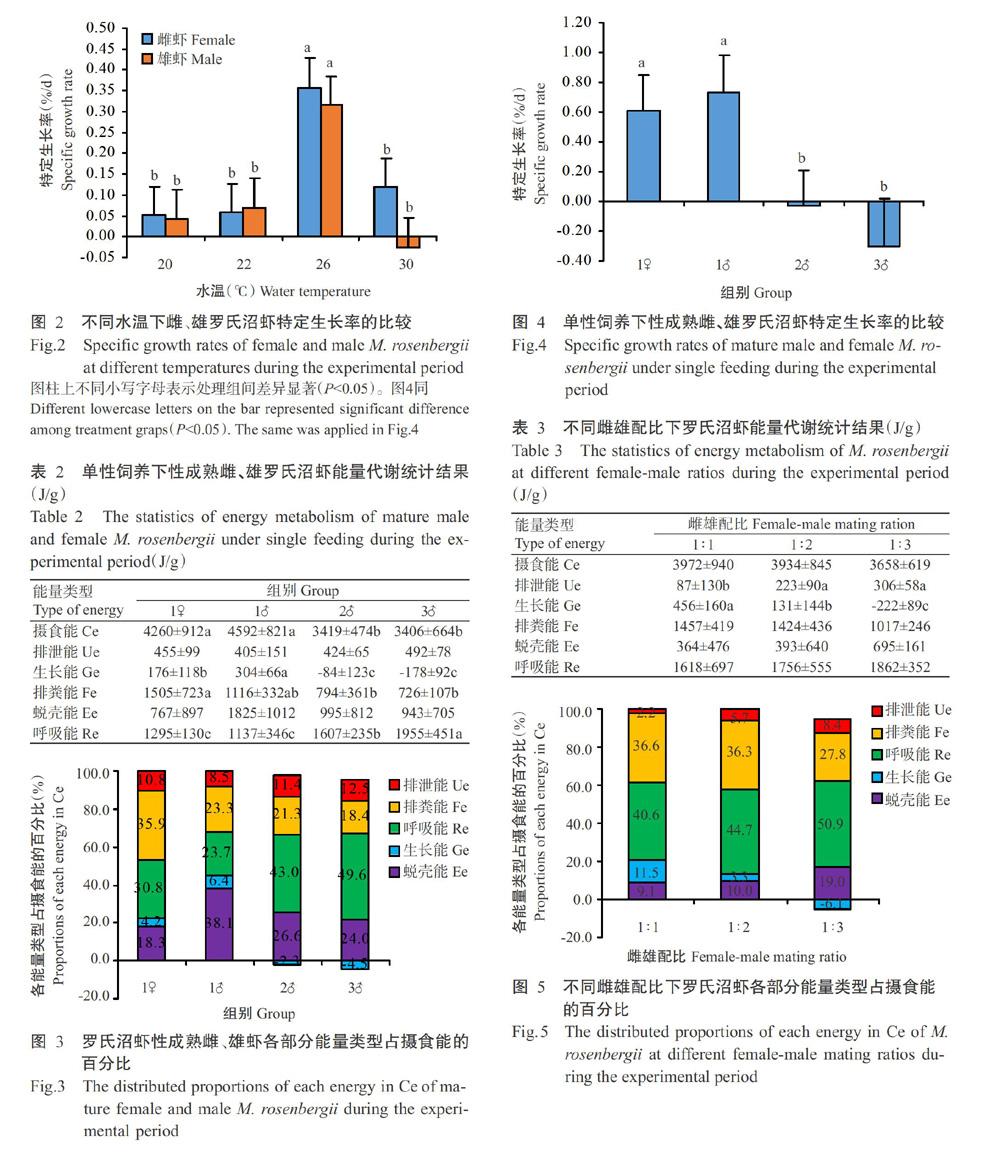
雌、雄罗氏沼虾绝对生长能在20~26 ℃下呈上升趋势,在26~30 ℃下呈下降趋势(表1),其中,雄虾在26 ℃下的绝对生长能显著高于其他3个水温处理组,雌虾在26 ℃下的绝对生长能显著高于20和22 ℃水温处理组;在20、22和26 ℃下雄虾的绝对生长能高于雌虾,但在30 ℃下雌虾高于雄虾。雌、雄罗氏沼虾在20 ℃下均未蜕壳,在22~30 ℃下其绝对蜕壳能均随水温上升而升高,在30 ℃时达最大值(雌虾1371±272 J/g,雄虾1035±598 J/g)。雌、雄罗氏沼虾的特定生长率在26 ℃下显著高于其他3个水温处理组(图2)。
罗氏沼虾绝对呼吸能在摄食能的分配中占最大比例(图1),且高温处理组(26和30 ℃)高于低温处理组(20和22 ℃),雌、雄虾的绝对呼吸能分别在26和30 ℃时达最大值(雌虾3486±1125 J/g,雄虾3874±474 J/g);而雌、雄虾的相对呼吸能(绝对呼吸能占绝对摄食能的百分比)均在20 ℃下达最大值,分别为77.7%和77.0%。雌、雄虾的绝对排泄能均随水温上升而升高,且雄虾普遍高于雌虾。说明水温影响着雌、雄罗氏沼虾的呼吸代谢水平。
2. 2 不同雌雄配比下罗氏沼虾的能量收支及生长差异情况
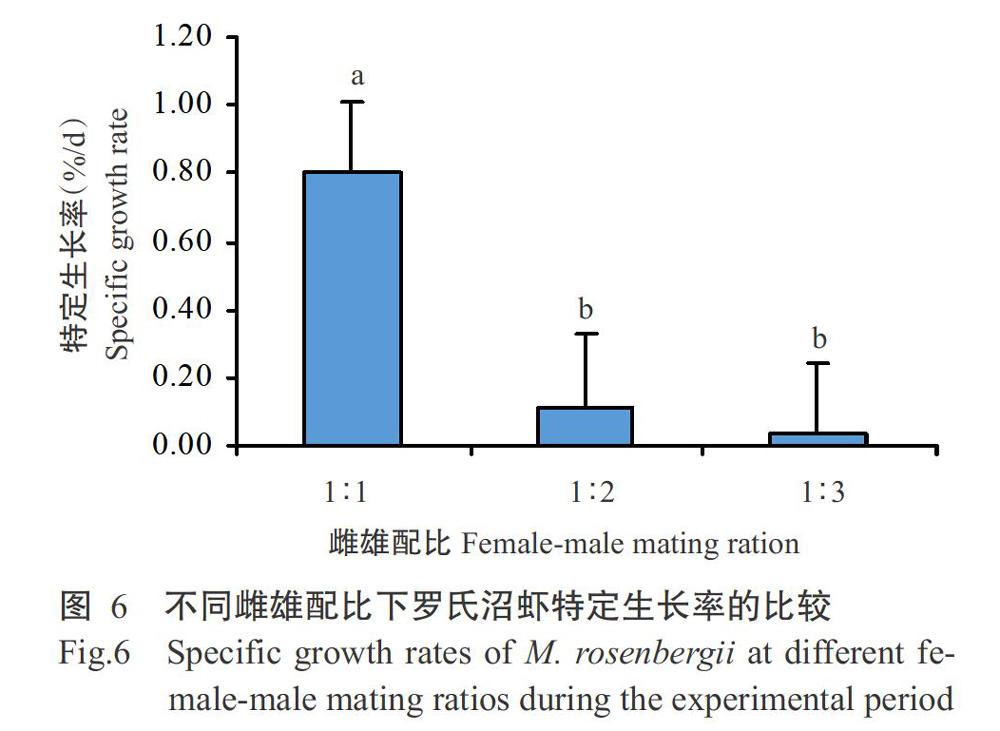
2. 2. 1 单性饲养下罗氏沼虾的能量收支及生长差异情况 由表2和图3可看出,性成熟雄罗氏沼虾的绝对摄食能和绝对排粪能水平均随数量上升而降低,其相对排粪能也随数量上升依次降低;1尾性成熟雌虾和1尾性成熟雄虾间的绝对摄食能差异不显著(P>0.05,下同)。性成熟雄虾的绝对生长能和特定生长率均随数量上升而降低,1尾性成熟雄虾的绝对生长能和特定生长率显著高于2尾和3尾性成熟雄虾(图4)。性成熟雄虾的绝对蜕壳能和相对蜕壳能均随数量上升而降低,绝对蜕壳能和绝对生长能的变化趋势基本一致。性成熟雄虾的绝对呼吸能随数量上升而显著升高,此时绝对排泄能也不断上升,但组间差异不显著。
