氨基酸对食物过敏的调控机制研究进展
2020-11-06郑雪玥印遇龙邓近平
郑雪玥,王 浩,印遇龙,邓近平
(华南农业大学 动物科学学院 广东省动物营养调控重点实验室,广东 广州510642)
食物过敏被定义为“机体通过食入、皮肤接触或者吸入某种食物蛋白介导特异性免疫的不良反应”。据不完全统计,全球约5.2亿人受食物过敏影响,且患病率逐年上升[1]。食物过敏大多从幼龄阶段开始,多数伴随终生。过敏过程中常伴随着皮肤瘙痒、腹痛、腹泻等症状,严重时甚至导致休克和死亡[1]。食物过敏不仅是诱发犬、猫等经济动物皮肤及胃肠道损伤的因素之一[2],大豆中的β-伴大豆球蛋白等过敏原更诱发了仔猪食物过敏,可影响其断奶后数周的生长性能[3]。食物过敏威胁着患病机体的健康与安全,也对畜牧生产及社会医疗造成了巨大的经济负担。
氨基酸在机体免疫中发挥着不可替代的作用。天然免疫和适应性免疫系统的功能维持均对机体氨基酸稳态高度依赖,氨基酸比例失衡或氨基酸之间的拮抗作用会通过抑制机体营养物质吸收和代谢调控机体免疫系统。在过敏反应抗原递呈及免疫应答中发挥关键作用的树突状细胞、肥大细胞及嗜碱粒细胞等细胞的功能均受氨基酸的调控[4]。研究发现,氨基酸的合理使用可有效缓解牛奶[5]、花生[6]等过敏原诱发的食物过敏反应。但是,目前氨基酸在食物过敏中的应用研究较少,其对过敏反应的调控机制也仍不明晰,因此,现结合国内外研究进展,对氨基酸调控机体过敏反应的可能机制及应用前景进行综述,为食物过敏的缓解及治疗提供新的研究思路。
1 食物过敏的致敏机理
食物过敏不是由单一物质或生理紊乱形成的疾病,常常伴随多种疾病反应。过敏反应根据介导物质不同分为免疫球蛋白E(immunoglobulins E,IgE)介导、非IgE介导以及混合介导3种方式。绝大多数的食物过敏是由IgE介导的Ⅰ型过敏反应(又叫速发型过敏反应)[1]。现重点对IgE介导的Ⅰ型过敏反应的机制及氨基酸是否影响过敏进行探讨。
通常机体对食物抗原的默认应答是免疫耐受,主要由胃肠道的树突状细胞介导。这些抗原提呈细胞(antigen presenting cell,APC)分别穿过肠系膜淋巴结和局部淋巴结,诱导T细胞分化为调节性T细胞(regulatory T cells,Treg)。当发生食物过敏时,Treg细胞的诱导受到抑制,T细胞被诱导为抗原特异性的辅助性T细胞2(T helper 2 cells,Th2),驱动IgE介导的过敏反应[1]。
