六株红酵母抗氧化活性的研究
2020-11-02王悦欧阳丹汤伟刘仕博顾燕何增国
王悦 欧阳丹 汤伟,2 刘仕博 顾燕 何增国,2,3
(1. 中国海洋大学医药学院,青岛 266000;2. 青岛海洋生物医药研究院,青岛 266000;3. 青岛百奥安泰生物科技有限公司,青岛 266000)
红酵母广泛分布在海洋和陆地环境中[1],能够耐受重金属、氧化、辐射及其他极端环境,具有营养丰富、发酵周期短、可高密度发酵[2]等特点。诸多研究表明红酵母含有大量的营养成分和活性物质,如蛋白质、多糖和维生素E等营养因子,红酵母红素、蛋-胡萝卜素、番茄红素、虾青素、超氧化物歧化酶(Superoxide dismutase,SOD)和过氧化氢酶(Catalase,CAT)等活性物质[3]。它们在抗氧化清除自由基[4]、抗肿瘤[5-6]及提高免疫力[7]等方面均有很好的效果。由于红酵母可以天然合成类胡萝卜素[8],安全风险低,因此人们对能产生类胡萝卜素的红酵母兴趣在逐年增加[9]。据统计,全球类胡萝卜素市场呈不断增长趋势,2017年全球类胡萝卜素市场总销售额达到15亿美元,预计2017-2022年复合年增长率(CAGR)为5.7%,到2022年将达到20亿美元[10]。
迄今为止,红酵母的研究多集中于菌株筛选、培养基优化、发酵条件优化、类胡萝卜素分离鉴定等方面[11-15];利用诱变技术、代谢工程和基因工程等方法提高类胡萝卜素产量的研究也在逐年增加[16-19]。然而有关红酵母抗氧化性能的研究并不是很多,针对红酵母不同组分的抗氧化性能研究更是少见。曾有研究发现重金属[20]和紫外辐射等[21]极端条件提高了红酵母的抗氧化性能,也有添加适量的红酵母有效改善幼参的消化酶和免疫活性以及提高尼罗罗非鱼的生长性能、消化酶及免疫酶活性的报道[22-23]。红酵母作为天然的微生态抗氧化剂,在其他水产养殖品种中的应用也初现端倪[24]。为更好地服务水产养殖,需要进一步挖掘红酵母的应用潜力,深入系统地探究红酵母抗氧化功能及功效已十分必要。因此,本研究先测定了6株红酵母的类胡萝卜素含量及过氧化氢耐受性来初步分析其潜在的抗氧化能力,又通过测定6株红酵母的3种组分的还原力、自由基清除能力、总抗氧化活性及超氧化物歧化酶活性来综合分析红酵母的抗氧化活性,旨在为开发应用于水产养殖的天然抗氧化剂提供菌株资源和理论依据。
1 材料与方法
1.1 材料
1.1.1 菌株与培养基 红酵母菌株LW1-LW6分别分离自海藻、海水、牡蛎、五彩苏、过山蕨和桃子,并于本实验室保存。
YPD液体培养基:葡萄糖20 g/L,蛋白胨20 g/L,酵母粉10 g/L,蒸馏水1 000 mL,自然pH,1×105Pa 灭菌20 min。
1.1.2 主要试剂和仪器 三氯乙酸(TCA)、30%过氧化氢、铁氰化钾、水杨酸、抗坏血酸、二甲亚砜(DMSO),国药集团化学试剂有限公司;二苯代苦味肼基自由基(DPPH·),源叶生物科技有限公司;总抗氧化能力(T-AOC)试剂盒、BCA法微量蛋白检测试剂盒、超氧化物歧化酶(SOD)测定试剂盒,南京建成生物工程研究所。
