恢复时间对力竭训练后的斑节对虾幼虾的抗氧化功能的影响*
2020-11-03陈梓聪陈丕茂袁华荣陈文静龙鑫玲张皓铭
陈梓聪 陈丕茂① 袁华荣 冯 雪 佟 飞 陈文静 龙鑫玲 张皓铭
(1. 中国水产科学研究院南海水产研究所 农业农村部南海渔业资源环境科学观测实验站中国水产科学研究院海洋牧场技术重点实验室 广东省渔业生态环境重点实验室 广东省海洋休闲渔业工程技术研究中心 广州 510300;2. 上海海洋大学海洋科学学院 上海 201306)
力竭运动作为体现游泳能力的标志和特殊的游泳状态,常见于野生水生动物在进行捕食、逃避敌害攻击、穿越激流等场景中。在养殖环境下,常见于水生动物在抢食、逃避捕捞、高密度暂养运输等环境中(Kieffer, 2000、2010)。对虾类运动能力较强,受到攻击或刺激时强烈收缩腹部肌肉,尾扇张开拨水,迅速后跳,逃离危险水域(Dall et al, 1990; 张沛东, 2006)。研究表明,对鱼虾进行适量运动训练可使其抗氧化能力提升,但过度运动会削弱机体的抗氧化能力,原因是在力竭运动后一段时间内,有氧呼吸强度显著升高,力竭运动积累的乳酸和有氧代谢产生的自由基等对组织器官形成胁迫,导致机体免疫力下降,体内抗氧化平衡被破坏(McKenzie et al, 2012; 虞顺年等,2018; 张海恩等, 2019)。大西洋鲑(Salmo salar)遭遇拥挤胁迫时,肌肉中的乳酸含量显著上升,糖原含量显著降低,皮质醇含量上升,出现机体呼吸代谢增强、细胞受损的迹象(Skjervold et al, 2001);黑鲷(Acanthopagrus schlegelii)在高密度运输过程中,呼吸代谢水平和SOD 活性均显著上升(李丹丹等, 2018a)。高密度养殖的斑节对虾(Penaeus monodon)与较低密度养殖组相比,出现耗氧率升高、生长性能低下的现象(Anand et al, 2019)。对虾类属于较为低等的无脊椎动物,进入力竭状态后相较于脊椎动物,需要消耗更多能量以调整紊乱的生理代谢状态,且需要更长时间恢复到生理平衡状态(Berry et al, 2019; 段妍, 2012;于晓明, 2009)。
斑节对虾是中国沿海重要养殖虾类和增殖放流品种(吴琴瑟, 2010; 邢诒炫等, 2014)。斑节对虾体内缺少特异性免疫应答反应,在陷入生理紊乱状态时,非特异性免疫能力下降,突然死亡率和疾病感染率升高,影响养殖或增殖放流的效果(Ratanapo et al, 1992;陈国福等, 2004)。因此,研究斑节对虾幼虾在力竭运动后的抗氧化能力恢复过程,对避免幼虾进入力竭状态和在幼虾进入力竭状态时的处理方法有重要指导意义,但目前尚未见相关的报道。本研究模拟斑节对虾在生活水体中避敌、抢食等力竭运动情况,探究其在力竭运动后,体内相关抗氧化能力指标随恢复时间延长的变动与恢复过程,以期为斑节对虾养殖生产及增殖放流活动期间幼虾的管护提供参考。
1 材料与方法
1.1 材料