IgE介导的食物过敏反应主要分为2个阶段,第1阶段主要为机体致敏过程即机体产生特异性抗体IgE的过程。当外来食物抗原与APC及幼稚T细胞接触后,将使幼稚T细胞向Th2型细胞分化,引发Th2型反应,并分泌IL-4和IL-13,这2种细胞将促进B细胞产生的Ig发生类别转换,从而产生抗原特异性IgE。抗原特异性的IgE与肥大细胞及嗜碱粒细胞表面的高亲和力受体FcεRI结合,此过程称为致敏。第2个阶段为过敏反应。当过敏原再次出现时,抗原与肥大细胞或嗜碱粒细胞表面的特异性IgE结合,激活肥大细胞脱颗粒释放大量组胺和类胰蛋白酶等介质,以及肿瘤坏死因子-α(TNF-α)和IL-6等细胞因子;嗜碱粒细胞大量释放IL-4、IL-13等细胞因子,从而产生过敏反应组织特异性症状。有研究者在食物过敏患者的十二指肠中发现了一种新的黏膜肥大细胞,与正常人相比可产生更高水平的IL-9和IL-13,进一步通过抑制Treg细胞产生和诱导Th2细胞达到激活肥大细胞的目的[1]。近几年的研究发现,机体肠道菌群在食物过敏反应的发生中发挥关键作用,肠道菌群可通过调控Th2细胞免疫应答,黏膜免疫系统中IL-10等细胞因子的释放,骨髓嗜碱粒细胞的增殖及嗜碱粒细胞介导炎症反应,以及机体肠道屏障功能等途径调控食物过敏反应[7]。
2 氨基酸对过敏反应的调控及其可能作用机理
氨基酸是免疫细胞防御机制中重要的能量底物。机体基础氨基酸代谢是维系机体自身免疫稳态的必要因素,而氨基酸催化活性的提高可导致机体的免疫抑制[4]。超敏反应是由于机体在清除过敏原过程中免疫应答过于强烈,造成机体的组织损伤。研究表明,氨基酸参与多种过敏效应细胞的成熟或活化过程,如树突状细胞通过谷氨酸的释放影响T细胞的增殖及Th1细胞激活[8];肥大细胞中颗粒性介质多巴胺的存储过程依赖于酪氨酸和丝氨酸代谢[9]。外源氨基酸的使用也可调控机体过敏反应,如色氨酸可通过调控肠道菌群及抑制Th2细胞免疫应答,缓解过敏性哮喘[10],甘氨酸可通过抑制细胞因子释放等途径有效缓解小鼠乳清蛋白过敏[11]。
2.1 免疫细胞介导氨基酸调控过敏的可能机制
2.1.1氨基酸对树突状细胞的调控 树突状细胞是一种重要的特异性APC,由于T细胞受体(T cell receptor,TCR)不能直接识别抗原,经过APC识别并加工后与主要组织相容性复合体Ⅱ(major histocompatibility complex,MHCⅡ)分子结合形成抗原肽,因此,树突状细胞对抗原的递呈与加工是免疫应答的关键环节[12]。树突状细胞表面的FcεRΙ受体还可大幅提高树突状细胞抗原递呈效率。此外IL-33等细胞因子通过上调树突状细胞OX40L配体表达可诱导Th2细胞偏移,同样是引发过敏反应的关键步骤[1]。