2. 2. 2 不同雌雄配比下罗氏沼虾的能量收支及生长差异情况 由表3和图5可看出,性成熟罗氏沼虾的绝对摄食能随雄虾数量上升而降低,绝对排粪能和绝对摄食能的变化趋势相似,其绝对值和相对值均随雄虾数量上升而降低。性成熟罗氏沼虾的绝对生长能随雄虾数量上升而显著降低,雌雄配比1∶1的特定生长率显著高于雌雄配比1∶2和1∶3(图6);绝对蜕壳能和绝对呼吸能随雄虾数量上升而上升,但差异不显著;雌雄配比1∶1的绝对排泄能显著低于雌雄配比1∶2和1∶3,其占摄食能的百分比也随雄虾数量上升而上升。
3 讨论
水温是影响水生动物呼吸的重要因素之一(张明明等,2016)。本研究发现,水温对罗氏沼虾的能量代谢水平影响显著,同时影响罗氏沼虾的呼吸代谢及能量供给方式。在合适的水温范围内,虾类代谢速率随水温的上升而升高,各项生命活动也有所加强(王吉桥等,2004)。蜕壳是甲壳类动物实现生长的重要方式(徐建荣等,2006),且每次生长蜕壳都伴随着体质量的增长(宋宗岩等,2002)。本研究结果表明,在相同饲养管理条件下,雌、雄罗氏沼虾在26 ℃下的绝对生长能和特定生长率均显著高于20和22 ℃水温处理组,而30 ℃下的绝对蜕壳能高于前3个水温处理组。蜕壳也是罗氏沼虾生长的重要方式(张进等,2017),一定程度上能反映其体长和体质量的生长状况。通过统计绝对生长能和绝对蜕壳能之和发现,高水温处理组(26和30 ℃)显著高于低水温处理组(20和22 ℃)。可见,相对于低水温,高温更加适合罗氏沼虾的生长发育,且以26 ℃最适宜,在此水温条件下罗氏沼虾新陈代谢更旺盛。在本研究中,可能由于试验周期有限性,30 ℃下的罗氏沼虾绝对蜕壳能达最大值(1203 J/g),分别是22和26 ℃水温下的3.31和1.25倍,但未能及时实现体内物质能量积累和体质量增长。在合适的水温范围内,变温动物如鱼类的呼吸运动频率随水温升高而加快,呼吸速率也随之加快,通过呼吸作用消耗的能量也更多(沈旭明和赵清良,2001),与本研究中罗氏沼虾绝对呼吸能随温度上升而逐步升高的结论一致。
