辽宁省水稻品种抗稻瘟病基因型鉴定
2020-10-23谷思涵李思博魏松红
谷思涵,李思博,魏松红,刘 伟
(沈阳农业大学 植物保护学院,沈阳 110161)
到2050 年,世界人口将达到90 亿,伴随着人口快速增加而来的粮食危机将愈演愈烈,而人口增长又主要集中在以水稻、小麦、玉米等为主食的发展中国家[1]。为了满足此需求,水稻生产将面临诸多挑战,由稻巨角间座壳(Magnaporthe oryzae)引起的稻瘟病世界性水稻病害,也是我国水稻生产上的三大病害之一[2]。水稻中天然携带能够抵御稻瘟病菌的抗瘟基因,目前大量的抗稻瘟病基因已被定位和克隆,为全面认识稻瘟病菌与水稻互作提供了有力依据[3]。水稻抗稻瘟病的基因已发现80 余个,其中Pib、Pita 和Pi9 等28 个基因已被成功克隆,这些基因大多分布于第6,11,12 号染色体上。作为稻瘟病抗性基因密集区域,第6 染色体的Pi9 和 Pi2 位点周围已被成功克隆10 个以上的抗病基因[4]。AMANTE-BORDEOS 等(1992)首次从野生稻中发现具有广谱抗性基因Pi9,并获得了RFLP 分子标记[5]。与Pi9 来自同一位点上的抗病基因Pi2 由MEW 等(1994)利用分子标记定位于RG64 和AP22 之间[6],ZHOU 等[7]运用图位克隆法完成克隆。抗病基因多数属于NBS-LRR 类基因,这类基因都包含两个结构域,核苷酸结合位点(NBS)与富亮氨酸重复序列(LRR)。LRR 结构域因重复序列数量、长度及组合各不相同,导致组成的抗性蛋白结构各有差异[8]。LRR 能够与无毒基因编码产物直接作用,影响植株的抗病性。当LRR 结构单一时,易被无毒基因编码变异产物攻克,从而使品种抗性丧失,所以多基因聚合品种与抗谱多样化已成为当前品种抗病性改良的主要研究方向[9]。本试验通过对185 份辽宁省水稻生产品种和实验材料中 9 种抗稻瘟病基因 Pi5、Pi36、Pi37、Pib、Pid2、Pid3、Pik、Pikh 和 Pita的携带情况进行鉴定,以期为稻瘟病的抗病育种品种、合理布局及科学防治提供依据[10]。
1 材料与方法
1.1 材料
供试185 份水稻(Oryza Sativa L.)材料收集自辽宁省内7 个主要水稻产区:沈阳、大连、丹东、营口、盘锦、抚顺和铁岭,其中29 份为生产品种,155 份为育种材料,1 份为对照(高感品种丽江新团黑谷)。
辽宁省稻瘟病菌主要致病菌株采自辽宁省7 个主要水稻产区沈阳、大连、丹东、营口、盘锦、抚顺和铁岭。
1.2 方法
1.2.1 稻瘟病菌单孢分离及产孢培养 病菌单孢分离:在TOA(番茄燕麦琼脂培养基)平板上将保存于4℃或-80℃冰箱内的稻瘟病菌活化,25℃恒温培养5~10d,至有灰色孢子产生。然后在显微镜下挑取单个孢子转入PDA 培养基培养,待菌丝生长较多时转入TOA 平板,25℃恒温培养7~10d。