全波长酶标仪,美国BioTek公司;SPX-250B-Z型生化培养箱,上海博迅实业有限公司医疗设备厂;THZ-C恒温振荡培养箱,苏州培英实验设备有限公司;J-25型高速冷冻离心机,美国BECKMAN公司;SW-CJ-1D型超净工作台,苏州净化;GZX-9240MBE型电热鼓风干燥箱,上海博迅实业有限公司医疗设备厂。
1.2 方法
1.2.1 菌株培养 将6株红酵母菌按2%接种量接种于YPD液体培养基中28℃、180 r/min摇床培养16 h,充分活化后按2%接种量转接至YPD液体培养基中继续培养48 h。
1.2.2 红酵母形态学及26S rRNA分子生物学鉴定
形态学鉴定参照《酵母菌的特征与鉴定手册》[25]和《真菌鉴定手册》[26]。
分子学鉴定参照Li等[27]方法,PCR扩增引物(NL1、NL4)由上海生工设计:NL1:5'-GCATATCAATAAGCGGAGGAAAAG-3',NL4:5' -GGTCCGTGTTTCAAGACGG-3'。
1.2.3 总类胡萝卜素含量的测定 有机溶剂破壁:将红酵母发酵液在常温下,4 000 r/min离心10 min,用蒸馏水洗涤两次,留菌体,在50℃烘箱中进行烘干,得到干菌体。取干菌体0.1 g,加蒸馏水溶胀后4 000 r/min离心10 min,弃去上清,加入5 mL DMSO,室温下破壁1 h,4 000 r/min离心10 min,收集上清。破壁提取2-3次,合并上清,得到色素提取物[28-29]。
总类胡萝卜素测定:参照王岁楼[30]的方法,色素提取物在478 nm有最大吸收峰,提取物中总类胡萝卜素含量按以下公式计算:总类胡萝卜素含量(μg/g)=(A478×V)/(0.16×W)式中,A478:待测样品在478 nm处的吸光值;V:加入的溶剂体积(mL);0.16:吸光系数;W:菌体干重(g)。
1.2.4 样品制备 发酵上清液、完整细胞和色素提取物的制备:取发酵48 h的红酵母菌液,4 000 r/min离心15 min,即可获得的发酵上清液。离心后的菌体沉淀用蒸馏水洗涤3次,并且调节胶红酵母细胞浓度为1.0×108CFU/mL,此为完整细胞。将菌体烘干,用DMSO进行色素提取,最终得到色素提取物[29]。
1.2.5 H2O2耐受性的研究 将活化好的菌液按2%接种量分别接种于含有不同浓度H2O2(终浓度为0、1、3、5、7、9、11 mmol/L)的YPD液体培养基中,28℃、180 r/min摇床培养48 h后,取100 μL菌液稀释涂布于YPD固体培养基上,在28℃恒温箱倒置培养48 h后进行菌落计数。以不添加H2O2的处理为对照组,同样条件下计数,以对照组菌活数定义为100%。
1.2.6 抗氧化活性的测定 采用化学法测定6株红酵母3种组分的还原力、DPPH·和·OH的清除能力,以及T-AOC和SOD活性,以此来评估3种组分的体外抗氧化能力。还原力的测定参照Ma等[31]的方法;DPPH·清除力的测定参照李慧翔等[32]和Lin等[33]的方法;·OH清除力的测定参照Gutteridge等[34-35]的方法;T-AOC和SOD活性的测定参照南京建成试剂盒说明书操作。