能量代谢水平是水产动物能量利用效率的重要体现。动物生理活动对能量代谢水平影响的研究已涉及鲢、鳙、鲤、暗纹东方鲀、斜带石斑鱼、鳊鱼等鱼类(童圣英,1997;沈旭明和赵清良,2001;黄建盛等,2017;曾令清等,2017)。单性饲养条件下,1尾雄虾的呼吸代谢耗能最小,能量利用效率和摄食率最高,生长率最大,与张硕和董双林(2002)在中国对虾最适盐度下的能量代谢结果基本一致。在罗氏沼虾外塘养殖模式下,常见池塘口有雄虾对峙或驱赶等行为现象,而这些常见的雄虾占区和打斗行为,其目的是抢占和守护属于自己的安全领域和食物(New,2005;张进等,2017),以保障自身安全及必要的生长发育条件。尤其在雌雄虾混养模式后期,性成熟个体增多,雄虾普遍出现争夺交配权行为(付广水,2018),且这种攻击行为是一种耗能活动(Briffa and Sneddon,2007;Copeland et al.,2011)。本研究结果表明,根据性成熟罗氏沼虾能量代谢,每个水槽1尾雄虾时的绝对摄食能显著高于每个水槽放养2~3尾;2尾雄虾由于打斗和抢食行为,导致单位虾体质量绝对摄食能降低1173 J/g,而绝对呼吸能增加470 J/g;3尾雄蝦时其呼吸能耗进一步增加。与雌雄配比1∶1相比,雌雄配比为1∶3时雄性个体间因抢夺交配权和打斗行为,致使其绝对呼吸能耗增加244 J/g。可见,3尾雄虾单养和雌雄配比1∶3时罗氏沼虾呼吸耗代谢耗能均有所增加,因而生长率降低。
伴随着罗氏沼虾的性成熟,一系列交配行为加剧了雄虾体内同化能的消耗(付广水,2018)。此外,雌虾个体通过摄食或消耗自身储存物质提供能量进行性腺发育和交配后的排卵和产卵,以及生殖细胞的发育和成熟(王春和成永旭,2009)。与1尾雌虾和1尾雄虾单养时的绝对呼吸能均值(1216 J/g)相比,罗氏沼虾雌雄配比1∶1时交配行为导致其绝对呼吸耗能增加402 J/g;与1尾雌虾和2尾雄虾单养时的绝对呼吸能加权均值(1503 J/g)相比,雌雄配比1∶2时交配行为导致其绝对呼吸耗能增加253 J/g;与1尾雌虾和3尾雄虾单养时的绝对呼吸能加权均值(1790 J/g)相比,雌雄配比1∶3时交配行为导致其绝对呼吸耗能增加72 J/g。可见,雌、雄罗氏沼虾的交配行为同样导致呼吸耗能有所增加。在本研究中,罗氏沼虾放养最大密度为11.4尾/m2,明显低于养殖生产密度(张进等,2017),足以消除或降低密度过高对罗氏沼虾能量代谢和生长的影响。