如图1所示,氨基酸可参与树突状细胞成熟、发育及活化T细胞的过程。EIJI等[13]研究发现,机体血浆氨基酸水平的失衡会扰乱树突状细胞胞内三羧酸循环代谢,降低胞内腺苷三磷酸(ATP)水平,继而抑制树突状细胞的成熟。其多项研究还发现支链氨基酸(branched-chain amino acids,BCAA)参与调控树突状细胞的成熟及功能,尤其是缬氨酸的缺失将导致树突状细胞的成熟被抑制,而缬氨酸的添加可通过PI3K/mTOR/S6K信号通路,剂量依赖性的促进炎症因子IL-12的生成。IL-12是刺激幼稚T细胞向Th1细胞分化的关键细胞因子,可影响Th1/Th2细胞平衡[14-15]。谷氨酸及半胱氨酸也参与树突状细胞激活T细胞的过程,如树突状细胞可通过释放谷氨酸刺激T细胞的活化[16],间充质干细胞可通过阻抑树突状细胞释放半胱氨酸,从而抑制幼稚T细胞的扩增[17]。精氨酸及色氨酸可通过其代谢物调控树突状细胞功能,色氨酸代谢产物3-羟基邻氨基苯甲酸通过抑制树突状细胞成熟,并降低炎症因子IL-12、IL-6及TNF-α的生成,最终抑制T细胞的分化及活化[18]。精氨酸酶1(arginase 1,Arg1)和吲哚胺2,3-双加氧酶1(indoleamine 2,3-dioxygenase1,IDO1)分别催化精氨酸及色氨酸的降解,可造成细胞的局部氨基酸缺失,树突状细胞多胺的产生依赖于Arg1的表达。由树突状细胞生成的多胺,可激活具有IDO1磷酸化活性的Src激酶,从而调节树突状细胞向依赖IDO1的免疫抑制表型分化[19]。该研究暗示树突状细胞胞内精氨酸及色氨酸的代谢相互关联,且树突状细胞的生理功能严格受精氨酸及色氨酸缺失的调控。
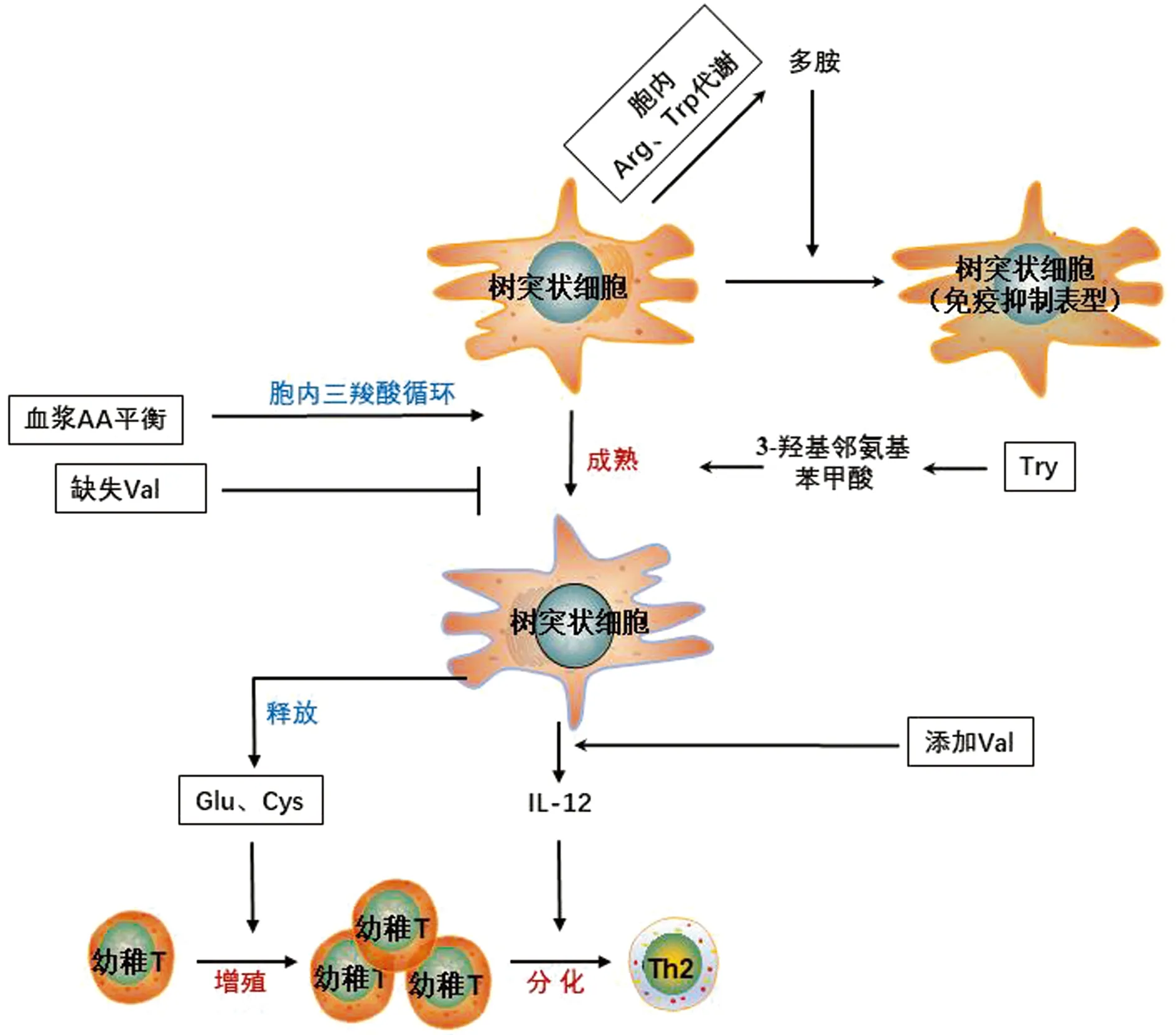
图1 氨基酸对树突细胞功能及代谢的调控
2.1.2氨基酸对Th2细胞的调控 正常机体内Th1和Th2细胞处于平衡状态,当Th1细胞活性降低或Th2细胞活性增强时会诱发机体的过敏反应。Th2细胞主要通过产生IL-2、IL-3、IL-4及IL-9等细胞因子调控肥大细胞、嗜碱粒细胞及B细胞等多种免疫细胞的免疫应答,从而调控过敏反应。其中IL-2作用于2型固有免疫细胞(group 2 innate lymphoid cells,ILC2)促进其分泌IL-4,抑制Treg细胞的生成及功能,加剧过敏反应;产生的IL-3主要作用为募集肥大细胞及嗜碱粒细胞进入淋巴结;IL-4则是驱动B细胞中Ig的类别转换,提高IgE水平的主要细胞因子;IL-9不仅是肥大细胞增殖的重要细胞因子,还可以增加肥大细胞对IL-3的敏感性[1]。