2 结果
2.1 菌株形态学及分子生物学鉴定
6株红酵母在YPD固体培养基上于28℃培养3-4 d,菌落直径约3-5 mm,橘红色,菌落呈典型的酵母形态(图1)。
对6株红酵母进行26S rRNA分子生物学鉴定,鉴定结果为5株胶红酵母和1株锁掷孢酵母(表1)。
2.2 总类胡萝卜素含量
6株红酵母总类胡萝卜素含量存在显著性差异(P<0.05),但总类胡萝卜素含量高低与菌株来源无对应关系(表2)。其中从五彩苏中筛选出的LW4总类胡萝卜素含量较高,达到162.91 μg/g,与其他菌株存在极显著性差异(P<0.01),总类胡萝卜素含量最低的是从桃子中筛选得到的锁掷孢酵母,其含量为117.78 μg/g。本研究中,红酵母属(Rhodotorula)的总类胡萝卜素含量要极显著高于锁掷孢酵母属(Sporidiobolus)。

图1 六株红酵母菌落形态
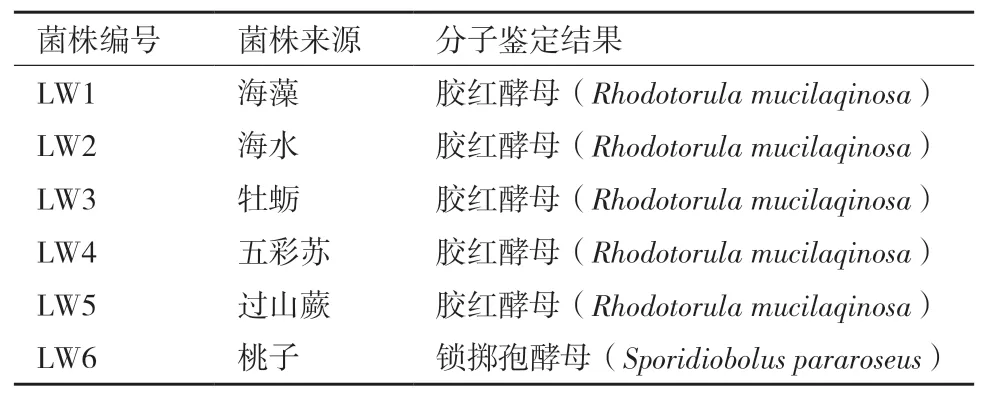
表1 六株红酵母分子鉴定结果

表2 六株红酵母的总类胡萝卜素含量
2.3 菌株的H2O2耐受性
如图2所示,1、3、5 mmol/L H2O2对于菌株的生长均有一定程度的促进作用。当H2O2浓度大于5 mmol/L时,部分菌株相对存活率开始降低,H2O2浓度过高会对红酵母生长有抑制作用。其中LW3的H2O2耐受性最强,在11 mmol/L H2O2添加量时仍有55.33%的存活率;其次是LW2和LW6,相对存活率分别为35.94%和31.51%;而LW1、LW5在11 mmol/L添加量时相对存活率为0。6株菌中耐受性最低的LW4,在9 mmol/L H2O2时菌体全部死亡。

图2 不同浓度H2O2对红酵母生长的影响
2.4 红酵母抗氧化活性
2.4.1 还原力 样品在700 nm的吸光值(图3),6株红酵母的3种组分均具有还原力,且存在显著性差异(P<0.05),并呈现一定趋势,6株红酵母3种组分的还原力大小依次为发酵上清液>色素提取物>完整细胞。

图3 六株红酵母各分离组分还原力
2.4.2 DPPH· 试验中分别对6株菌的发酵上清液、完整细胞和色素提取物进行了DPPH·清除力测定。由图4可知,6株红酵母的DPPH·清除能力具有显著性差异(P<0.05),且趋势明显。完整细胞的清除能力最强,清除率均大于89%,其中以 LW3清除率最高,达到97.48%,显著高于其他菌株;其次为发酵上清液,其DPPH·清除率在18%-55%之间;色素提取物清除DPPH·的能力最低,均维持在20%-35%。

图4 六株红酵母各分离组分DPPH·清除率
2.4.3 ·OH清除能力 试验中分别对6株红酵母的3种组分发酵上清液、完整细胞和色素提取物进行了·OH清除力测定。由图 5可以看出,完整细胞和色素提取物的·OH清除能力均优于发酵上清液,且6株红酵母的清除能力存在一定的显著性差异(P<0.05),可能是由于红酵母清除·OH的活性物质主要存在于细胞内,而发酵上清液中存在较少。

图5 六株红酵母各分离组分羟自由基清除率