病菌产孢培养:在湿润条件下用无菌棉签轻刮去产孢平板表面菌丝。于超净工作台无菌风下吹干,盖双层灭菌纱布。在模拟自然条件下培养5~7d,直至孢子大量产生。
菌悬液配制:用3mL 的无菌蒸馏水将产孢培养平板上的孢子洗脱。用血球计数法在显微镜下将稻瘟病菌的分生孢子浓度调整为每毫升2.5×106个孢子,以备接种采用。
1.2.2 水稻秧苗培育 将有机土与基质按2∶1 混合均匀,每个穴盘内放置一半底土,再将鉴定材料穴播于育苗盘内,每份材料播1 穴,每穴10 粒种子,后用混合土封顶均匀。育苗期间每天换水1 次,每天保持通风。
1.2.3 接种鉴定 于水稻3 叶1 心时期,将配制好的各菌株菌悬液均匀喷于叶片上,每品种3 次重复,接种后24h 用塑料膜覆盖黑暗条件下保湿,模拟自然条件观察发病情况。
1.2.4 调查记载标准 记载标准根据全国稻瘟病菌生理小种联合试验组于1980 年制定的统一标准[11],于接种8~10d 后调查菌株在不同供试品种上的抗病(R)反应类型和感病(S)反应类型。
1.2.5 抗病比率计算 抗病比率即为表现为抗病的植株占总数的比率。

1.2.6 水稻品种基因组提取 采用TIANGEN 植物基因组DNA 提取试剂盒(DP320-02)提取水稻DNA。
1.2.7 引物及反应程序 参考文献引物[12-13]及优化反应体系和条件(表1),Primer1~9 用于稻瘟病抗性基因的鉴定。每个水稻品种检测各类稻瘟病抗性基因设3 次重复。PCR 反应体系 (25μL):2×Taq PCR Master Mix(康为世纪生物科技有限公司,CW0682M)12.5μL,正向引物和反向引物各 1μL(10μg·mL-1),DNA 模板1μL(20~80ng·mL-1),之后用 ddH2O 补足至 25μL。PCR 反应程序:94℃预变性 5 min,94℃变性 30~60s,54~60℃退火60s,具体依据引物的G/C 含量来定,72℃延伸30~90s,具体依据目的片段长度而定,30 个循环,72℃延伸5min。
1.2.8 PCR 扩增结果的电泳检测 4μL PCR 产物与 2μL 1.5×Loading buffer 混合液(4∶1)混合后,在 1%琼脂糖凝胶中电泳,Goldview 染色后,在凝胶成像系统下拍照。
2 结果与分析
2.1 185 份供试水稻材料的表型鉴定
由表2 可知,185 份材料中,接种稻瘟病菌表现高抗(HR)有9 份,占总数的4.86%,分别为盐粳231、辽粳401、沈农 315、沈农 Y16、沈农 Y65、沈农 H4、沈农 H9、沈农 H17 和沈农 H28;表现抗病(R)和中抗(MR)的材料各70 和71 份,分别占总数的37.84%和38.38%;表现感病(S)的材料为4 份,表现中感(MS)的材料为30份,分别占总数的2.16%和16.22%。

表1 抗病基因引物Table1 Primers for resistance genes