高能化合物三磷酸腺苷(ATP)是动物体内的直接能量来源,生物体合成ATP所需的能量来源于消化吸收饵料中的营养物质。动物的所有生命活动都离不开ATP,当细胞内ATP不足时,细胞内积累的营养物质发生分解,释放能量合成更多的ATP。雄性罗氏沼虾个体间的打斗、抢食、抢夺交配权和交配等行为,以及雌虾性腺发育、生殖蜕壳、怀卵和产卵等生殖活动,均需要细胞内线粒体产生大量ATP保障能量供给,呼吸代谢能耗上升,进而消耗罗氏沼虾体内积累的物质和能量。
4 结论
高温更适合罗氏沼虾的生长发育,且以26 ℃最适宜,在此水温条件下罗氏沼虾新陈代谢更旺盛。在性成熟罗氏沼虾生长过程中,其能量积累可能受打斗、占区和交配等生物学行为影响,且相同密度下雄虾比例增加有可能加剧影响。
参考文献:
安振华,孙龙生,陈佳毅. 2014. 罗氏沼虾“铁壳”现象出现原因探究[J]. 科学养鱼,(1):56-58. [An Z H,Sun L S,Chen J Y. 2014. Causes of the “iron shell” phenomenon of Macrobrachium rosenbergii[J]. Scientific Fish Far-ming,(1):56-58.]
陈文静,熊国根,邓勇辉,周智勇,裴建明. 2000. 罗氏沼虾生物学研究[J]. 江西水产科技,(4):15-19. [Chen W J,Xiong G G,Deng Y H,Zhou Z Y,Pei J M. 2000. Biology of Macrobrachium rosenbergii[J]. Jiangxi Fisheries Science and Technology,(4):15-19.]
程东远. 2017. 虾肝肠胞虫的流行病学及其致对虾生长缓慢机理的探索[D]. 上海:上海海洋大学. [Cheng D Y. 2017. The epidemiology of Enterocytozoon hepatopenaei and mechanism of retarded growth in shrimp[D]. Shanghai:Shanghai Ocean University.]