2.4.4 总抗氧化能力(T-AOC)和超氧化物歧化酶(SOD)能力 根据DPPH·清除力、·OH清除力及还原力的测定结果,选取两株抗氧化能力较好的菌株LW3和LW5,对其发酵上清液和完整细胞进行T-AOC及SOD活性的测定。表3表明LW3的发酵上清液和完整细胞的T-AOC和SOD活性均高于LW5。
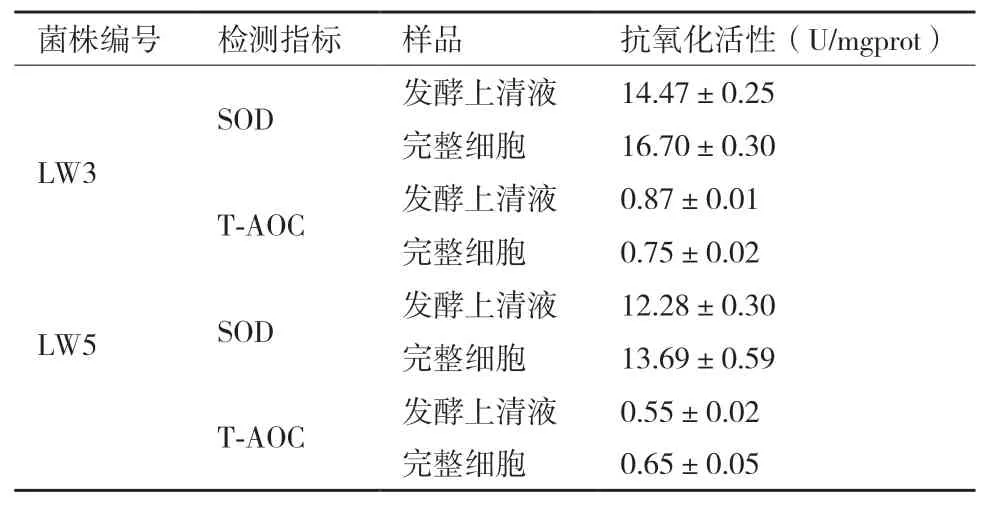
表3 LW3和LW5的SOD和T-AOC活性
3 讨论
红酵母作为一种重要的真核微生物,在生长过程中可能会面临各种各样的胁迫,氧化胁迫是其中之一。H2O2作为强氧化剂,能刺激红酵母迅速合成具有自我修复功能的活酵母细胞衍生物(Live yeast cell derivative,LYCD),对细胞起到抗氧化的保护作用[36]。Janda等[37]研究H2O2(0.9-5.3 mmol/L)对黏红酵母生长状况的影响,随着加大H2O2浓度,发酵液中的菌活数逐渐降低。Irazusta等[38]发现暴露于0.5 mmol/L H2O2的胶红酵母RCL-11生长受到严重抑制,在培养24 h后才开始缓慢生长。仓一华等[39]也曾通过添加H2O2筛选耐受性菌株,并发现当H2O2浓度达到3 mmol/L时受试红酵母菌株100%致死。在本研究中发现6株红酵母在9 mmol/L H2O2添加量存在下仍具有较高的存活率,均高于上述报道。
有意思的是,在本试验中H2O2耐受性最高的菌株LW3在总类胡萝卜素含量上显著低于菌株LW4,但总类胡萝卜素含量最高的LW4其H2O2耐受性反而是6株菌中最低的。除此之外,其他4株红酵母总类胡萝卜素和H2O2耐受性也不存在线性关系。有研究表明16 mol/L H2O2处理后降低了R. mucilaginosaC2.5t1的类胡萝卜素含量和胡萝卜素生成基因的转录水平,与胶红酵母可以调节类胡萝卜素含量来抵消H2O2的促氧化作用的假设相符[40]。更重要是的,不同种类、含量的类胡萝卜素对于H2O2的敏感性也存在差异,可能会出现总类胡萝卜素含量高的菌株其H2O2耐受性不一定最显著的现象。
有关红酵母抗氧化性能方面的研究陆续已有报道,但到目前为止,对于红酵母不同组分的抗氧化能力研究甚少。本研究分别对6株红酵母的发酵上清液、完整细胞和色素提取物的抗氧化能力进行了比较分析。实验采用了简便的化学方法,应用不同自由基系统(还原力、DPPH·自由基和·OH自由基等),以期准确地反映红酵母3种组分的抗氧化能力[41]。实验结果显示6株红酵母的发酵上清液、完整细胞和色素提取物均具有一定的清除自由基的能力。样品的还原力与抗氧化能力之间存在着正向关系,还原力越大,抗氧化能力越强[42],实验发现6株红酵母3种组分的还原力依次为发酵上清液>色素提取物>完整细胞,发酵上清液还原力在0.26-0.39之间,显著优于其他两个组分。DPPH·作为一种人工合成的具有单电子的有机自由基,可以与抗氧化剂进行单电子配对,溶液的褪色程度与单电子配对数呈现一定关系[43],实验表明6株红酵母DPPH·清除能力依次为完整细胞>发酵上清液>色素提取物。·OH是生物分子与过渡金属离子参与芬顿反应造成氧化损伤时所产生的,抗氧化剂能够螯合过渡金属离子从而抑制·OH的生成[44],实验表明6株红酵母完整细胞和色素提取物的·OH清除能力显著优于发酵上清液。对于3种不同自由基的清除,3个组分均展现出不同的能力,这可能是与不同菌株胞内胞外的抗氧化活性物质的差异和发酵液中代谢产物的不同有关。