2.2 185 份供试水稻材料的抗稻瘟病基因型鉴定
由表3 可知,在185 份供试水稻材料中,携带抗病基因Pita 的共184 份,占总数的99.46%,携带该基因且表现抗病的水稻材料共149 份,占总数的80.54%,为分布频率最高的抗病基因类型。携带抗病基因Pik 的共63 份,占总数的33.51%,携带该基因且表现抗病的水稻材料共53 份,占28.65%,为分布频率最低的抗病基因类型。其余的分布频率各不相同。
2.3 单基因及多基因聚合的材料统计及其抗性分析
如表4 所示,当水稻材料只携带单基因Pita 时抗病比率为75%;当水稻材料同时携带2 种基因Pib 和Pid3、Pid3 和 Pita、Pikh 和 Pita 时抗病比率为 100.00%;当水稻材料同时携带 3 种基因 Pid3、Pikh、Pita 时抗病比率为 100.00%;当同时携带 4 种基因 Pi5、Pid3、Pikh、Pita 和 Pid2、Pid3、Pikh、Pita 时抗病比率为 100.00%;当同时携带 5 种基因 Pi36、Pid2、Pid3、Pikh、Pita 和 Pid2、Pid3、Pik、Pikh、Pita 时抗病比率为 100.00%,而携带Pi36、Pi37、Pid2、Pid3、Pita 抗病比率为0;当同时携带6 种抗病基因时,抗病比率由 0.00%到100.00%不等;当同时携带7 种抗病基因时抗病比率由42.86%到100.00%不等;当同时携带8 种抗病基因时抗病比率由80.00%到100.00%不等;当同时携带9 种抗病基因时抗病比率仅为72.73%。
3 讨论与结论
植物受体常常是由抗病R 基因编码的富亮氨酸重复蛋白或者激酶。因为抗性依赖于受体配体的相互作用,大多数R 基因在抗病品种与易感品种的遗传杂交中占主导地位[14-16]。大多数R 基因编码NLR 蛋白质,但Pid2 是例外,它对受体型激酶细胞外B 凝集素和细胞内的血清素苏氨酸激酶域进行编码。第二个例外是Pi21 基因,它编码了一种富含蛋白质的假定重金属结合域。大多数的抗性基因聚集在一起,通常位于6 号和11 号染色体上[17]。Pi9与 Pi2 位点包含一组 NLR 基因,其中一类基因有5 个等位基因分别为 Pi9、Pi2、Piz-t、Pi50 和Pigm,另一类有3 个等位基因Pi-d3、Pid3-A4 和Pi25。抗病基因与无毒基因互作有3 种形式,第1 类主要是典型的一种R 基因和一种Avr 基因符合基因对基因的互作模式,例如Pi-ta 与Avr-pita[18]和Pi54 与Avr-pi54[19],端粒连接的毒性基因Avr-pita,用中性的方式对分泌蛋白进行编码,其编码的功能蛋白质是一种成熟的蛋白酶包含c 端176 个氨基酸[20],pita 编码了一种有928 种氨基酸的细胞质NLR 受体。