实验使用的斑节对虾幼苗为中国水产科学研究院南海水产研究所深圳基地自行繁育的养殖虾苗,标粗后,挑选约1000尾规格相近、体长为(3.0±0.4) cm、体重为(0.20±0.07) g的幼虾进行力竭运动胁迫实验。恢复实验期间,前24 h禁食,之后每日分3次(08:00、16:00和23:00)投喂虾苗总体重的3%的零号料(约2.25 g)。力竭运动实验装置为自行设计的环形水道(外径为1004 mm,内置隔离装置直径为604 mm,高为395 mm,容量为200 L)。
1.2 实验方法
力竭实验前,从未进行力竭运动胁迫的养殖幼虾中随机挑选12 尾作为空白组,将其余幼虾分为7 组,每组3 个平行,分别放入环形水道中进行力竭运动胁迫。胁迫实验期间,水道中流速保持在0.25 m/s,用小抄网从幼虾游泳方向前方惊吓幼虾,使幼虾持续进行跳跃逃避运动,当幼虾对刺激无法进行逃避、游泳足摆动频率下降、无法再逆水流游泳的时候即进入力竭状态。此时,将全部存活幼虾放入室内的暂养池的网箱中恢复,同时,挑选12 尾力竭状态的幼虾,置于冰冷的取样盘中取样,并且在未进行捕捞刺激的幼虾中取样作为对照组样品,经液氮急冻后保存于–80℃环境中,此为力竭运动后0 h 的样品。之后在放入恢复水体后开始计时,在1、4、8、12、24、48和72 h 以同样方法取12 尾幼虾,并在取样前观察幼虾的行为。
在所有样品采集完成后,测定整尾幼虾的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、谷胱甘肽过氧化物酶(GSH-Px)的活性,总抗氧化能力(T-AOC)、丙二醛(MDA)含量,以上指标的测定均使用南京建成生物工程研究所的试剂盒。
1.3 数据处理
实验数据经Excel 2016 处理后,使用SPSS 21 软件进行分析,以力竭运动后的恢复时间为自变量,对各生理指标按取样时间进行单因素方差分析(One-way ANOVA),若组间出现极显著(P<0.01)或显著差异(P<0.05),则用Duncan 法进行组间差异显著性检验,数据以平均值±标准差(Mean±SD)表示。
2 结果
2.1 斑节对虾幼虾力竭运动后不同恢复时间的体内抗氧化相关指标变化
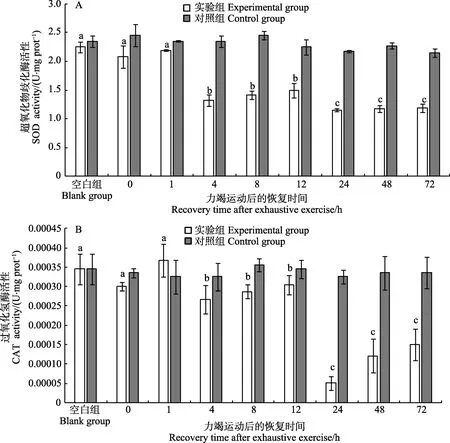
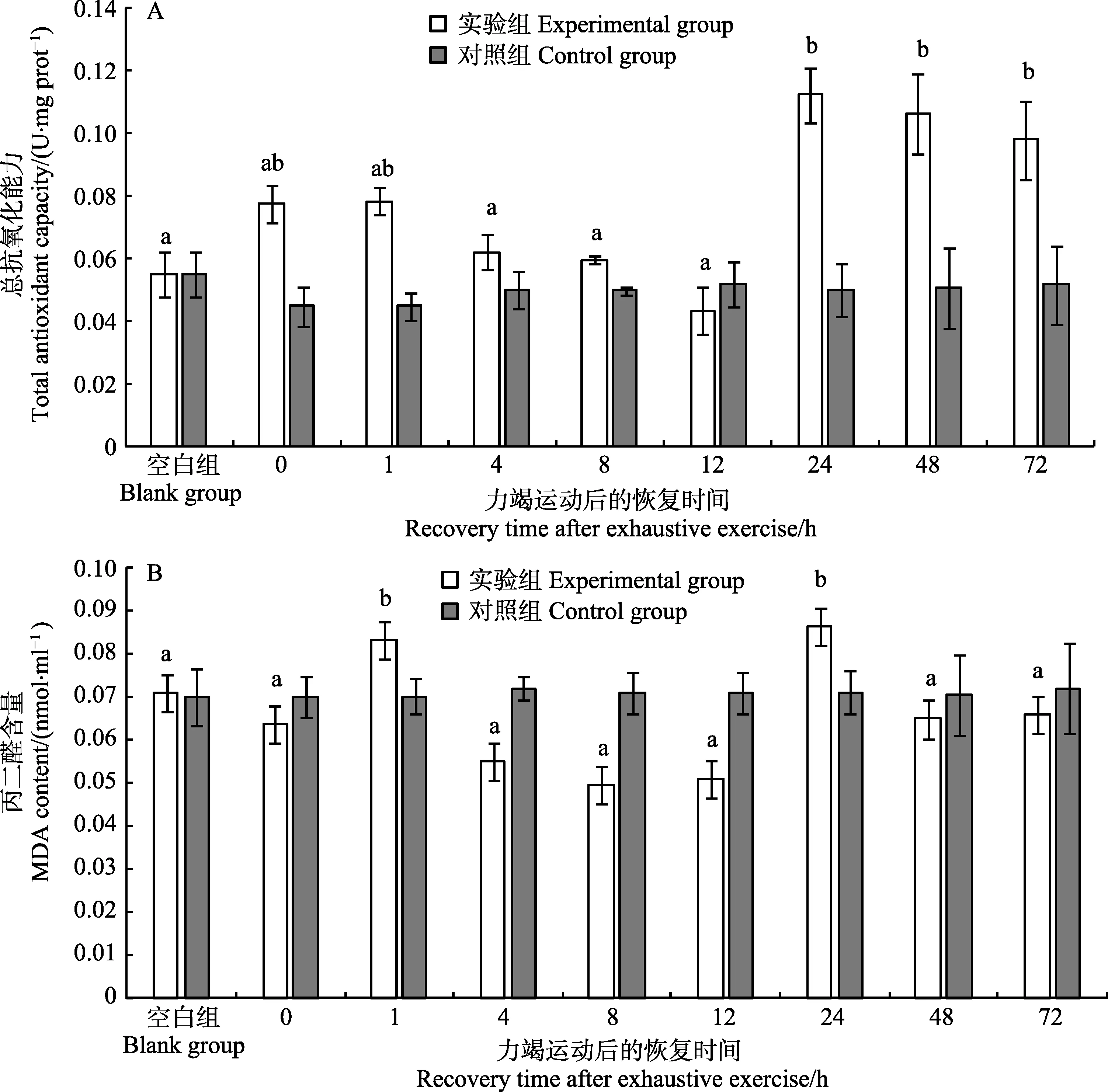
如表1 所示,斑节对虾幼虾在力竭运动后,随恢复时间的延长,总抗氧化能力、SOD、CAT 和POD 活性均有显著差异(P<0.05);MDA 含量存在差异,但差异不显著(0.05 表1 斑节对虾幼虾在力竭运动后各恢复时间点的抗氧化指标单因素分析结果Tab.1 Analysis of oxidation resistance in juvenile P. monodon after exhaustive exercise by one-way ANOVA 在力竭运动后恢复的0~4 h 期间,斑节对虾幼虾无附壁、游动等行为,基本保持在放入网箱时所处位置,对捕捞刺激无躲避行为;8 h 后,幼虾开始躲藏在网箱角落等位置,并有少量附壁行为,并能对捕捞作出反应;12 h 后,附壁的幼虾数量增多;24~72 h期间,附壁、游动和爬行的幼虾数量持续增加。 