红酵母细胞内具有多种抗氧化酶以及类胡萝卜素,不同的类胡萝卜素保护细胞氧化应激和调控抗氧化的能力各有不同[45-46]。Xu等[47]用5 mg/kg番茄红素和0.75 mg/kg黄曲霉毒素(Aflatoxin B1,AFB1)给小鼠灌胃给药30 d后发现番茄红素不仅减轻了AFB1诱导的免疫抑制,还增加了总抗氧化能力(T-AOC)和酶活性;裴凌鹏等[48]发现添加适量的虾青素可以部分修复小鼠由于H2O2诱导造成的成骨氧化损伤。另外,研究还发现耐辐射奇球菌(Deinococcus radiodurans,DR)也能够产生类胡萝卜素,并且具有很强的抗氧化能力,它所产生的一种特殊类胡萝卜素deinoxanthin其自由基清除能力远高于番茄红素(lycopene)和 β-胡萝卜素(β-carotene),当D. radiodurans的 类 胡萝卜 素提取物浓度为 0.6 μg/mL时,对DPPH·清除率达到46%[49],而本实验中6株红酵母色素提取物浓度在1.18-1.96 μg/mL,但是DPPH·清除率均低于35%,6株红酵母的抗氧化能力显著低于耐辐射奇球菌,这可能与菌株胞内的类胡萝卜素种类及含量有关。红酵母胞外发酵液中还可能存在多糖、抗氧化蛋白等抗氧化物质。本试验中6株红酵母发酵上清液还原力明显高于完整细胞和色素提取物,可能与发酵液中存在的抗氧化代谢物质有关。马文锦等[50]研究发现胶红酵母胞外多糖具有清除自由基的能力,其多糖组分REPS2-A的还原力为 0.352,DPPH·清除率为49.1%。本实验中LW1发酵上清液还原力最大为0.39,DPPH·清除率为55.05%,清除自由基能力优于上述结果。另外,红酵母细胞壁的组成成分几丁质具有结合重金属离子的特性,在重金属诱导条件下,产生更多的金属硫蛋白(MT)[51],体外研究表明MT具有抗氧化活性,能有效清除羟自由基[52],因此完整细胞表现出较高的·OH清除能力,可能与细胞表面的抗氧化物质组成成分有关。通过分析红酵母3种组分的抗氧化活性,推测受测菌株的抗氧化能力与胞内胞外抗氧化物质的差异及发酵液中代谢产物的不同有关,其他未知的抗氧化物质等相关研究还有待深入。在未来的研究中我们将进一步分析红酵母菌株的类胡萝卜素组分及含量,并动态跟踪红酵母在发酵过程中产生抗氧化物质的能力,以期建立全面系统评价红酵母作为直用液体产品的功效评价方法。
4 结论
6株红酵母具有很强的H2O2耐受性(在9 mmol/L H2O2添加量时仍有较高的存活率);3种组分均具有一定程度的还原力、自由基清除能力、总抗氧化活性及超氧化物歧化酶活性。通过综合分析研究红酵母的抗氧化活性,为开发应用于水产养殖的天然抗氧化剂提供菌株资源和理论依据。