氨基酸对T细胞功能及代谢的调控作用如图2所示。研究发现,0.6 mmol/L谷氨酰胺可促进T淋巴细胞增殖,促进Th1细胞因子干扰素-γ(IFN-γ)和IL-2的生成及Th2细胞因子IL-4及IL-10的生成。但当谷氨酰胺水平提高时,Th1细胞免疫应答增强,Th2细胞免疫应答被抑制[20]。高海拔刺激的人体通过摄入谷氨酰胺也可调节机体的Th1/Th2细胞平衡向Th1偏移[21],此结果暗示可通过谷氨酰胺调节机体Th1/Th2细胞平衡,从而缓解机体过敏反应。1 mmol/L或10 mmol/L亮氨酸代谢中间产物β-羟基-β甲基丁酸,可导致Th1/Th2细胞平衡向Th2偏移[22]。

图2 氨基酸对T细胞功能及代谢的调控
2.1.3氨基酸对Treg细胞的调控 Treg主要通过诱导并维持机体对过敏原的免疫耐受,从而调控过敏反应。Treg可抑制树突状细胞的促炎特性以及树突状细胞对Th细胞的启动效应;此外,Treg还可通过直接抑制过敏原对Th2细胞的活化,从而抑制IL-4、IL-5等促炎细胞因子的分泌;Treg对肥大细胞及嗜碱粒细胞等效应细胞活化的抑制还会影响组织的重塑,继而抑制过敏性炎症。但在食物过敏的生理条件下,Treg细胞的生成被抑制,T细胞趋向于向Th2细胞分化,继而驱动Ig的类别转换及过敏效应细胞扩增[1]。
外源营养物质可调控CD4+幼稚T细胞向不同亚型分化。研究发现色氨酸、谷氨酰胺及支链氨基酸在维持T细胞向Treg亚型分化过程中均发挥关键作用。色氨酸缺失[23]或谷氨酰胺缺失[24]都将抑制幼稚CD4+T细胞向Th1细胞分化,并诱导其向Foxp3+Treg细胞分化。而支链氨基酸尤其是异亮氨酸可通过转运载体Slc3a2促进T细胞向Foxp3+Treg细胞分化,维持细胞增殖能力,并通过激活mTOR信号通路维持Treg细胞的正常生理功能[25]。精氨酸及瓜氨酸均可通过上调SIRT-1基因表达、下调SMAD7基因表达,促进Treg细胞的IL-10分泌。此外,瓜氨酸还可促进TGF-β的分泌,从而影响Treg细胞的免疫调节功能[26]。IL-10是Treg细胞调控机体免疫耐受的关键细胞因子,其可通过抑制CD28和ICOS依赖的T细胞共刺激,从而抑制T细胞活化;IL-10还可通过抑制单核细胞和树突状细胞的促炎因子、趋化因子及趋化因子受体表达增强机体免疫耐受;此外,IL-10可诱导Ig向非炎性的IgG4亚型转换,抑制IgE的生成,抑制肥大细胞及嗜碱粒细胞脱颗粒[27]。
2.1.4氨基酸对肥大细胞功能的调控 肥大细胞是关键的过敏效应细胞,肥大细胞在AGS细胞刺激下,分泌的IL-4不仅会抑制Treg细胞所维持的机体食物过敏原免疫耐受,还可驱动B细胞中Ig类别转换,提高机体抗原特异性IgE水平。抗原特异性IgE与肥大细胞表面的高亲和力受体FcεRΙ结合后,激活肥大细胞释放组胺、白三烯、5-羟色胺等多种炎性介质,作用于机体血管、平滑肌、结缔组织、腺体及炎症细胞,继而诱发过敏症状[1]。