2.3.1 超氧化物歧化酶活性的变化 如图1A 所示,斑节对虾幼虾在力竭运动后的0~1 h 内,SOD活性并没有显著变化;4 h 时,下降至显著低于对照组的水平(P<0.05);此后,SOD 活性持续保持在较低水平,72 h 后,SOD 活性仍未恢复至对照组水平(P<0.05)。 图1 斑节对虾幼虾力竭运动后体内的SOD(A)和CAT(B)活性的变化Fig.1 SOD (A) and CAT (B) activities in juvenile P. monodon after exhaustive exercise 2.3.2 过氧化氢酶活性的变化 如图1B 所示,斑节对虾幼虾在力竭运动后的0~12 h 内,CAT 活性并无显著变化;但CAT 活性在12~24 h 内降低至显著低于对照组的水平(P<0.05),在72 h 后开始缓慢恢复。 2.3.3 过氧化物酶活性的变化 如图2A 所示,斑节对虾幼虾在力竭运动后的0~1 h 内,POD 活性与对照组水平并无显著差异;但POD 活性从1 h 后开始下降,并在8 h 下降至最低点(P<0.05);POD 活性在第8 h 后开始缓慢恢复,在48 h 后恢复至比对照组的略高的水平(P<0.05)。 2.3.4 谷胱甘肽过氧化物酶活性的变化 如图2B所示,斑节对虾幼虾在力竭运动后的 0~1 h 内,GSH-Px 活性上升至显著高于对照组的水平(P<0.05);之后在1~24 h 内波动上升,在24 h,活性上升至显著高于对照组的水平(P<0.05);在24 h 后,GSH-Px活性开始下降,并在72 h 恢复至对照组水平。 如图3A 所示,斑节对虾幼虾在力竭运动后,体内的总抗氧化能力上升,但与对照组相比并无显著差异;在1 h 后,总抗氧化能力持续降低,在12 h 至最低水平,显著低于1 h;而后,总抗氧化能力在24 h达到最高点并高于对照组(P<0.05),并在72 h 后仍保持较高水平。 如图3B 所示,斑节对虾幼虾在力竭运动后1 h内,体内的MDA 含量上升,显著高于对照组;而后在1~12 h 内持续显著下降,低于对照组水平(P<0.05),之后,12~24 h 内MDA 含量显著升高(P<0.05),然后又逐渐回落至对照组水平。 图2 斑节对虾幼虾力竭运动后体内的POD (A)和GSH-Px (B)的变化Fig.2 POD (A) and GSH-Px (B) activities in juvenile P. monodon after exhaustive exercise 图3 斑节对虾幼虾力竭运动后体内的T-AOC 能力(A)和MDA 含量(B)的变化Fig.3 Total antioxidant capacity (A) and MDA content (B) in juvenile P. monodon after exhaustive exercise 抗氧化酶分布于机体各个组织,可高效清除细胞内自由基,保护细胞和脂质免于自由基的伤害(陈建波等, 1999)。幼虾的能量物质储存量少,相较于成熟个体,免疫力和维稳力更低(陈勇, 2016; 张海恩等, 2019)。超氧化物歧化酶(SOD)通过将生物体内超氧根阴离子( O2-)转化成H2O2以清除超氧根阴离子(董亮等, 2013),谷胱甘肽过氧化物酶(GSH-Px)、过氧化氢酶(CAT)和过氧化物酶(POD)通常协同工作,将生物体内的H2O2分解成H2O 和O2。