董学洪,陈正兴. 2015. 罗氏沼虾养殖中常见病害及防治[J]. 水产养殖,(11):46-48. [Dong X H,Chen Z X. 2015. Common diseases and prevention in cultivation of Macrobrachium rosenbergii[J]. Aquaculture,(11):46-48.]
付广水. 2018. 池塘养殖罗氏沼虾经验总结[J]. 渔业致富指南,(20):48-49. [Fu G S. 2018. Summary of experience in pond culture of Macrobrachium rosenbergii[J]. Fishery Guide to be Rich,(20):48-49.]
胡婷. 2012. 对虾传染性皮下及造血组织坏死病毒全基因序列的测定及快速诊断方法的建立[D]. 南京:南京农业大学. [Hu T. 2012. The determination of the whole gene sequence and establishment of the rapid diagnosis method of infectious hypodermal and hematopoietic necrosis virus[D]. Nanjing:Nanjing Agricultural University.]
黄光华,马华威,黎建斌,杨彦豪,黄立斌,冯鹏霏,吕敏,卢小花,杨学明. 2017. 人工海水对罗氏沼虾幼体生长发育及能量收支影響初探[J]. 水生态学杂志,38(2):76-81. [Huang G H,Ma H W,Li J B,Yang Y H,Huang L B,Feng P F,Lü M,Lu X H,Yang X M. 2017. Effects of artificial seawater on growth and energy budget of Macrobrachium rosenbergii larvae[J]. Journal of Aquatic Eco-logy,38(2):76-81.]
黄建盛,陈刚,张健东,汤保贵,王忠良,周晖. 2017. 响应曲面法分析低盐条件下温度与pH对斜带石斑鱼幼鱼能量代谢的联合效应[J]. 海洋渔业,39(1):41-50. [Huang J S,Chen G,Zhang J D,Tang B G,Wang Z L,Zhou H. 2017. Combined effects of temperature and pH on energy metabolism of Epinephelus coioides juveniles cultured in low salinity using response surface methodology[J]. Marine Fisheries,39(1):41-50.]
贾若鲁. 2018. 罗氏沼虾池塘养殖技术[J]. 现代农业科技,(15):221. [Jia R L. 2018. Pond culture technology of Macrobrachium rosenbergii[J]. Modern Agricultural Science and Technology,(15):221.]
李杉杉. 2018. 凯氏定氮法测定蛋白质的方法分析[J]. 现代食品,(21):140-142. [Li S S. 2018. Analysis of methods protein determination by Kjeldahl method[J]. Modern Food,(21):140-142.]
李树国,催海鹏,朱善央. 2007. 河蟹和罗氏沼虾幼体对海水中K+、Ca2+、Mg2+的需要量[J]. 水产科学,26(10):587-588. [Li S G,Cui H P,Zhu S Y. 2007. Requirements of chinese mitten card (Eriocheir sinensis) and prawn Macrobrachium rosenbergii Larvae of K+,Ca2+,Mg2+ in seawater[J]. Fisheries Science,26(10):587-588.]
沈旭明,赵清良. 2001. 温度、溶解氧对暗纹东方鲀幼鱼呼吸频率的影响[J]. 生态学杂志,20(4):13-15. [Shen X M,Zhao Q L. 2001. The effects of water temperature and DO on respiratory rates of immature Fugu obscurus[J]. Chinese Journal of Ecology,20(4):13-15.]
史建华,肖雨,徐琴英. 2001. 罗氏沼虾引种复壮技术的研究[J]. 水产科技情报,28(2):64-67. [Shi J H,Xiao Y,Xu Q Y. 2001. Studies on the introductional rejuvenated technique of Macrobrachium rosenbergii[J]. Fisheries Science & Technology Information,28(2):64-67.]
宋宗岩,王世党,王华东,徐承斌. 2002. 如何促进三疣梭子蟹正常蜕壳生长[J]. 齐鲁渔业,19(9):6-7. [Song Z Y,Wang S D,Wang H D,Xu C B. 2002. How to promote normal molting growth of Portunus trituberculatus[J]. Shandong Fisheries,19(9):6-7.]