胸腺基质淋巴细胞生成素(thymic stromal lymphopoietin,TSLP)被认为是通过树突状细胞及嗜碱粒细胞促进过敏化的关键因子之一。半胱氨酸可通过下调Caspase-1及NF-κB的基因表达,抑制肥大细胞生成TSLP[28]。肥大细胞来源于骨髓祖细胞,由趋化因子诱导募集进入外周组织,并在此过程中分化及成熟后发挥生理功能,精氨酸与谷氨酸的共同作用可抑制肥大细胞趋化因子单核细胞趋化蛋白(chemokinesmonocyte chemoattractant protein-1,CCL2)的表达,还可抑制活化后的肥大细胞胞内新合成介质白三烯C4(leukotriene C4,LTC4)的释放,并通过MAPK信号通路抑制促炎因子的基因表达[29]。在骨髓单核肥大细胞上的研究发现,组氨酸、赖氨酸和苏氨酸可抑制肥大细胞mTOR信号通路及IgE介导的肥大细胞激活[30]。氨基酸代谢产物也参与肥大细胞的活化过程,如色氨酸的代谢产物犬尿酸通过增强IgE介导的肥大细胞脱颗粒,促进介质LTC4的释放,并通过激活肥大细胞的PLCγ1、Akt及MAPK促进促炎因子IL-13的生成,从而促使肥大细胞被激活[31]。在疟疾相关研究的报道中发现,机体的精氨酸缺乏症与肠道肥大细胞增多及组胺水平升高有关,暗示精氨酸可能也参与肠道肥大细胞募集及细胞活化过程[32]。
2.1.5氨基酸对嗜碱粒细胞的调控 嗜碱粒细胞参与过敏反应的机制与肥大细胞类似,Th2细胞分泌的IL-3在淋巴组织中招募嗜碱性粒细胞,并增强嗜碱粒细胞中IL-4的产生,Th2细胞及嗜碱粒细胞通过IL-4的分泌促进Ig的类别转换,受到食物抗原刺激后,特异性的IgE通过血液和淋巴循环递呈给嗜碱粒细胞,与嗜碱粒细胞表面的FcεRI结合后,激活嗜碱粒细胞,导致组胺、白三烯、IL-13等介质的释放,继而引发过敏反应。而与肥大细胞不同的是,TSLP可直接招募嗜碱粒细胞进入淋巴结[1]。
氨基酸在嗜碱粒细胞中调控作用的研究较少,仅有研究探讨了甘氨酸对嗜碱粒细胞功能的影响,结果发现其对嗜碱粒细胞脱颗粒无影响,但可抑制细胞因子IL-4和IL-13的生成[11]。而IL-4和IL-13是诱导Th细胞分化,驱动B细胞中Ig类别转换,以及启动肥大细胞介导的黏膜屏障损坏的关键细胞因子[1]。有研究通过单细胞代谢组学解析了嗜碱粒细胞中色氨酸及组氨酸的代谢通路,研究发现胞内颗粒性介质组胺由组氨酸经L-组氨酸脱羧酶催化生成,血清素是由色氨酸转化而来,表明色氨酸及组氨酸参与嗜碱粒细胞胞内颗粒性介质的生成,但外源添加这2种氨基酸对嗜碱粒细胞介质生成及细胞功能的影响仍有待探讨[33]。