GSH-Px 除能清除H2O2外,亦有清除能使脂类过氧化的物质( ⋅OH)的功能(刘冰等, 2005;任海等, 2014;田清涞等,1992)。因此,抗氧化酶活性高低是反映动物机体的抵御氧化应激能力高低的生理指标。有氧代谢强度升高会对机体的抗氧化平衡造成冲击并影响机体健康,中华倒刺鲃(Spinibarbus sinensis)在力竭训练后肌肉组织中的SOD 活性显著上升(于丽娟, 2014);力竭运动会导致欧鲢(Leuciscus cephalus)的体内出现氧化应激的迹象,但SOD、GSH-Px 活性并无显著变化,说明抗氧化系统并没有因运动刺激而激活(Aniagu et al,2006);拥挤使南美白对虾(Penaeus vannamei)肌肉中的SOD、GSH-Px 活性显著降低(王彦波等, 2013)。 本研究中,如图1、图2 所示,力竭运动后0~1 h内,SOD、CAT 和POD 活性无显著变化,说明力竭运动过程中,幼虾机体酶促抗应激体系并未激活;但在4 h 后,SOD、CAT 和POD 活性持续下降,其中,POD 活性在8 h 后逐渐恢复,并在48 h 恢复至对照组水平,SOD、CAT 活性72 h 后仍显著低于对照组水平,说明机体在力竭运动后的4 h 后,幼虾机体开始出现氧化应激迹象,并消耗机体中常驻的SOD、CAT 和POD 进行调节;GSH-Px 活性在力竭运动后前24 h 内显著上升,与总抗氧化能力的变化趋势接近,说明GSH-Px 在幼虾的抗氧化体系中占据主要地位。通过观察对虾恢复期间的行为学变化后发现,幼虾在力竭运动后前4 h 内基本不进行运动,推测此时机体的主要工作为清除机体中积累 O2-和H2O2。产生氧化应激反应的原因可能是力竭运动后的爆发性有氧代谢(李丹丹等, 2018b),并且GSH-Px 活性比其他3 种抗氧化酶的活性高出几个数量级,说明在力竭运动恢复过程中的氧化应激反应中,GSH-Px 扮演比其他抗氧化酶更重要的角色;在48 h 时,各抗氧化酶活性恢复到对照组水平,说明幼虾在力竭运动后的48 h 后的抗氧化能力足以消除体内的氧化压力,但机体需要到72 h 才能使体内的抗氧化能力恢复至新的平衡状态。 总抗氧化能力反映了机体清除氧化性物质的能力和免疫与保护能力的高低(郑洁等, 2017)。组成机体总抗氧化能力的2 个体系为酶促反应体系和非酶促反应体系,酶促反应为抗氧化酶催化的氧化还原反应链;非酶促反应包括各类生物还原性物质(陈建波等, 1999; 韩星星等, 2018; 张涛等, 2018)。谢剑华(2008)在日本沼虾(Macrobrachium nipponense)日粮中额外添加虾青素,显著提升了其养殖成活率和抗应激能力。在运动训练后,黑鲷与中华倒刺鲃组织中的总抗氧化能力均有显著提升(于丽娟, 2014; 虞顺年等,2018)。 丙二醛作为生物体内脂类过氧化反应的最终产物,而脂类是生物体内构成膜系统主要的物质和储能物质,丙二醛含量被认为是反映生物机体受损伤程度的关键指标(宋志明等, 2015; 王彦波等, 2013)。