Avr-pi54 编码一个预测n-末端分泌一个信号肽(SP)的分泌蛋白,Pi54 前身为Pik-h,是从水稻特特普中克隆出来的编码预测330 aa-NLR 蛋白质,与Pita 不同Pi54 并不直接表达,而是诱导病原体攻击的防御表达。第2 类是两种R 基因和一种Avr 基因的互作模式,例如Pik-1 和Pik-2 与Avr-pik[21-22]。第3 种互作类型建立在第2 种类型之上的模式,两种不同的Avr 蛋白可以被一个复杂的R 蛋白识别,例如,复合NLR 蛋白质RGA4 与 RGA5异质二聚体可以与 Avr-pia 或 Avr1-co39 其中任何一个相互作用[23]。本试验中携带 Pi36、Pi37、Pib、Pid2、Pid3、Pik、Pikh 和Pita 的23 份水稻材料的抗病比率高达100%,而携带9 种抗病基因时抗病比率仅为72.73%。由此可见并非携带抗病基因种类越多抗性越好,这可能是由于抗病基因之间存在某种关联,或是协同作用或是拮抗作用,这种关联促进了水稻抗病性的提高,另一方面当水稻所携带抗病基因数量较多时,体内的抗性基因之间是否存在拮抗作用反而降低了水稻抗性,对于抗性基因之间的互作还需进一步研究。

表2 185 份供试材料的表型鉴定Table 2 Phenotypic identification of 185 test varieties
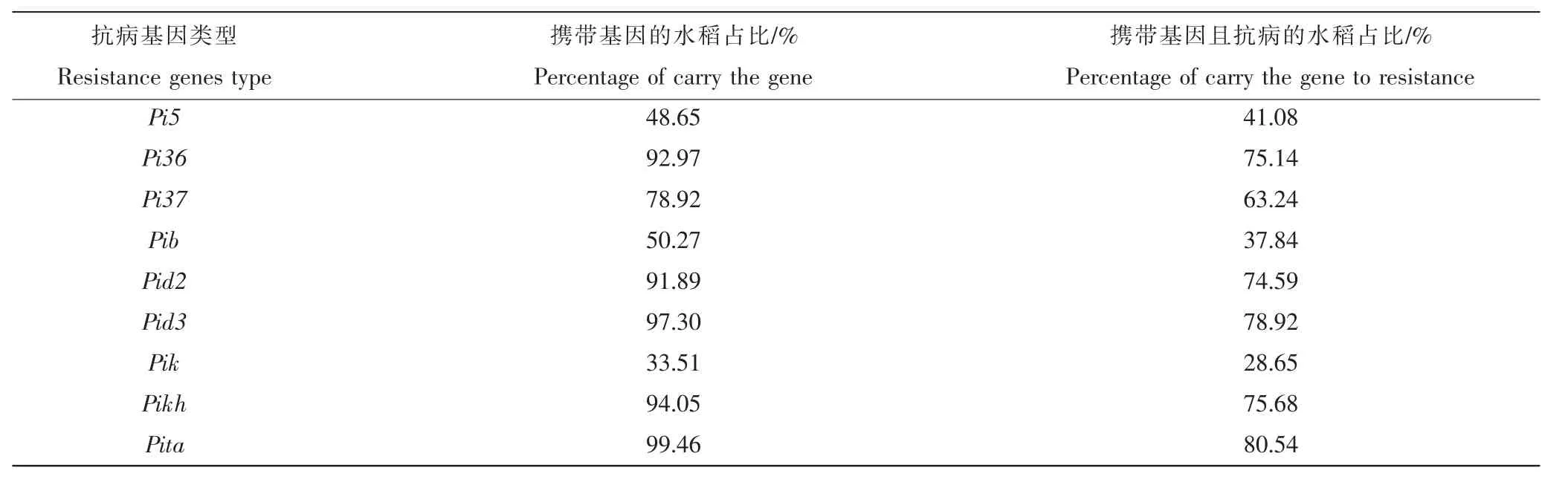
表3 水稻抗病基因分布频率Table 3 Gene distribution frequency of resistance genes
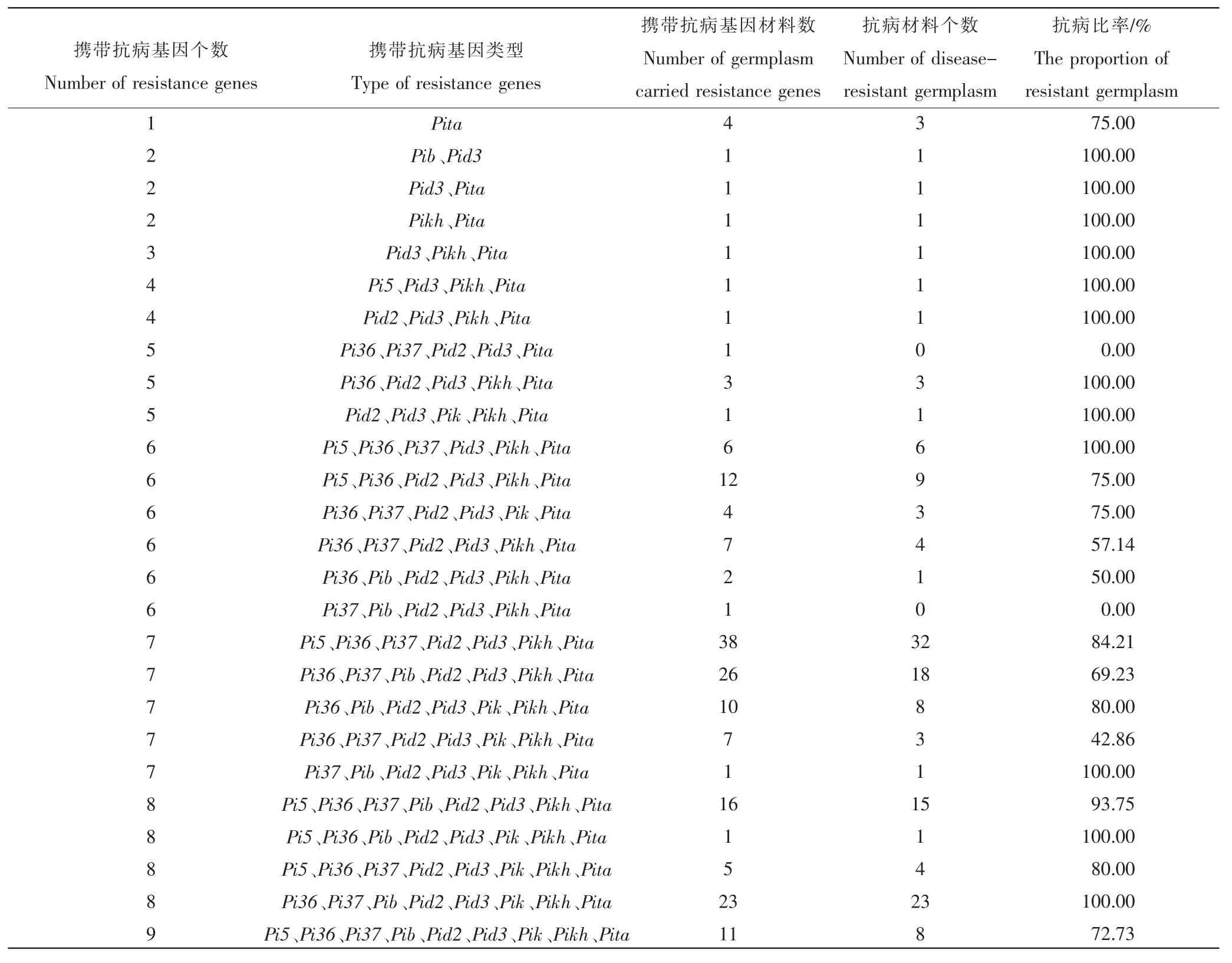
表4 供试185 份水稻材料所携带的抗稻瘟病基因类型及数目统计Table 4 The type and number of blast resistant genes in 185 rice samples were analyzed
试验结果表明携带抗病基因Pita 共184 份占总数的99.46%,为分布频率最高的抗病基因类型。携带抗病基因Pik 共63 份占总数的33.51%,为分布频率最低的抗病基因类型。其余基因Pi5、Pi36、Pi37、Pib、Pid2、Pid3、Pikh 的分布频率各不相同。有 23 份水稻品种同时携带有 Pi36、Pi37、Pib、Pid2、Pid3、Pik、Pikh 和 Pita 且抗病比率为 100%,说明 Pi36、Pi37、Pib、Pid2、Pid3、Pik、Pikh 和 Pita 基因之间可能存在协同效应,它们共同作用增加了水稻材料的抗性。为了阐明寄主和病原体的共同进化,并提供预防或控制稻瘟病的适当方法,全世界研究人员时刻关注并开发新的能有效鉴定无毒基因和抗性基因的方法。本研究从分子生物学的角度出发,通过对辽宁省水稻品种抗稻瘟病基因型鉴定,为抗病育种及辽宁省稻瘟病的防控提供了依据。