2.2 肠道菌群介导氨基酸对过敏的调控越来越多的研究证明,肠道菌群在过敏的发病机制及发病过程中发挥关键的调控作用[7]。对过敏性哮喘儿童的研究发现,其肠道大肠杆菌(Escherichiacoli)丰度及艰难梭状芽孢杆菌(Clostridiumdifficile)丰度与哮喘发生率显著正相关[34]。从猪小肠分离的大肠杆菌Escherichiacoli等细菌对谷氨酰胺的利用呈剂量依赖性增加[35]。此结果暗示谷氨酰胺可能通过对机体肠道Escherichiacoli代谢的调控缓解机体过敏反应。多项研究发现肠道分离的菌株可缓解机体过敏反应,多个梭状芽胞杆菌菌株,如Subdoligranulumvariabile、Anaerostipescaccae可缓解牛奶、鸡蛋等多种食物过敏[36]。梭状芽胞杆菌具有代谢色氨酸[37]及苯丙氨酸[38]的能力,色氨酸及苯丙氨酸参与树突状细胞、T细胞及肥大细胞等多个过敏效应细胞的功能及代谢调控。因此,梭状芽胞杆菌色氨酸及苯丙氨酸代谢可能是其调控机体过敏反应的机制之一。在日粮中添加蛋氨酸可提高家禽肠道Bacteroides,Bifidobacterium及Clostridium相对丰度[39],暗示可以通过外源添加氨基酸调控机体肠道菌群区系缓解机体过敏反应。
3 总结与展望
基于以上国内外氨基酸对过敏效应细胞及肠道菌群的调控研究进展,推测氨基酸调控过敏反应的主要途径可能有以下几点:(1)精氨酸、色氨酸参与树突状细胞分化;支链氨基酸、缬氨酸及色氨酸可调控树突状细胞成熟;谷氨酸、半胱氨酸参与树突状细胞激活T细胞的过程。(2)谷氨酰胺可调控T细胞增殖;谷氨酰胺及亮氨酸可调控Th1/Th2细胞平衡。(3)色氨酸及谷氨酸参与Treg细胞分化;支链氨基酸调控Treg细胞增殖,维持细胞功能;精氨酸、瓜氨酸可调控Treg细胞IL-10的分泌。(4)精氨酸可调控肥大细胞数量;精氨酸与谷氨酸可调控趋化因子水平;半胱氨酸、组氨酸、赖氨酸、苏氨酸及色氨酸可调控肥大细胞激活。(5)甘氨酸调控嗜碱粒细胞细胞因子分泌,组氨酸及色氨酸参与嗜碱粒细胞颗粒介质的合成。(6)谷氨酰胺、色氨酸、苯丙氨酸参与肠道菌群代谢,蛋氨酸调控有益菌群丰度。
综上所述,氨基酸的干预可能是缓解机体过敏症状的有效手段。大量的研究表明,氨基酸可维持过敏中免疫细胞的功能并调控与过敏相关肠道菌群的丰度。上述结果为开展及设计氨基酸缓解机体过敏相关方面的研究提供了理论依据,也揭示了开展相关研究需注意的一些客观问题:同一种氨基酸对不同的过敏效应细胞存在不同的调控作用及机制;不同浓度的氨基酸对同一过敏效应细胞可能存在相反的调控作用;多种氨基酸同时使用时可能存在协同或拮抗作用;机体在一些病理条件下,氨基酸平衡会出现紊乱。因此,在开展临床应用前,应开展大量的试验验证在不同的生理状态下,氨基酸对机体食物过敏调控效果的影响,从而为在不同生理或病理条件下,食物过敏机体的氨基酸应用量提供理论参考。