童圣英. 1997. 四种鲤科鱼类越冬时脂肪酸组成的变化[J]. 水产学报,21(4):373-379. [Tong S Y. 1997. Changes in fatty acid composition in carp during winter[J]. Journal of Fisheries of China,21(4):373-379.]
王春,成永旭. 2009. 罗氏沼虾繁殖行为的再描述[J]. 动物学杂志,44(4):102-110. [Wang C,Cheng Y X. 2009. Re-description of reproductive behaviors of Macrobrachium rosenbergii[J]. Chinese Journal of Zoology,44(4):102-110.]
王吉桥,罗鸣,张德治,庞超. 2004. 水温和盐度对南美白对虾幼虾能量收支的影响[J]. 水产学报,28(2):161-166. [Wang J Q,Luo M,Zhang D Z,Pang C. 2004. Effects of water temperature and salinity on energy budget of Penaeus vannamei juveniles[J]. Journal of Fisheries of China,28(2):161-166.]
王顺昌,于敏,魏亦军. 2003. 水温对罗氏沼虾和青虾心率及组织中镁的影响[J]. 安徽大学学报(自然科学版),27(1):98-102. [Wang S C,Yu M,Wei Y J. 2003. Effects of ambient temperature on heart rate and tissue magnesium concentration between Macrobrachium rosenbergii and Macrobrachium nipponese[J]. Journal of Anhui University(Natural Science Edition),27(1):98-102.]
徐建荣,沈颂东,张加梅,蔡剑洪. 2006. 环境条件对中华绒螯蟹大眼幼体蜕壳生长的影响[J]. 水产科学,25(10):505-508. [Xu J R,Shen S D,Zhang J M,Cai J H. 2006. Influence of environment on molting and growth of Chinese mitten crab(Eriocheir sinensis) megalopa[J]. Fishe-ries Science,25(10):505-508.]
徐卫国,姚振海. 2018. 罗氏沼虾产业发展策略[J]. 渔业致富指南,(10):12-15. [Xu W G,Yao Z H. 2018. Development strategy of Macrobrachium rosenbergii industry[J]. Fishery Guide to be Rich,(10):12-15.]
徐洋,沈锦玉,姚嘉赟,潘晓艺,郝贵杰,尹文林. 2012. 罗氏沼虾主要病害研究概况[J]. 生物学杂志,29(6):74-76. [Xu Y,Shen J Y,Yao J Y,Pan X Y,Hao G J,Yin W L. 2012. Main diseases of the giant fresh water prawn Macrobrachium rosenbergii:A review[J]. Journal of Biology,29(6):74-76.]
楊国梁. 2008. 罗氏沼虾人工育苗及养殖新技术(上)[J]. 科学养鱼,(10):12-13. [Yang G L. 2008. New techniques of artificial nursery and culture of Macrobrachium rosenbergii (Part I)[J]. Scientific Fish Farming,(10):12-13.]
叶键,许婷,施礼科,王力,陈凡,郭水荣. 2018. 虾肝肠胞虫的研究进展[J]. 科学养鱼,(1):59-61. [Ye J,Xu T,Shi L K,Wang L,Chen F,Guo S R. 2018. Advances in studies on shrimp Enterocytozoon hepatopenaei[J]. Scientific Fish Farming,(1):59-61.]
袁锐,张朝晖,陈辉,方苹,陈静,刘训猛,吴亚锋,王晶晶. 2017. 罗氏沼虾“铁壳”现象及其防控研究进展[J]. 水产科学,36(3):383-390. [Yuan R,Zhang Z H,Chen H,Fang P,Chen J,Liu X M,Wu Y F,Wang J J. 2017. Phenomenon and research progress on prevention and control of “Iron Shell” in giant freshwater prawn Macrobrachium rosenbergii[J]. Fisheries Science,36(3):383-390.]
曾令清,付成,奚露,彭姜岚,付世建. 2017. 鳊幼鱼能量代谢和个性行为的个体变异及表型关联[J]. 生态学报,37(14):4807-4816. [Zeng L Q,Fu C,Xi L,Peng J L,Fu S J. 2017. Phenotypic correlations and individual variation of energy metabolism and personality in juvenile Chinese bream(Parabramis pekinensis)[J]. Acta Ecologica Sinica,37(14):4807-4816.]