日本沼虾在面对低氧胁迫时,肝胰腺与肌肉中的MDA 含量在0~12 h 内呈逐渐上升的趋势(管越强等, 2010)。养殖密度较高组的中国对虾(Fenneropenaeus chinensis)幼虾体内的MDA 含量比密度较低组更高(张海恩等,2019)。MDA 含量是反映机体受损伤程度、对胁迫敏感度和反应烈度的直接指标(杨玉姣等, 2006)。 如图3 所示,斑节对虾幼虾在力竭运动结束后的72 h 内,体内的MDA 含量的变化呈双峰形状,第1个峰值出现在1 h 处,第2 个峰值出现在24 h 处。幼虾力竭运动前后MDA 水平并无显著差异,说明力竭运动过程并没有对幼虾机体造成伤害,但总抗氧化能力已出现上升趋势,说明机体的非酶抗氧化体系已经激活。如上文所述,力竭运动前后各抗氧化酶活性与对照组水平无显著差异,说明在力竭运动后恢复的1 h 内,机体中的非酶还原物质提供了一定的抗氧化能力。1 h 时,幼虾的体内MDA 含量出现较明显的升高,总抗氧化能力与MDA 含量同步上升,说明在短时间内幼虾的抗氧化系统感受到体内出现氧化应激并作出响应,但MDA 含量的上升反映了机体的非酶体系无法在短时间内消除氧化物质对机体的损伤;在1~12 h期间,幼虾体内的MDA 含量和总抗氧化能力逐渐降至最低点,均显著低于最高点的1 h 的水平,说明幼虾通过消耗体内的抗氧化物质,合成和消耗相关抗氧化酶维持了体内的抗氧化平衡,并且幼虾在12 h 后恢复了常见的附壁和游泳等行为;12~24 h 期间,由于凌晨时幼虾开始恢复游泳摄食行为,呼吸代谢活动增强,抗氧化平衡再次波动,MDA 含量显著升高,抗氧化体系被持续激活,机体通过合成GSH-Px 等抗氧化酶提升机体总抗氧化能力;24~72 h,幼虾体内的MDA 含量恢复到对照组水平,总抗氧化能力仍高于对照组水平,说明幼虾机体的抗氧化能力恢复过程存在过量补充的“矫枉过正”现象,使机体存在消除此前的氧化应激并应对潜在氧化应激的可能。 斑节对虾幼虾在力竭运动过程中,体内不会产生氧化应激反应,但在力竭运动后恢复过程中,由于有氧代谢补偿性增强,导致机体氧化压力升高,首先激活了幼虾的非酶氧化应激反应,在非酶抗氧化体系无法消除氧化压力后,激活酶促抗氧化应激体系。而后,较长时间内幼虾体内的抗氧化能力显著高于对照组水平,之后缓慢下降,抗氧化应激体系在48 h 时以足以消除因力竭运动而影响的内环境失衡产生的氧化应激,但幼虾保持体内的高抗氧化能力至72 h,以应对具有潜在可能的下一次氧化应激。 在实际的斑节对虾养殖和增殖放流过程中,应避免频繁捕捞和持续高密度暂养。在捕捞或运输之前,增加日粮中的维生素D、类胡萝卜素、虾青素等非酶抗氧化添加剂,铜、锌和硒等合成抗氧化酶的元素的含量,在暂养和运输水体中添加抗应激剂。在对虾类放流苗种育肥过程中尝试进行运动训练,增强幼虾的抗氧化能力,保障增殖放流效果和养殖效益。
2.2 斑节对虾幼虾在力竭运动后行为学的变化
2.3 斑节对虾幼虾力竭运动后抗氧化酶活性的变化
2.4 斑节对虾幼虾力竭运动后总抗氧化能力的变化
2.5 斑节对虾幼虾力竭运动后MDA 含量的变化

3 讨论
3.1 力竭运动后斑节对虾幼虾体内抗氧化酶活性的变化
3.2 斑节对虾幼虾力竭运动后体内总抗氧化能力与MDA 含量的变化
4 小结