张进,贺中华,王建新,邓家杰. 2017. 利用罗氏沼虾生物习性进行节水高效养殖试验[J]. 海洋与渔业,(1):62-64. [Zhang J,He Z H,Wang J X,Deng J J. 2017. Water-sa-ving and efficient aquaculture experiment of Macrobrachium rosenbergii by its biological habits[J]. Oceans and Fishery,(1):62-64.]
张明明,乔帼,仇明,徐梦尧,黄金田. 2016. 水生动物呼吸代谢研究进展[J]. 现代农业科技,(23):234-235. [Zhang M M,Qiao G,Qiu M,Xu M Y,Huang J T. 2016. Advances in respiratory metabolism of aquatic animals[J]. Modern Agricultural Science and Technology,(23):234-235.]
张硕,董双林. 2002. 饵料和盐度对中国对虾幼虾能量收支的影响[J]. 大连水产学院学报,17(3):227-233. [Zhang S,Dong S L. 2002. The effects of food and salinity on energy budget of juvenile shrimp of Penaeus chinensis[J]. Journal of Dalian Fisheries College,17(3):227-233.]
赵云龙,堵南山,赖伟. 1995. 罗氏沼虾繁殖习性的研究[J]. 水产科技情报,22(4):147-149. [Zhao Y L,Du N S,Lai W. 1995. Observation on the Reproductive behaviour of Macrobrachium rosenbergii(de Man)(Crustacea,Deca-poda)[J]. Fisheries Science & Technology Information,22(4):147-149.]
朱光来,齐富刚,王东博,陆丹,胡继悦,蔡朝霞. 2019. 益生菌对罗氏沼虾生长性能和水质的影响[J]. 中国饲料,(20):97-101. [Zhu G L,Qi F G,Wang D B,Lu D,Hu J Y,Cai Z X. 2019. Effect of probiotics on growth performance and water quality of Macrobrachium rosenbergii[J]. China Feed,(20):97-101.]
朱凝瑜,孔蕾,郑天伦. 2015. 2013年浙江省南美白对虾虾苗病毒携带情况分析[J]. 浙江农业学报,27(5):756-760. [Zhu N Y,Kong L,Zheng T L. 2015. Analysis on pre-sence of virus in Penaeus vannamei shrimp larvae in Zhejiang Province in 2013[J]. Acta Agriculturae Zhejiangensis,27(5):756-760.]
朱其建,鞠波,戴习林. 2019. 亚硝态氮对罗氏沼虾存活率、生长及能量代谢的影响[J]. 水产科技情报,46(1):14-18. [Zhu Q J,Ju B,Dai X L. 2019. Effect of nitrite nitrogen on survival rate,growth and energy metabolism of Macrobrachium rosenbergii[J]. Fisheries Science & Techno-logy Information,46(1):14-18.]
Briffa M,Sneddon L U. 2007. Physiological constraints on contest behaviour[J]. Functional Ecology,21(4):627-637.
Chettri J K,Sahu N P,Pal A K,Reddy A K,Kumar S,Kumar V. 2007. Comparative performance of Gamma Amino Butyric Acid(GABA) and 5-Hydroxytryptamine(5-HT) in the diet of larvae and post larvae of giant freshwater prawn,Macrobrachium rosenbergii:Effect of dose and route of administration on growth and survival[J]. Aquaculture,270(1-4):240-248.
Copeland D L,Levay B,Sivaraman B,Beebe-Fugloni C,Earley R L. 2011. Metabolic costs of fighting are driven by contest performance in male convict cichlid fish[J]. Animal Behaviour,82(2):271-280.
Karaket T,Poompuang S,Na-Nakorn U,Kamonrat W,Hallerman E M. 2011. DNA microsatellite-based evaluation of early growth performance among strains of freshwater prawn Macrobrachium rosenbergii de Man[J]. Aquaculture,311(1-4):115-122.
New M B. 2005. Freshwater prawn farming:Global status,recent research and a glance at the future[J]. Aquaculture Research,36(3):210-230.
(責任编辑 兰宗宝)
